
Gephyromantis decaryi Angel, 1930
|
publication ID |
https://doi.org/ 10.5281/zenodo.212849 |
|
DOI |
https://doi.org/10.5281/zenodo.5672704 |
|
persistent identifier |
https://treatment.plazi.org/id/28038792-4C31-FFFE-FF7E-FB3532537E73 |
|
treatment provided by |
Plazi |
|
scientific name |
Gephyromantis decaryi Angel, 1930 |
| status |
|
Gephyromantis decaryi Angel, 1930 View in CoL
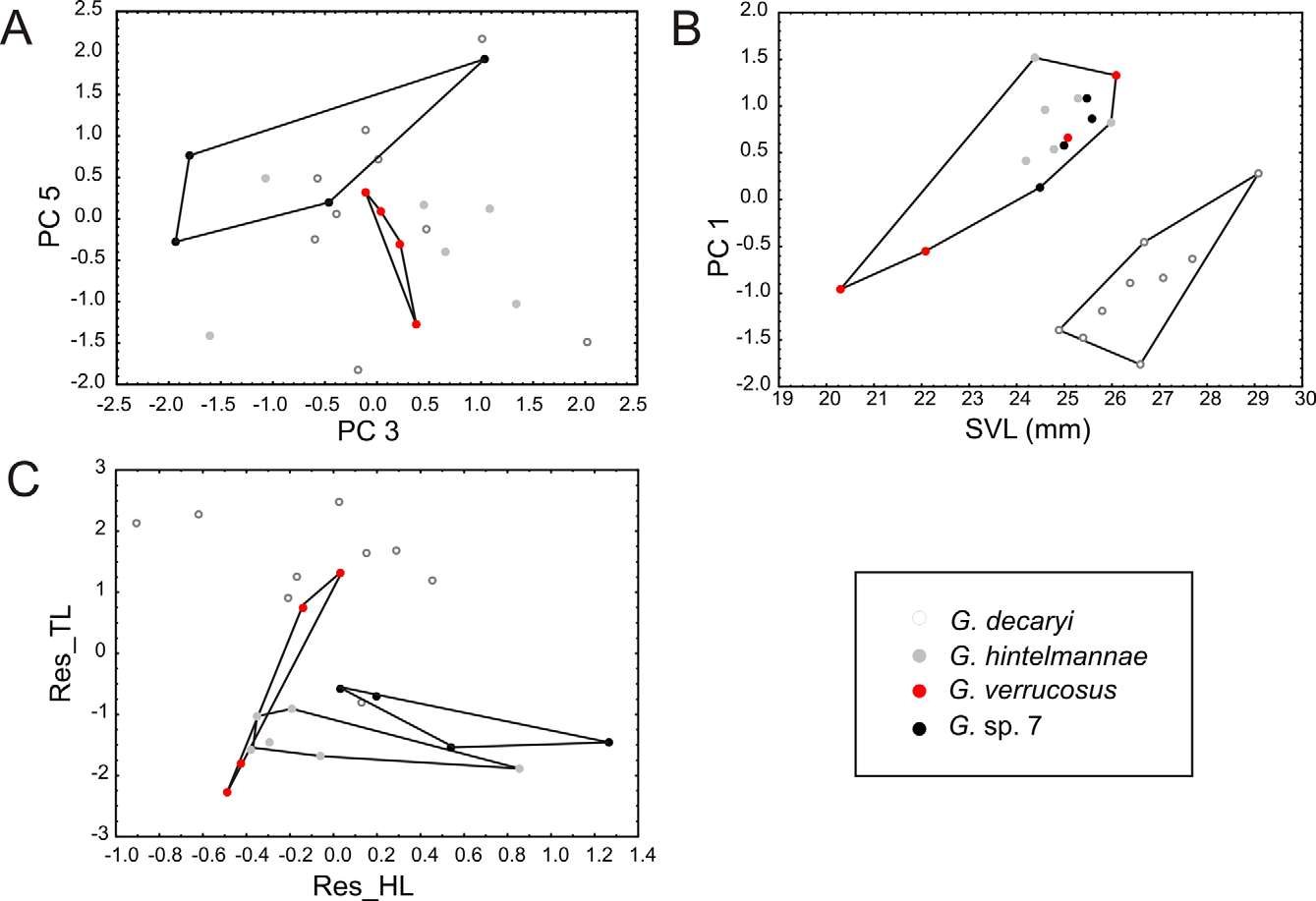
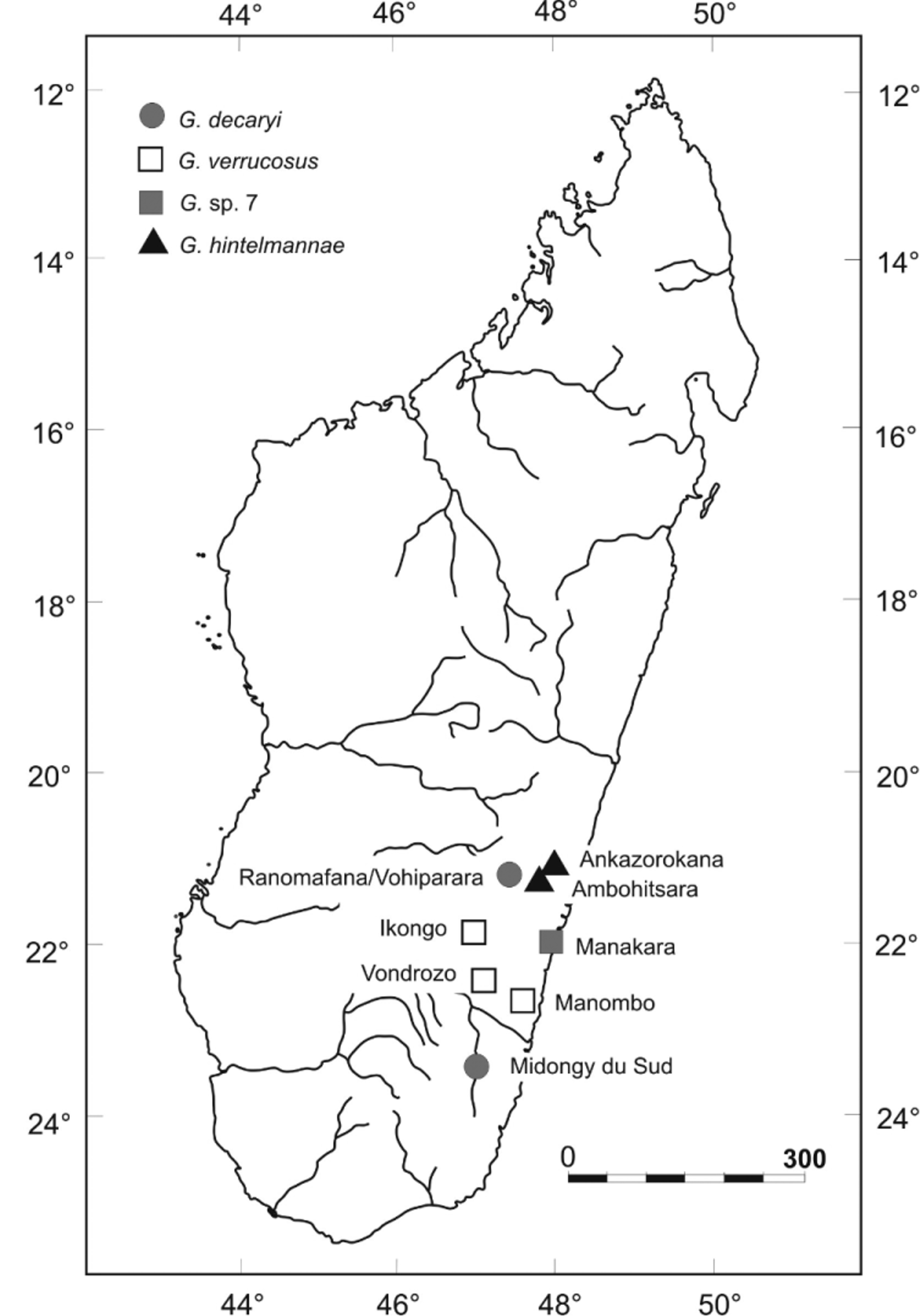
Identity. Gephyromantis decaryi was described by Angel (1930) on the basis of four specimens collected by R. Decary from forest near Midongy du Sud (= Midongy Atsimo) at 500–700 m a.s.l. ( MNHN 1930.435) and a forest at Pic d'Ivohibe (Andringitra Massif, ca. 1100 m a.s.l.) in the South East of Madagascar ( MNHN 1930.436-438). Blommers-Schlösser & Blanc (1991) assigned the species to the genus Mantidactylus and designated the (female) specimen from Midongy ( MNHN 1930.435) as lectotype. As reported in detail by Glaw & Vences (2000), the lectotype is currently in a very poor state of preservation, with dissociated and partially missing hindlimbs. Furthermore, the original description ( Angel 1930) provides a joint description for a total of four syntypes, of which three come from a highland locality (Pic d'Ivohibe) and probably are not conspecific with the lectotype. To assign this lectotype specimen and thus the nomen decaryi to any of the species that we have identified by molecular and bioacoustic data, we therefore can only use indirect evidence. The coloration characters mentioned by Glaw & Vences (2000) and derived from their examination of the lectotype are rather unspecific and in principle could apply to any of the species identified by us. There are two lines of evidence, however, supporting the species definition of Glaw & Vences (2000): (1) The lectotype upon examination in 1999 had a slightly converging dark-bordered ridge on each side of the neck that is followed dorsolaterally by a further ridge. This character is most clearly expressed in the specimens from the Ranomafana region, although also some specimens from Manombo have irregular expressions of such ridges. (2) The original description ( Angel 1930) describes G. decaryi as a species with long hindlimbs, with the tibiotarsal articulation widely surpassing the snout tip when limbs are adpressed along body. Although this was probably written mainly referring to the paralectotype specimen MNHN 1930.437, a male with very long hindlimbs (see Glaw & Vences 2000), a comparatively large hindlimb length also appears to characterize the lectotype. Its poor state of preservation did not allow to take a full range of measurements, but we here give a number of measurements of the lectotype ( MNHN 1930.435) kindly provided by A. Miralles who examined the specimens in 2010 (all in mm): SVL 22.1, FOTL (only left hindlimb) 22.0, TL (on left and right hindlimb) 14.4–14.7 ( Fig. 12 View FIGURE 12 ). Relative FOTL and TL (as FOTL /SVL and TL/SVL ratios) are thus 1.0 and 0.65–0.67, in perfect agreement with the values of specimens from Ranomafana National Park assigned to G. decaryi : 0.87–0.96 for relative FOTL, and 0.59–0.69 for relative TL. In comparison, relative FOTL is <0.79 and relative TL is <0.57 in the other three (candidate) species of the G. decaryi complex (calculated from original values in Table 1). Therefore, the long hindlimbs of the G. decaryi lectotype agree most congruently with the species from Ranomafana and we therefore propose to continue assigning this name to the Ranomafana population. This is the taxonomically most parsimonious solution, but unfortunately implies the biogeographic anomaly of considering the northernmost population from Ranomafana to be conspecific with G. decaryi whose lectotype was collected at the southernmost locality known for the complex (see Fig. 10 View FIGURE 10 ). We hypothesize that in fact, G. d e c a r y i might in the future prove to be yet a different species (and the Ranomafana populations thus represent an undescribed species), but for such a conclusion, fresh material associated to molecular and bioacoustic data from Midongy du Sud will be needed.
Remark. Vieites et al. (2009) and Wollenberg et al. (2011) considered the Ranomafana population as candidate species Gephyromantis sp. 9, and used the name G. decaryi for populations from Vondrozo and Manombo which geographically are closer to the type locality of G. decaryi . However, after re-examination of the type material, we here confirm the view of Glaw & Vences (2000) and assign the name decaryi to the Ranomafana population as was also done by Kaffenberger et al. (2012). A detailed redescription of this species has already been provided by Glaw & Vences (2000).
Material examined. ZSM 2495/2007 ( ZCMV 5203), ZSM 2496/2007 ( ZCMV 5222), ZSM 2497/2007 ( ZCMV 5223), ZSM 2498/2007 ( ZCMV 5589), four adult males, collected in the vicinities of Valbio research station, south-eastern Madagascar, 21°15,191'S, 47°25,271'E, by K. C. Wollenberg, P. Nürnberger, E. Rajeriarison and T. Rajoafiarison on 26 February, 1 March, 2 March, and 24 February 2007, respectively. ZSM 1850/2008 ( ZCMV 8153), adult male, collected near Ambatolahy village, close to Ranomafana National Park, at a site with geographical coordinates 21° 14,897'S, 47° 25,800'E, 948 m a.s.l., by K. C. Wollenberg on 16 February 2008; and four adult males and one probable female to be catalogued in UADBA and here represented by their field numbers only: ZCMV 5353, collected in the vicinities of Valbio research station by K. C. Wollenberg, P. Nürnberger, E. Rajeriarison and T. Rajoafiarison; ZSM 1849/2008 ( ZCMV 8152), ZSM 1851/2008 ( ZCMV 8154), ZSM 1852/2008 ( ZCMV 8187), and ZSM 1853/2008 ( ZCMV 8188) collected near Ambatolahy by K. C. Wollenberg on 15–16 February 2008.
Diagnosis. A member of the subgenus Gephyromantis in the genus Gephyromantis on the basis of (1) presence of intercalary elements between ultimate and penultimate phalanges of fingers and toes (verified by external examination), (2) small size (SVL below 35 mm), (3) slightly enlarged terminal discs of fingers, (4) presence of outer metatarsal tubercle, (5) absence of webbing on hands and presence of only rudimentary webbing on feet, (6) tight connection of tissue surrounding the two lateral metatarsalia, (7) presence of femoral glands in males ( Fig. 9 View FIGURE 9 ), (8) presence of paired or bilobed blackish vocal sacs in males, (9) diurnal calling behavior not restricted to water bodies.
Within the subgenus, distinguished from all other species by combination of (1) comparatively moderate size (male SVL 25–28 mm), (2) dorsum with several larger tubercles; usually with presence of distinct inner and outer dorsolateral ridges which usually are continuous over a significant stretch and not merely rows of elongated tubercles, (3) upper lip with light gray (but not white or yellow) color interrupted by 2–4 dark brown markings, (4) lower lip ventrally without a distinct alternating series of brown and yellowish markings; (5) body slender, forelimbs and hindlimbs very long (tibiotarsal articulation reaching beyond snout tip when hindlimbs are adpressed along body), (6) calls consisting of a rather long series of>30 notes of indistinctly pulsed structure with a low note repetition rate of 3.6–4.6/s.
Assigned to the group of species here informally defined as the Gephyromantis decaryi complex based on its molecular phylogenetic relationships (see above) and its advertisement call characters namely long series of notes with a low note repetition rate. For a morphological and bioacoustic comparison to the newly described species in the complex see the diagnoses of these species below, Table 2 View TABLE 2 and Fig. 5 View FIGURE 5 .
Natural history. Specimens were regularly observed calling from shrubs and fern vegetation at the edge of the rainforest. They were common in the vegetation along the main road near Valbio biological station and the entrance of Ranomafana National Park. Calling males were heard from low perches of ca. 10 cm in ferns, but also from higher perches of 100–150 cm above the ground. They typically called during the day, but calls were also heard at night. Reproduction and larval stages are unknown but the fact that calling males are not concentrated close to water bodies suggests a possible nidicolous reproduction as in other species of the subgenus Gephyromantis .
Advertisement calls. Glaw & Vences (2000) described the call of Gephyromantis decaryi (under the name Mantidactylus decaryi ) based on recordings from 26 February 1996 at Ranomafana. Temporal data from this description are reproduced in Table 2 View TABLE 2 . One call consisted of 44 notes, each peaking in intensity at its beginning. Fundamental frequency was 1450–1650 Hz, dominant frequency was 2950–3600 Hz.
Own recordings from the campsite next to Valbio biological station (from specimen ZCMV 5585) largely agree with this previous description (see Table 2 View TABLE 2 for temporal call data). Note repetition rate was 4.6/s. The notes had a very indistinctly pulsed structure, with 14–16 intensity peaks per note (n=3) that can be regarded as pulses. A fundamental frequency band was present at 1800–2000 Hz, dominant frequency ranges between 3000–4200 Hz, some low intensity components seemed to reach up to 8000 Hz.
Distribution. So far known from two geographically quite distant localities, Midongy du Sud (the type locality) and Ranomafana National Park. In Ranomafana, the species is locally common at mid-elevations (ca. 600–1050 m a.s.l.), but clearly has a more fragmented area of occupancy than other species of Gephyromantis , such as G. enki .


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.