
Araeosoma Mortensen, 1903
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3609.6.1 |
|
publication LSID |
lsid:zoobank.org:pub:046410E8-079F-4FBF-B0EF-361DFCE92E2C |
|
DOI |
https://doi.org/10.5281/zenodo.5622874 |
|
persistent identifier |
https://treatment.plazi.org/id/275FE573-6D71-FFFA-8C99-FEADFB183C16 |
|
treatment provided by |
Plazi |
|
scientific name |
Araeosoma Mortensen, 1903 |
| status |
|
Araeosoma Mortensen, 1903 View in CoL
Type species: Calveria fenestrata Wyville Thomson, 1872 , by original designation.
Diagnosis —Test large, depressed. Conspicuous membranous interstices between plates, especially those of aboral surface. Primary ambulacral plates in contact for most of their circumference, fully occluding the much smaller accessory-plates. Pore-pairs on oral surface arranged in three series on either side of the interporiferous zone. Blades of the dactylous pedicellariae with an ornately curved and fenestrated distal expansion (emended from Fell 1966).
Remarks —There are 13 assigned species, all recent ( Table 1 View TABLE 1 ).
Holotype —From the Chatham Rise, east of New Zealand (from Scroll seamount in the Graveyard seamount complex): 42° 47.4´S, 180° 0.0´W, 870–1060 m (131 mm TD), NIWA23720. Deposited in the NIC, Wellington. Taken with a bottom trawl on 21 April 2001 by RV Tangaroa during research into the ecology and effects of fishing on seamounts. Stored in 80% ethanol.
Paratypes —Three specimens, all from the Graveyard seamount complex; one specimen from Diabolical seamount, 42° 48.0´S, 179° 58.8´W, 900–1058 m (138 mm TD), NIWA23719; one specimen from Zombie seamount, 42° 46.2´S, 179° 55.8´W, 932–1070 m (131 mm TD), NIWA25961; one specimen from Graveyard seamount, 42° 46.2´S, 179° 59.4´W, 951–1076 m, (68 mm TD), NIWA25964. Specimen NIWA25961 stored dry, the rest stored in 80% ethanol.
Other material —Chatham Rise: 1 specimen (130 mm), 42° 46.5´S, 177° 15.9´W, 987–996 m, NIWA23725; 1 specimen (125 mm TD), 42° 45.9´S, 179° 59.2´W, 951–1076 m, NIWA25962; 2 specimens (80, 74 mm TD), 44° 10.6´S, 174° 33.6´W, 760–960 m, NIWA53906. Macquarie Ridge (south of New Zealand): 2 specimens (100, 99 mm TD), 51° 3.9´S, 164° 35.3´E, 968–973 m, NIWA23717; 4 specimens (56, 70, 70, 84, mm TD), 51° 3.2´S, 164° 36.4´E, 990– 973 m, NIWA23722. Ritchie Banks (east of New Zealand): 1 specimen (107 mm TD), 39° 28.6´S, 178° 25.2´E, 874 m, NIWA23727; 1 specimen, 39° 30.1´S, 178° 23.2´E, Te Papa J 070124. Northwest Challenger Plateau (west of New Zealand): 1 specimen (101 mm TD), 37° 30.0´S, 167° 41.0´E, 943 m, NIWA45137. Tasmania (south of): 1 specimen, 44° 5.9´S, 147° 24.7´W, 810–1020 m, AM J24496 View Materials . All stored in 80% ethanol, except for NIWA25962 and one specimen in NIWA23717 which are stored dry.
Etymology —Named after the “bidentate” tridentate pedicellaria present in the species, used as a noun in the genitive case.
Diagnosis —Adults moderate size, up to 138 mm TD; colour of test and appendages deep red, peristome mustard yellow; relatively narrow membranous spaces between plates, wider in the interambulacra and aborally; primary tubercles on every plate in the adradial interambulacral series on the oral side; spines delicate and frequently lost; hoofs white, short, narrower than the spine, not flared; bidentate pedicellariae abundant, dactylous pedicellariae with 5 valves.
Description —Test of holotype ( Figure 1 View FIGURE 1 ) large (test diameter 131 mm), circular, flattened, ambitus rounded. Colour of test and spines deep red, owing to the pigment in the skin, mustard yellow around peristome (extending partially along ambulacra in NIWA25964). Cleaned sections showing white to light pink membranous connective tissue between coronal plates, which remain dark red. Primary tubercles perforate and non-crenulate. Ratio of interambulacrum to ambulacrum width at ambitus, 4:3.
There are 39–40 plates in the interambulacral columns cleaned for examination, 18 on the oral side and 21 or 22 on the aboral side. Ambulacral columns have 56–57 plates, 26 on the oral side and 30–31 aborally. Although plate boundaries partly concealed by skin and a thick layer of connective tissue, distinct membranous spaces are evident in the median area between successive plates in a column, particularly aborally and in the interambulacra. This feature more obvious in the dried specimens. Lateral edges of the coronal plates overlap, however, in the usual manner for this genus, and details of this were revealed by cutting open a section of the test and examining the internal plate surfaces. This showed upper (adapical) plates overlapping lower plates in the ambulacra, lower plates overlapping upper plates in the interambulacra and, at the radial suture, interambulacral plates overlapping ambulacral plates. All coronal plates short and wide, roughly oblong. Adapical-most interambulacral plates angled slightly downward to accommodate extension of genital plates into the interambulacrum.
Aboral test plating ( Figure 2 View FIGURE 2 ): There is a fairly regular series of primary tubercles in the aboral interambulacra, occurring on every second or third plate and located near the plate centre at the ambitus but angling across the plate adradially as the series is followed adapically. This series limited to the outer two-thirds of the aboral side (as it is in the other material examined except for NIWA 25962 in which it extends distinctly further toward the apical system). This pattern is more regular on the un-cleaned columns—the cleaned column having an irregular group of plates near the centre, possibly due to having been broken and repaired. Other primary tubercles (between about 1 and 6 per column) irregularly placed interradially, also mostly limited to the outer two-thirds of the aboral side. In the ambulacra, primary tubercles—slightly smaller than those in the interambulacra—occur irregularly between every second to seventh plate, forming a series on the perradial side of the pore-pairs. The remainder of the aboral plates covered with secondary and miliary tubercles—up to a maximum of about 12 (interambulacra), and 16 (ambulacra).
Apical system about 22 mm across, monocyclic, sub-pentagonal (the sides only slightly curved) but misshapen by the enlarged G2, the component plates difficult to distinguish due to a thick covering of skin. Genital pores small, opening in the membranous space just beyond the outer point of the genital plates. The genital pores of one of the paratypes (NIWA23719) distinctly larger, possibly indicating sexual dimorphism (female). Madreporite compact, hemispherical, apparently limited to plate G2 (in NIWA23717 (dry) G2 is greatly enlarged to accommodate the madreporite which, as in the holotype, is confined to this plate). This hemispherical shape also seen, where intact, in other specimens. Ocular plates with 2–5 small tubercles of different sizes, pore at the outer edge. Anus surrounded by a rough ring of plates and spinelets, and about 30 small plates (each with 1–4 small tubercles) fill the space between the main plates and the anus.
Oral test plating ( Figure 2 View FIGURE 2 ): There is a regular adradial series of primary tubercles in the interambulacra, occurring on every plate from the ambitus to the peristome, gradually diminishing in size. These tubercles frequently abutting and some of the larger adambital ones overlapping or compressed to fit within the height of the plate. An inner series of slightly smaller tubercles occurs on most plates, located near the interradial suture at the ambitus and placed gradually further across the plates, but not beyond mid-plate, as the series is traced adorally. Between these two series another, smaller, primary tubercle occurs on most plates but not so as to form an obvious series. Smaller secondary and miliary tubercles (up to about 14) scattered over the remainder of the plates. In the oral ambulacra, an inner series of primary tubercles occurs generally on every second plate in the outer half (where they are largest) and on every plate in the adoral half. A few smaller primary tubercles also found around the pore zone in the outer half of the ambulacra. Secondary and miliary tubercles otherwise occupy the remaining space on the plates—up to about 14 on the larger plates.
Peristome about 27 mm in diameter, bearing ambulacral plates only. Two overlapping columns of about 11 boomerang-shaped plates in each zone (11 also in NIWA25961, 10 in NIWA23717), each with a single tube-foot and a row of tubercles (up to six or seven) which span the width of the plate. An occasional extra plate (with tubefoot) crammed in near the outer margin. Peristomial spines, tube-feet, and pedicellariae mustard coloured, quite distinct from the red colour over the rest of the test. In NIWA25964 this mustard colour extends also onto a few adoral plates of the ambulacral columns.
Spines: Cylindrical, hollow, the largest all broken—longest remaining 15 mm long, 0.7 mm in diameter. Primary spines smooth, with about 25 fine, longitudinal striations. These striations wedge-shaped in cross-section, with an inner ring of perforations in the oral spines examined and none in the aboral spines—the spine walls relatively thin ( Figure 3 View FIGURE 3 ). Both primary and secondary spines uniformly red in colour. Several hoofs were preserved on the oral spines of the holotype but on only one or two other specimens—the oral spines being relatively fine and easily broken, even compared to other species of Araeosoma . Several hoofs were preserved on the oral spines of the holotype but only one or two remained on other specimens – the oral spines being relatively fine and easily broken, even compared to other species of Araeosoma . Glands visible on secondary spine tips.
Pedicellariae ( Figure 5 View FIGURE 5 ). The general forms of pedicellariae expected in this genus all present in the holotype; Tridentate: Large (about 2.5 mm long) rostrate form on a long stalk with a short neck; the long, involuted section with a distinct elbow and the coarsely serrated blade widening to almost the width of the base. Bidentate: In a range of sizes from about 0.5–1.5 mm, straight sided, the base barely wider than the blade; the neck nearly as long as the head, on a long stalk. The larger specimens, at least, are occasionally tridentate. These unusual pedicellariae are distinctive and readily found over much of the test. Triphyllous: typical echinothurioid form about 0.5–0.8 mm long on a long neck (several times the length of the head) and longer stalk; blade relatively short, the involuted basal part completely enclosed only for a short distance above the base. Dactylous: Typical form for Araeosoma , about 3.0– 3.5 mm long with 5-valves, blade edges finely “crimped” rather than serrated. Exceedingly rare on the holotype and other specimens searched, those found were on the aboral side near the ambitus, as is typical for this form.
Sphaeridia, elongate spherical (about 0.5 mm long by 0.2 mm wide), located adjacent (slightly perradial and adoral) to the inner tube foot in each ambulacral plate, from the apical system to the peristome ( Figure 5 View FIGURE 5 ).
Size range —The average test diameter of the 16 specimens measured was 98 mm, and the largest specimen was 138 mm TD.
Occurrence —There have been 13 confirmed records of A. bidentatum since the first specimen was taken in 1987, representing 19 individuals.
Seven of the records (representing eight specimens) are from the north and east Chatham Rise, with five of these (five specimens) taken from the Graveyard seamounts in the central north Chatham Rise ( Figure 6 View FIGURE 6 ). The other records are from the northern Macquarie Ridge, south of New Zealand at about 50° S (the southernmost records), the northwest Challenger Plateau at about 37.5° S (the northernmost record), the Ritchie Banks to the east of the North Island, and south of Tasmania (the westernmost record). Most, if not all, records are associated with seamount or ridge features and many were caught during commercial or research trawling for orange roughy ( Hoplostethus atlanticus ) and oreos ( Oreosomatidae ).
The conservative depth range for the species is 960–990 m, with a potential range of 760–1080 m. Remarks —The plate structure and tuberculation patterns of A. bidentatum are typical of Araeosoma , with no remarkable features and, in terms of the proportions of the test elements and placements of the tubercles, appear most similar A. fenestratum and A. violaceum .
The bidentate pedicellariae of A. bidentatum are unique within the order Echinothurioida , the three-valved form being otherwise the universal condition, and rarely seen in any group of regular echinoids. Although bidentate and bi-phyllous pedicellariae are common in clypeasteroids (especially Scutellidae (Mortensen 1948)) , the only other regular echinoid to have them may be Poriocidaris purpurata (Thomson, 1872) , a widespread deepwater cidaroid also present in northern New Zealand. This characteristic pedicellaria, being quite common on the specimens examined, makes the species easy to distinguish with the aid of a microscope.
The colouration of A. bidentatum is also distinctive, and is retained in all preserved material. Other distinctly red echinothurioids; A. splendens ( Indonesia) , A. belli (West Indies), as well as the new species described below ( A. tertii and Hapalosoma amynina ) can be differentiated by their pedicellariae and peristome colour.
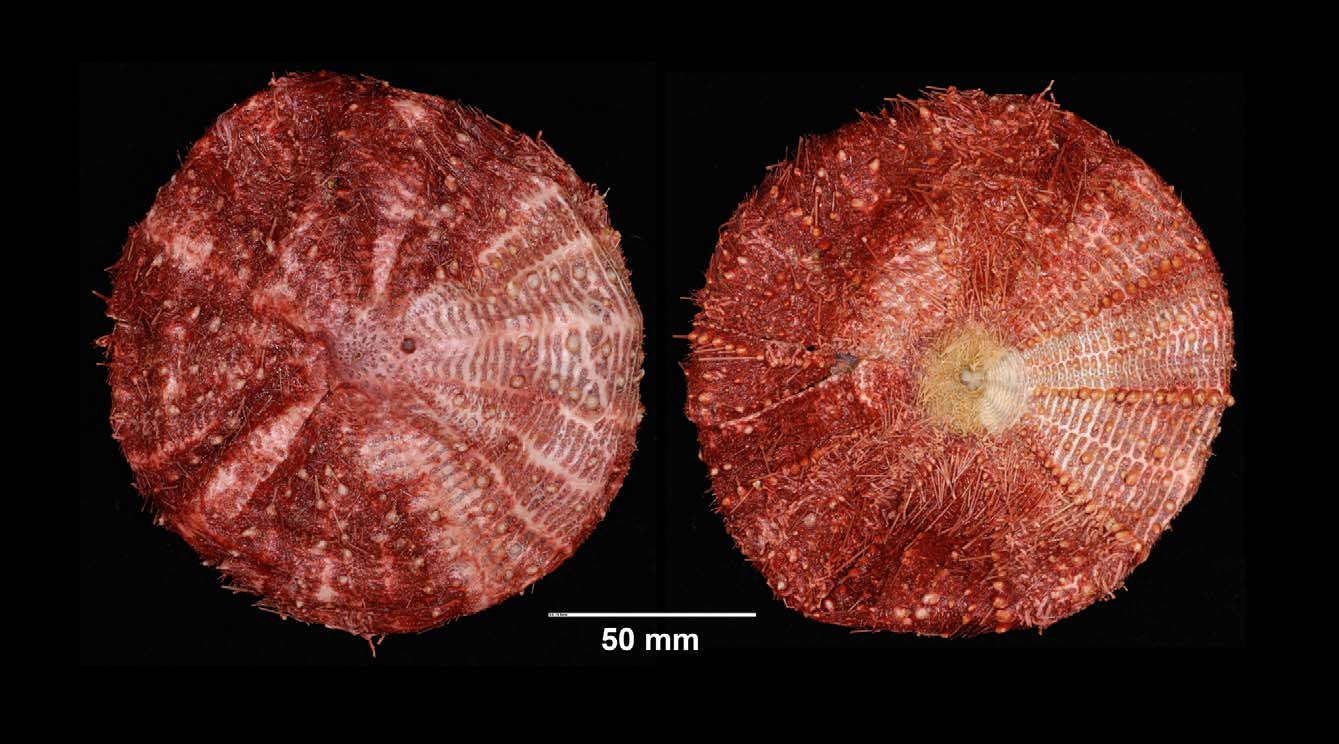

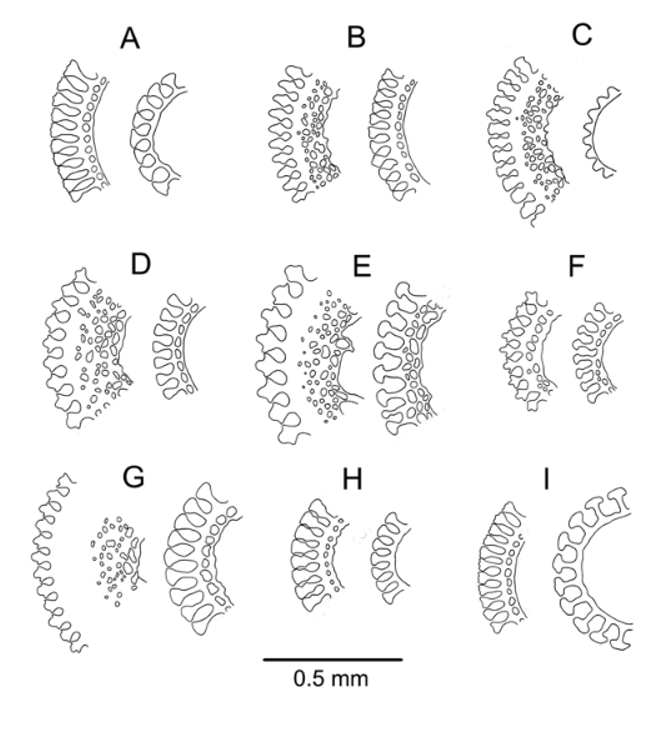

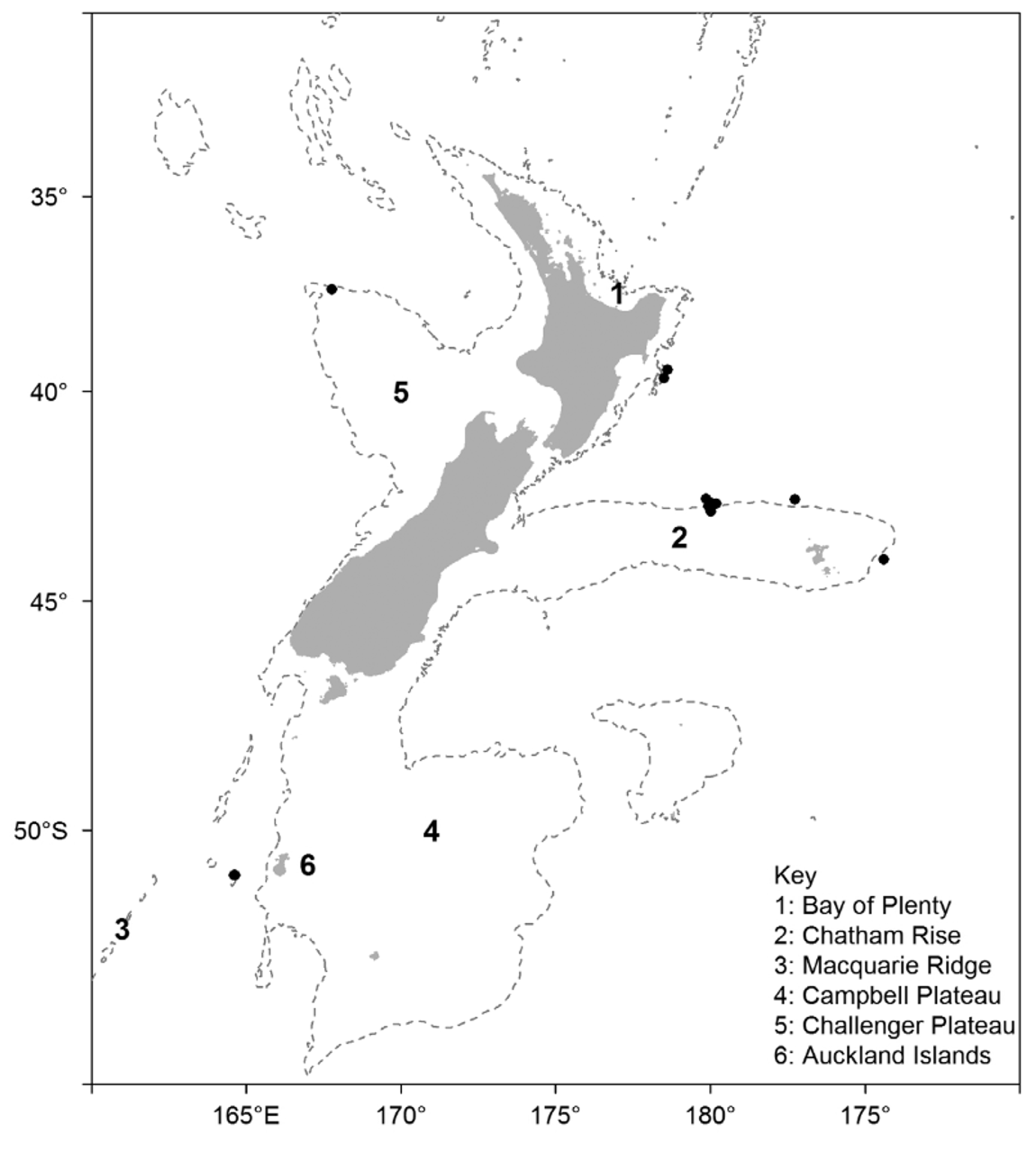
TABLE 1: Species assigned to the genus Araeosoma.
| Species | Era | Known distribution |
|---|---|---|
| Araeosoma fenestratum (Thomson, 1872) | Recent | NE Atlantic |
| Araeosoma violaceum Mortensen, 1903 | Recent | NE Atlantic |
| Araeosoma belli Mortensen, 1903 | Recent | West Indies |
| Araeosoma parviungulatum Mortensen, 1934 | Recent | Celebes |
| Araeosoma eurypatum Agassiz & Clark, 1909 | Recent | Hawaii |
| Araeosoma leptaleum Agassiz & Clark, 1909 | Recent | East Pacific |
| Araeosoma splendens Mortensen, 1934 | Recent | Kei Islands |
| Araeosoma owstoni Mortensen, 1904 | Recent | West Pacific |
| Araeosoma coriaceum (Agasssiz, 1879) | Recent | Indo-Pacific |
| Araeosoma tessellatum (Agasssiz, 1879) | Recent | Malay archipelago, China Sea |
| Araeosoma paucispinum H.L. Clark, 1924 | Recent | South Africa |
| Araeosoma thetidis (H.L. Clark, 1909) | Recent | Australia, New Zealand |
| Araeosoma alternatum Mortensen, 1934 | Recent | Western Indian Ocean |
| Araeosoma bidentatum sp. nov. Figures 1, 2, 3H, 4H, 5, 6 |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.