
Squalus lobularis, De, Sarah T., De, Marcelo R. & Gomes, Ulisses L., 2016
|
publication ID |
https://doi.org/10.11646/zootaxa.4133.1.1 |
|
publication LSID |
lsid:zoobank.org:pub:E7276A63-67C8-4BC5-8419-2EBDAE4432B0 |
|
DOI |
https://doi.org/10.5281/zenodo.6075723 |
|
persistent identifier |
https://treatment.plazi.org/id/282F878E-FF97-FFA3-14C0-2650FDB65D51 |
|
treatment provided by |
Plazi |
|
scientific name |
Squalus lobularis |
| status |
sp. nov. |
Squalus lobularis View in CoL sp. nov.
( Figs. 16–24 View FIGURE 16 View FIGURE 17 View FIGURE 18 View FIGURE 19 View FIGURE 20 View FIGURE 21 View FIGURE 22 View FIGURE 23 View FIGURE 24 ; Tables 3 View TABLE 3 , 5–6 View TABLE 5 View TABLE 6 )
Suggested common names: Atlantic lobefin dogfish; Cação-bagre-de-nadadeiras-lobadas (Portuguese).
Squalus cubensis View in CoL (not Howell-Rivero): Figueiredo, 1977: 8, fig. 7 (description; Southern Brazil and Uruguay).
Squalus View in CoL sp. of the blainvillei View in CoL group: Figueiredo, 1981: 17 (listed; Brazil); Gomes et al., 1997: 94 –95 (listed; Brazil); Marques, 1999 (description; Brazil).
Squalus mitsukurii View in CoL (not Jordan & Snyder): Calderón, 1994: 1 –43, fig. 5b (cited; Brazil); Lessa et al., 1999: 61, 150 (cited, listed; South Brazil); Lamilla & Bustamante, 2005: 9, 26 (listed; Chile); Compagno, 2002: 385 (description; Northeast Brazil and Argentina); Compagno et al., 2005: 77 –78, plate 3 (cited; Southeast Brazil).
Squalus View in CoL of the blainvillei View in CoL / mitsukurii View in CoL group: Gadig, 2001 (in part): 29, 36, 54–55, fig. 29 (cited; Brazil).
Squalus View in CoL sp. B: Soto, 2001: 96 (listed; Brazil); Soto & Mincarone, 2004: 79 –82 (listed; Brazil).
Squalus View in CoL sp. 1: Gomes et al., 2010: 44, 45 (cited; Brazil).
Squalus View in CoL sp.: Viana, 2011: 163–185, figs. 78–80, 83, 85, 87–92 (revision; Brazil, Argentina, Uruguay); Naylor et al., 2012: 57, 205, fig. 42 (molecular systematics; Western South Atlantic).
Holotype. HUMZ 91806, juvenile female, 557 mm TL, off Patagonia, Argentina, 3930’S, 5828’W. Unknown collector. Collecting date 16 January 1979.
Paratypes ( 9 specimens). HUMZ 30026, adult female, 725 mm TL, off Patagonia, Argentina, 4235’S, 6311’W; HUMZ 30032, adult female, 675 mm TL, off Patagonia, Argentina, 4235’S, 6311’W; HUMZ 91801, adult female, 650 mm TL, off Patagonia, Argentina, 3930’S, 5828’W; HUMZ 91804, juvenile female, 545 mm TL, off Patagonia, Argentina, 3930’S, 5828’W; HUMZ 91807, juvenile female, 605 mm TL, off Patagonia, Argentina, 3930’S, 5828’W; UERJ 1661, adult male, 640 mm TL, unknown locality (dissected), Southeast Brazil; UERJ 2024, juvenile female, 530 mm TL, off Espírito Santo, Brazil; UERJ 2025, adult female, 600 mm TL; same locality as UERJ 2024. UERJ 2026, adult female, 675 mm TL, same locality as UERJ 2024.
Non-type material ( 32 specimens). FURG 80.0607, juvenile female, 510 mm TL, off Rio Grande do Sul coast, Brazil, 3219'S, 5152'W; FURG 80.0609, adult male, 630 mm TL, South of Mar del Plata, Argentina, 38°S, 51°W; FURG 98.0031, two juvenile males, 380–450 mm TL, off Rio Grande do Sul coast, Brazil, 3219'S, 5152'W; MZUSP 37351, neonate female, 240 mm TL, juvenile male, 340 mm TL, Uruguay, 3550'S, 5306'W; MZUSP 37355, two juvenile female, 335–340 mm TL; three juvenile male, 335–360 mm TL, Uruguay, 3544'S, 5322'W; UERJ 1112, adult male, 635 mm TL, Rio Grande do Sul, Brazil (dissected); ZMH 104558, adult male, 600 mm TL, Argentina, 380'S, 5633'W; ZMH 104707, juvenile male, 247 mm TL, Argentina, 3649'S, 5437'W; ZMH 104945, neonate female, 235 mm TL, Argentina, 4216'S, 6153'W; ZMH 107897, adult male, 605 mm TL, Argentina, 400'S, 590'W.
Diagnosis. A species of Squalus from SWAO that differs from its congeners by the combination of having markedly broad and lobe-like dorsal fins and short clasper groove, not reaching the rhipidion. Squalus lobularis sp. nov. is clearly distinct from S. mitsukurii from Japan, its morphologically closest congener, by having a slender body ( vs. highly robust body in S. mitsukurii ), pectoral-fin posterior margin concave ( vs. pectoral-fin posterior margin straight in S. mitsukurii ), narrower interorbital space (8.8%, 6.8%–8.5% TL vs. 9.3%, 9.1%–9.8% TL in S. mitsukurii ); and inner clasper length 1.4 times pelvic-fin inner margin length ( vs. 0.7–1.1 times in S. mitsukurii ). Squalus lobularis sp. nov. further differs from species of the Squalus megalops group by: snout relatively long vs. snout short; pectoral fins markedly broad vs. pectoral fins often very narrow; pectoral free rear tips rounded and lobe-like vs. pectoral free rear tips often triangular; dermal denticles tricuspidate vs. lanceolate. It further differs from all its congeners by having a much larger pectoral-inner margin length (10.5%, 9.2%–11.0% TL), except from S. acanthias .
Description. External morphology. Measurements and meristic data are summarized in Tables 5–6 View TABLE 5 View TABLE 6 . Body fusiform, slender and depressed through, with head height 0.9 (0.8–1.3) times trunk height and 1.0 (0.8–1.2) times abdomen height; greatest width of body at head with head width 1.3 (1.0–1.3) times trunk width and 1.6 (1.2–1.7) times abdomen width. Head flattened dorsally and elongate, its length 22.7% (21.5%–24.3% TL); head broader at mouth than at nostrils (width at mouth 12.2%, 9.7%–12.2% TL vs. width at nostrils 7.5%, 6.3%–7.4% TL). Snout very large (preorbital length 7.9%, 7.0%–7.9% TL), obtuse and rounded at tip; anterior margin of nostrils strongly bilobate, positioned more laterally; prenarial length 1.2 (1.0–1.2) times distance from nostril to upper labial furrow; prenarial length 1.3 (1.1–1.4) times eye length; internarial space 1.1 (0.9–1.5) times eye length. Eyes oval, placed laterally closer to the snout tip than first gill slit, with anterior margin convex and posterior margin notched; eyes enlarged, their length 2.3 (1.7–2.9) times their height. Prespiracular length 1.6 (1.6–1.7) times preorbital length. Spiracles subtriangular and conspicuously broad, their length 1.2% (1.0%–1.6% TL), and corresponding to 0.3 (0.3–0.4) times eyes length. Prebranchial length 1.5 (1.5–1.7) times prespiracular length. Gill slits vertical and tall, with fifth gill slit 1.2 (1.0–1.6) times higher than first gill slit.
Preoral length 9.7% (8.6%–9.9% TL), corresponding to 1.3 (1.1–1.4) times mouth width. Mouth somewhat arched, markedly broad, its width 1.7 (1.5–2.0) times internarial width and 1.4 (1.4–1.7) prenarial length; upper labial furrow elongate, its length 2.4% (2.0%–2.6% TL), with a slender fold; lower labial furrow conspicuously deep and elongate, reaching laterally on head, without a fold. Unicuspid teeth, similar in both jaws, markedly broad at crown, flattened labial-lingually and alternating; upper teeth smaller and narrower than lower teeth; cusp thick and short, oblique and directed laterally; mesial cutting edge conspicuously convex; mesial heel pointed; distal heel markedly rounded; apron thick, although larger on lower teeth than upper teeth; two series of functional teeth in upper and lower jaws; tooth rows varying from 13–13 (13–13 paratypes) in upper jaw and 11–10 (12–11 paratypes) in lower jaw ( Fig. 17 View FIGURE 17 ).
Pre-first dorsal fin length 1.4 (1.3–1.5) times prepectoral length; distance from first dorsal fin origin to pectoral fin origin 1.2 (0.8–1.4) times preorbital length. Origin of first dorsal fin prior to pectoral free rear tips. First dorsal fin markedly broad at fin web, lobe-like, and elongate (its length 1.9, 1.6–2.0 times its height); first dorsal fin tall, its height 7.6%, 6.5%–8.1% TL, corresponding to 1.0 (0.9–1.1) times preorbital length and 1.1 (1.0–2.2) times its inner margin length; first dorsal-fin anterior margin evidently convex and elongate, its length 11.1% (10.3%–11.9% TL); its posterior margin almost straight, its length 9.6% (6.7%–9.2% TL) ( Fig. 18 View FIGURE 18 ); first dorsal-fin apex strongly rounded and its free rear tip triangular; inner margin large, its length 6.6% (3.3%–7.1% TL). First dorsal-fin spine somewhat broad at base (width at base 0.7%, 0.6%–0.9% TL) and short (its length 3.8%, 2.2%–4.0% TL), reaching half of fin height. First dorsal fin 1.1 (1.0–1.2) times larger than second dorsal fin. Interdorsal space short, its length 24.6% (21.9%–24.1% TL), and corresponding to 1.1 (1.0–1.2) times prepectoral length and 2.4 (2.1–2.3) times dorsal-caudal space. Second dorsal fin also broad at fin web, lobe-like, and low with its height 1.0 (0.6–1.4) times its inner margin length; anterior margin markedly convex and elongate (length 11.1%, 9.2%–11.6% TL); its posterior margin conspicuously falcate, its length 5.5% (4.1%–6.4% TL); second dorsal-fin apex rounded and free rear tip triangular; its inner margin very large, its length 5.8% (2.8%–6.1% TL). Second dorsal-fin spine slender and conspicuously elongate, reaching fin apex, its length 0.9 (0.7–1.4) times second dorsal-fin height and 1.4 (1.0– 1.6) times larger than first dorsal-fin spine ( Fig. 18 View FIGURE 18 ).
Pre-pectoral length 0.7 (0.7–0.8 paratypes) times pre-first dorsal length and 0.4 (0.4–0.5 paratypes) times prevent length. Pectoral fin markedly broad posteriorly (posterior margin length 11.2%, 9.7%–11.7% TL) and elongate, its anterior margin length 16.0%, 13.8%–17.2% TL; anterior margin length 1.5 (1.5–1.8) times inner margin length; posterior margin length 0.9 (0.9–1.8) times trunk height; anterior and inner margins rather straight, posterior margin concave; apex strongly rounded and lobe-like; free rear tips also rounded and lobe-like reaching the horizontal line traced at pectoral fin apex.
Squalus lobularis sp. nov. Squalus mitsukurii
Measurements Holotype Holotype Paratypes HUMZ 91806 N Range x SD SU 12793 N Range x SD TL (mm) 557.0 8 530.0 – 725.0 637.5 59.3 710.0 5 240.0 – 770.0 349.6 235.0 PCL 78.5 8 78.3 – 80.5 79.5 0.8 77.5 5 76.9 – 78.6 77.6 0.7 PD2 61.0 8 60.0 – 64.1 61.3 1.4 61.0 5 58.4 – 60.5 59.7 0.9 PD1 31.1 8 29.8 – 31.1 30.4 0.5 32.4 5 30.2 – 31.3 30.8 0.4 SVL 47.0 8 46.9 – 51.9 49.5 1.8 50.0 5 47.3 – 48.9 48.1 0.6 PP2 44.9 8 44.6 – 48.9 46.6 1.5 47.9 5 45.3 – 47.6 46.3 1.0 PP1 22.0 8 20.5 – 23.4 21.8 0.9 24.6 5 22.1 – 24.7 22.9 1.1 HDL 22.7 8 21.5 – 24.3 22.2 0.9 24.2 5 22.7 – 24.4 23.3 0.7 PG1 19.2 8 17.7 – 20.3 18.8 0.9 20.4 5 19.9 – 20.9 20.3 0.4 PSP 12.6 8 11.5 – 12.7 12.1 0.4 12.8 5 12.7 – 21.5 14.9 3.7 POB 7.9 8 7.0 – 7.9 7.4 0.3 7.3 5 7.2 – 8.8 7.8 0.6 PRN 5.4 8 4.5 – 5.3 4.7 0.3 5.6 5 4.7 – 6.4 5.4 0.7 POR 9.7 8 8.6 – 9.9 9.4 0.4 10.3 5 8.9 – 11.5 10.2 0.9 INLF 4.5 8 3.6 – 4.4 4.1 0.3 4.3 5 4.4 – 5.3 4.7 0.4 MOW 7.4 8 7.0 – 8.1 7.6 0.3 8.6 5 7.6 – 8.1 7.8 0.2 ULA 2.4 8 2.0 – 2.6 2.4 0.2 2.5 5 2.1 – 2.7 2.5 0.2 INW 4.4 8 3.8 – 4.9 4.2 0.4 4.7 5 4.0 – 4.5 4.2 0.2 INO 8.8 8 6.8 – 8.5 8.0 0.6 9.3 5 9.1 – 9.8 9.5 0.3 EYL 4.1 8 3.1 – 4.4 3.9 0.5 3.6 5 3.1 – 4.6 3.8 0.5 EYH 1.8 8 1.4 – 2.3 1.7 0.3 0.9 5 1.2 – 2.5 1.7 0.5 SPL 1.2 8 1.0 – 1.6 1.4 0.2 1.3 5 1.3 – 1.5 1.4 0.1 GS1 1.9 8 1.4 – 1.9 1.7 0.2 1.7 5 1.7 – 2.3 2.0 0.2 GS5 2.2 8 1.9 – 2.4 2.1 0.2 2.3 5 1.8 – 2.7 2.3 0.3 IDS 24.6 8 21.9 – 24.1 23.3 0.9 21.1 5 21.9 – 24.7 23.1 1.1 DCS 10.1 8 10.4 – 11.5 10.7 0.4 10.6 5 8.1 – 11.9 10.4 1.5 PPS 18.9 8 20.3 – 24.4 22.2 1.4 21.8 5 18.2 – 23.4 20.1 2.1 PCA 26.0 8 24.8 – 28.1 26.0 1.1 23.7 5 23.0 – 26.2 24.7 1.3 D1L 14.4 8 11.8 – 14.1 13.2 0.7 13.6 5 11.8 – 14.5 13.5 1.3 D1A 11.1 8 10.3 – 11.9 11.2 0.6 12.0 5 10.2 – 13.2 12.0 1.3 D1B 7.5 8 7.1 – 8.4 7.8 0.5 8.2 5 5.3 – 8.8 7.4 1.5 D1H 7.6 8 6.5 – 8.1 7.3 0.5 9.8 5 7.0 – 9.8 8.0 1.2 D1I 6.6 8 3.3 – 7.1 5.7 1.2 6.2 5 6.1 – 6.5 6.3 0.1 D1P 9.6 8 6.7 – 9.2 8.0 0.9 9.3 5 7.0 – 10.6 8.7 1.3 D1ES 3.8 7 2.2 – 4.0 3.3 0.6 3.9 5 2.1 – 5.4 2.9 1.4 D1BS 0.7 8 0.6 – 0.9 0.8 0.1 1.0 5 0.5 – 1.0 0.7 0.2 D2L 13.7 8 11.3 – 13.0 12.3 0.7 12.3 5 11.6 – 12.6 12.0 0.4 D2A 11.1 8 9.2 – 11.6 10.0 0.8 10.2 5 9.9 – 11.6 10.5 0.7 D2B 8.2 8 6.8 – 9.2 7.5 0.8 7.2 5 6.2 – 7.1 6.6 0.4 D2H 6.0 8 3.7 – 5.6 4.7 0.8 6.8 5 4.8 – 7.9 6.0 1.2
Paratypes
Pelvic fins somewhat wide with anterior margin slightly concave; inner and posterior margins straight; free rear tips and apex rounded, the former conspicuously lobe-like; pelvic fin length 10.4% (9.9%–11.6% TL); origin of pelvic fins 3.0 (2.9–3.2) times distance between origins of the two dorsal fins, nearer to first dorsal fin than to second dorsal fin (although closer to midline between the origins of two dorsal fins in paratypes); pectoral-pelvic space 0.7 (0.8–0.9) times pelvic-caudal space. Claspers in male paratype cylindrical, markedly thick and compressed dorsoventrally throughout, extending beyond free rear tips of pelvic fins, its outer length 4.3% TL; elongate and wide siphon, placed medioventrally from midline of basipterygium to origin of ventral marginal cartilage; clasper groove longitudinal, short (not reaching the rhipidion) and very constricted, dorsally located; apopyle narrow anteriorly in clasper groove; hypopyle also narrow, located posteriorly in clasper groove; rhipidion conspicuously large, blade-like and thin, positioned medially at distal end of clasper ( Fig. 19 View FIGURE 19 ).
Lateral keels well demarked on each side of caudal peduncle with their origin behind insertion of second dorsal fin; upper and lower precaudal furrows deep. Caudal fin subrectangular with dorsal caudal margin straight and large, its length 21.7% (19.7%–22.0% TL); dorsal caudal margin length 1.0 (0.8–1.0) times head length and 1.8 (1.7–2.0) times greater than preventral caudal margin length; posterior caudal tip rounded; both postventral caudal margins convex; upper caudal lobe narrow, its width at caudal fork 6.9% (6.3%–7.4% TL); caudal fork between lobes strongly concave; preventral caudal margin convex and elongate, its length 11.8% (10.6%–12.2% TL) and corresponding to 2.2 (2.0–2.7) times pelvic inner margin length; ventral caudal tip rounded ( Fig. 20 View FIGURE 20 ).
Dermal denticles ( Fig. 21 View FIGURE 21 ). Denticles tricuspid with cusps conspicuous and pointed; median projection anteriorly on crown prominent and rounded with small expansions posterolaterally on each side, and crown furrow medially; lateral cusps much shorter than median cusp; lateral ridges prominent and bifurcate, reaching tips of lateral cusps; median ridge conspicuous and tall with its origin far anteriorly at crown base and reaching the tip of median cusp distally; dermal denticles markedly broad at crown, although their length slightly greater than their width; dermal denticles imbricated and adjacent to each other. Dermal denticles also tricuspid in juveniles, although much smaller than in adults and not imbricated; their cusps markedly slender and pointed with lateral cusps forming a prominent concavity with a median cusp on each side.
Coloration ( Figs. 16 View FIGURE 16 , 20 View FIGURE 20 ). Body darkish gray dorsally and pale ventrally. Dorsal fins also dark gray, whitish at fin base; first dorsal fin slightly white at posterior margin and free rear tip, darker at apex and upper half of posterior margin; second dorsal fin also dark gray with posterior margin white and apex slightly blackish; dorsal spines gray, blackish anteriorly, white posteriorly and at tips. Pectoral fins dark gray dorsoventrally and pale under pectoral radials; pectoral-fin posterior margin markedly white; pectoral-fin apex broadly white; pectoral-fin inner margin light gray. Pelvic fins gray dorsally and light gray ventrally; both pelvic anterior and posterior margins white. Caudal fin also dark gray and whitish over vertebral column; dorsal caudal margin somewhat whitish, although not uniform; posterior caudal tip white; upper postventral caudal margin white, except for dark gray caudal fork; lower postventral caudal margin also white; ventral caudal tip broadly white; preventral caudal margin slightly white; black caudal stripe conspicuous in dorsal lobe ( Fig. 20 View FIGURE 20 ). Juveniles with body much darker than adults.
Skeletal morphology. Measurements and meristic data are summarized in Tables 3 View TABLE 3 , 6 View TABLE 6 .
Neurocranium ( Fig. 22 View FIGURE 22 ). Rostrum spoon-shaped, thick and elongate (precerebral fossa 40.6% CL), narrow anteriorly and broad posteriorly; lateral rostral cartilages somewhat cylindrical, although compressed more anteriorly; precerebral fossa markedly profound and broad, its width 13.6% CL; three conspicuous rostral appendices ventrally with median rostral appendix shorter than lateral appendices; prefrontal fontanelle rounded and narrow, placed dorsally and posterior to the precerebral fossa at the base of rostrum; rostral keel strongly prominent and elongate, its length 20.8% CL, reaching posterior margin of subnasal fenestra. Nasal capsules markedly rounded and broad (width across nasal capsules 55.2% CL); subnasal fossa vertical, large, placed ventrally on each side of rostral keel.
Cranial roof strongly concave medially and delimited by preorbital processes anteriorly, supraorbital crest laterally, and postorbital processes posteriorly; its greatest width at orbital processes (width across preorbital processes 52.3% CL; width across postorbital processes 55.8% CL), clearly narrow and concave in the interorbital space (interorbital width 30.2% CL); preorbital processes inconspicuous; postorbital processes triangular and small, its length 6.9% CL; supraorbital crest prominent laterally with a deep longitudinal sulcus located at its base, carrying a series of eight foramina for the superficial ophthalmic branches of trigeminal (V) and facial (VII) nerves; preorbital canal markedly broad and rounded, placed anteriorly to the series of foramina; profundus canal for ophthalmicus profundus nerve rounded, located anteriorly to the preorbital canal in the longitudinal sulcus; supraethmoidal processes conspicuous, cylindrical and elongate, placed mediodorsally at the base of the ethmoidal region; ethmoidal canal rounded and wide, although narrower than the preorbital canal, placed dorsoanteriorly at the lateral base of ethmoidal region; epiphyseal pit rounded and wide, located medially behind supraethmoidal processes; ectethmoid process prominent, slightly pointed posteriorly on each side; subethmoidean chamber small, broader anteriorly than posteriorly (its width 15.3% TL); subethmoidal ridge follows rostral keel ventrally in the subethmoidean region.
Otic region comprised by two oval otic capsules on each side, separated by deep endolymphatic fossa; each otic capsule delimited by anterior semicircular canal anteriorly, posterior semicircular canal posteriorly, sphenopterotic ridge dorsolaterally, and lateral semicircular canal laterally which is dorsal to the hyomandibular facet; endolymphatic fossa rounded, placed medially in otic region, with two anterior endolymphatic foramina and two posterior perilymphatic foramina, with the later much broader than the former; otic crest prominent located medioposteriorly, running from posterior margin of the endolymphatic fossa to foramen magnum; opisthotic process conspicuous posteriorly in the otic capsule; lateral auditory groove shallow, placed afore the hyomandibular facet; hyomandibular facet profound and broad, located lateroventrally in otic region.
Interorbital wall wide and profound, delimited by preorbital wall anteriorly, otic capsule posteriorly, supraorbital crest dorsally and groove for orbital process ventrally; preorbital wall concave, with a single orbitonasal canal at its base; optic foramen (II) conspicuously broad, placed midventrally in interorbital wall near groove for orbital process; troclear foramen (IV) tiny, located middorsally just below supraorbital crest; eye-stalk prominent, posteriorly in interorbital wall anterior to prooticum foramen (V, VII); oculomotor foramen (III) and abducens foramen (VI) somewhat wide, placed dorsal and ventrally to eye-stalk, respectively; prooticum foramen for the trigeminal (V) and facial (VII) nerves with markedly wide aperture, placed ventroposteriorly in interorbital wall, just anterior to postorbital process; two apertures for hyomandibular branch of the facial nerve (VII) opening more posteriorly at base of hyomandibular facet; transbasal canal for the pituitary vein clearly wide, placed ventral to eye-stalk; foramen for efferent branch of pseudobranchial artery broad, located just anterior to transbasal canal. Occipital region with two triangular occipital condyles ventrally; glossopharyngeal bases strongly conspicuous, subtriangular and thick, located laterally and with a broad foramen for glossopharyngeal nerve (IX) at base; foramen for vagus nerve (X) rounded and wide, medial to foramen for glossopharyngeal nerve (IX), lateral to occipital condyles; foramen magnum broad, its width 8.8% CL.
Basal plate flattened and elongate, its length 42.4% CL, narrow anteriorly at basal angle (its width 18.1% CL) and wide posteriorly at prootic process (width across hyomandibular facets 43.5% CL); basitrabecular process conspicuous and bean-shaped; lateral sinuosity prominent between basitrabecular process and first cartilaginous process; first cartilaginous process conspicuous and triangular, with a single foramen for orbital artery at its base on each side of basal plate; single foramen for carotid artery anteromedially placed at level of lateral sinuosity.
Pelvic fin and girdle ( Fig. 23 View FIGURE 23 ). Puboischiadic bar transverse, rectangular, somewhat thick, especially at sides, biconvex medially and biconcave laterally; lateral prepelvic process rounded evident on each side of the puboischiadic bar; single foramen for pelvic nerve present lateroanteriorly in the puboischiadic bar; three prominent expansions lateroposteriorly on puboischiadic bar for pelvic fin articulation. Basipterygium vertical, elongate, concave medially and convex laterally; anterior pelvic basal small, its length not reaching one-third the basipterygium length, and subrectangular, with four tiny series of irregular radials; pelvic radials slender and elongate, segmented into proximal and distal elements, the former larger than the later; 15 total pelvic radials.
Claspers ( Fig. 23 View FIGURE 23 ). Intermediate element barrel-shaped, connecting pelvic fin basipterygium to axial cartilage; single beta cartilage evident dorsally, thin and cylindrical, somewhat small, its length not reaching one-third the dorsal terminal cartilage (or claw) length, placed laterodorsally for attaching pelvic fin basipterygium to axial cartilage. Axial cartilage elongate and somewhat thick, slightly sinuous; dorsal marginal cartilage inconspicuous, slightly thicker distally, located laterally over axial cartilage; end-style slim and elongate, reaching more than onehalf the length of dorsal terminal cartilage, medially at end of axial cartilage between dorsal marginal cartilage and dorsal terminal cartilage; dorsal terminal cartilage (or claw) clearly slender and elongate, its length more than twothirds axial cartilage length, strongly hook-like with prominent concavity distally and pointed at its tip, connected proximally to dorsal marginal cartilage and axial cartilage, and medially to end-style; dorsal terminal 2 cartilage leaf-like and compressed, noticeably large, connected medially to dorsal terminal cartilage and proximally to dorsal marginal cartilage, supporting the rhipidion. Ventral marginal cartilage inconspicuous, thicker and broader posteriorly than anteriorly, placed distally under the axial cartilage as an evident folded plate laterally; accessory terminal 3 cartilage (or spur) markedly slender and elongate, thicker proximally and distally strongly pointed with single groove present medially, associated to folded plate of ventral marginal cartilage; ventral terminal cartilage conspicuously large, its length more than one-third the length of axial cartilage, spatula-like, rounded at its distal tip and sinuous laterally, placed at the distal end of clasper and connected proximally to ventral marginal cartilage and medially to end-style.
Vertebral counts ( Table 6 View TABLE 6 ). Monospondylous vertebrae 45 in holotype ( 44–48 in paratypes); precaudal vertebrae 88 (80–89); caudal vertebrae 30 (28–31); total vertebrae 118 (110–119).
Geographical distribution ( Fig. 24 View FIGURE 24 ). Squalus lobularis occurs from southern Brazil to Uruguay and Patagonia in Argentina, where it seems to be more common. Its occurrence in the coast of Espírito Santo, Brazil (C. Magenta, pers. comm.), is still doubtful due to lack of precise locality information.
Etymology. From the Latin lobus, in reference to its diagnostic broad and lobe-like dorsal and pectoral fins.
Remarks. Squalus lobularis has previously been misidentified in the SWAO as S. mitsukurii a valid species originally described from Japan ( Fig. 25 View FIGURE 25 ), or as a member of the S. mitsukurii group at least in the SWAO (e.g. Figueiredo, 1981; Lucifora et al., 1999; Marques, 1999; Gomes et al., 1997, 2010). Calderón (1994) recognized S. mitsukurii off the southern Brazilian coast based on morphological characters provided by Compagno (1984), specifically proportional measurements of the head, pectoral and dorsal-fin base lengths, and pelvic-fin length. These characters occur, however, in more than one species of Squalus , which may have led to the misidentification S. mitsukurii in this region ( Fig. 25 View FIGURE 25 ).
Squalus lobularis shares with S. mitsukurii a few morphological characters ( Table 5 View TABLE 5 ): body moderately elongate; snout obtuse and markedly elongate (preorbital length 7.3%, 7.0%–7.4% TL for S. lobularis vs. 7.3%, 7.2%–8.8% TL for S. mitsukurii ); nostrils closer to the mouth than to snout tip (prenarial length 1.0, 1.1–1.2 times inner nostril-labial length vs. 1.3, 1.1–1.4 times for S. mitsukurii ); pectoral fins broad and smaller than head length; dermal denticles tricuspid; and presence of a dark caudal bar. However, this species can be distinguished from S. mitsukurii by the characters provided in its diagnosis above, most notably in having a slender body ( vs. highly robust body in S. mitsukurii ), and concave pectoral-fin posterior margin ( vs. pectoral-fin posterior margin straight in S. mitsukurii ). Despite overlapping, the distance from nostrils to upper labial furrow may be useful for separating them: 4.5%, 3.6%–4.4% TL for S. lobularis vs. 4.3%, 4.4%–5.3% TL for S. mitsukurii . Squalus lobularis also has a shorter prespiracular length (12.6% TL, 11.5%–12.7% TL vs. 12.8% TL, 12.7%–21.5% TL for S. mitsukurii ).
Chen et al. (1979) provided vertebral counts for S. mitsukurii from Japan that are higher than those for S. lobularis despite of some overlapping, respectively: total vertebrae (118–127 vs. 118, 110–119). According to Last et al. (2007), members of S. mitsukurii group share a relatively great total length, presence of dark caudal bar, dorsal spines small, first dorsal fin short and falcate, tricuspidate dermal denticles, pectoral fin not falcate with rounded free rear tips, and high number of vertebrae. Squalus lobularis differs from members of this group by having dorsal spines robust and elongate (first dorsal-fin spine length 3.8% for holotype, 2.2%–4.0% TL for paratypes; second dorsal-fin spine length 5.3%, 3.4%–5.3% TL), as well as first dorsal fin conspicuously broad and lobe-like, not falcate.
A paratype of S. mitsukurii (SU 7184) was not taken into account in our morphometric comparisons because it corresponds to a specimen of S. acanthias clearly identified by the presence of white spots dorsally and first dorsalfin spine located posterior to the pectoral fins, as previously noticed by Bigelow & Schroeder (1957) and Muñoz- Chápuli & Ramos (1989). This type specimen was illustrated in the original description of S. mitsukurii and posteriorly reproduced by Tanaka (1912), but it does not agree with the diagnosis of the Japanese species. Recently, investigations concerning the taxonomy and geographical distribution of S. mitsukurii (e.g. Last et al., 2007) have strongly demonstrated that it might be a complex of species rather than a single valid species (e.g. Bigelow & Schroeder, 1957; Chen et al., 1979; White & Iglésias, 2011).
Squalus lobularis is also distinguished from S. edmundsi , S. grahami , and S. nasutus when compared to data taken from original descriptions provided in Last et al. (2007), by a smaller snout (prenarial length 5.4%, 4.5%– 5.3% TL for S. lobularis ; 5.8%, 5.6%–5.8% TL for S. edmundsi ; 5.6%, 5.7%–6.3% for S. grahami ; and 7.3%, 6.4%–6.7% TL for S. nasutus ). It also differs from S. acanthias from SWAO, S. montalbani , S. nasutus , S. hemipinnis , S. crassispinus , S. megalops , and S. raoulensis by greater total and precaudal vertebrae ( Table 12 View TABLE 12 ). It also differs from S. grahami , S. griffini , S. nasutus , S. hemipinnis , S. crassispinus , S. megalops , S. bucephalus , S. raoulensis , and S. notocaudatus by number of monospondylous vertebrae ( Table 12 View TABLE 12 ).
The recognition of S. lobularis as new species supports that the current acceptance of S. mitsukurii as a cosmopolitan species is probably erroneous, propagated when using the general descriptive literature (e.g. Compagno, 1984) for identifying species regionally. Further analysis of material identified as S. mitsukurii from other localities where it is reported to occur, such as in the Southeastern Atlantic and Western Indian Oceans, and Northeastern Pacific Ocean, is needed to check if this species is indeed widespread or more restricted (e.g. Compagno et al., 2005).
Comparative material. Squalus mitsukurii ( 8 specimens): SU 12793 ( holotype of Squalus mitsukurii ), adult female, 710 mm TL, Honshu Island, Misaki, Japan; SU 12794 ( paratype of S. mitsukurii ) adult male, 770 mm TL, same as SU 12793; SU 7184, paratype of S. mitsukurii (identified as S. acanthias ), neonate male, 277 mm TL, same as SU 12793; SU 7748 ( paratypes of S. mitsukurii ), two neonate females, 240–243 mm TL, neonate male, 247 mm TL, same as SU 12793; AMNH 8822 ( paratype of S. mitsukurii ), neonate female, 248 mm TL, same as SU 12793; USNM 62467, Hawaii, U.S. A..


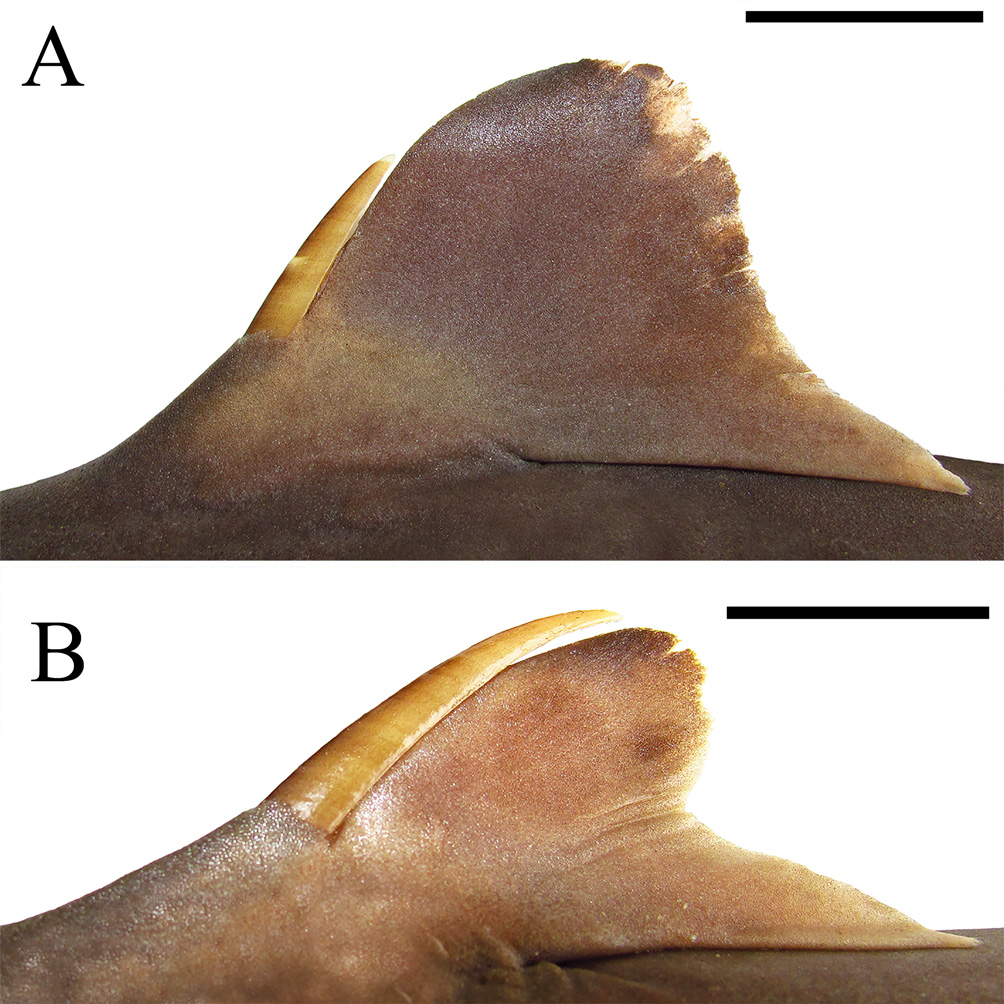



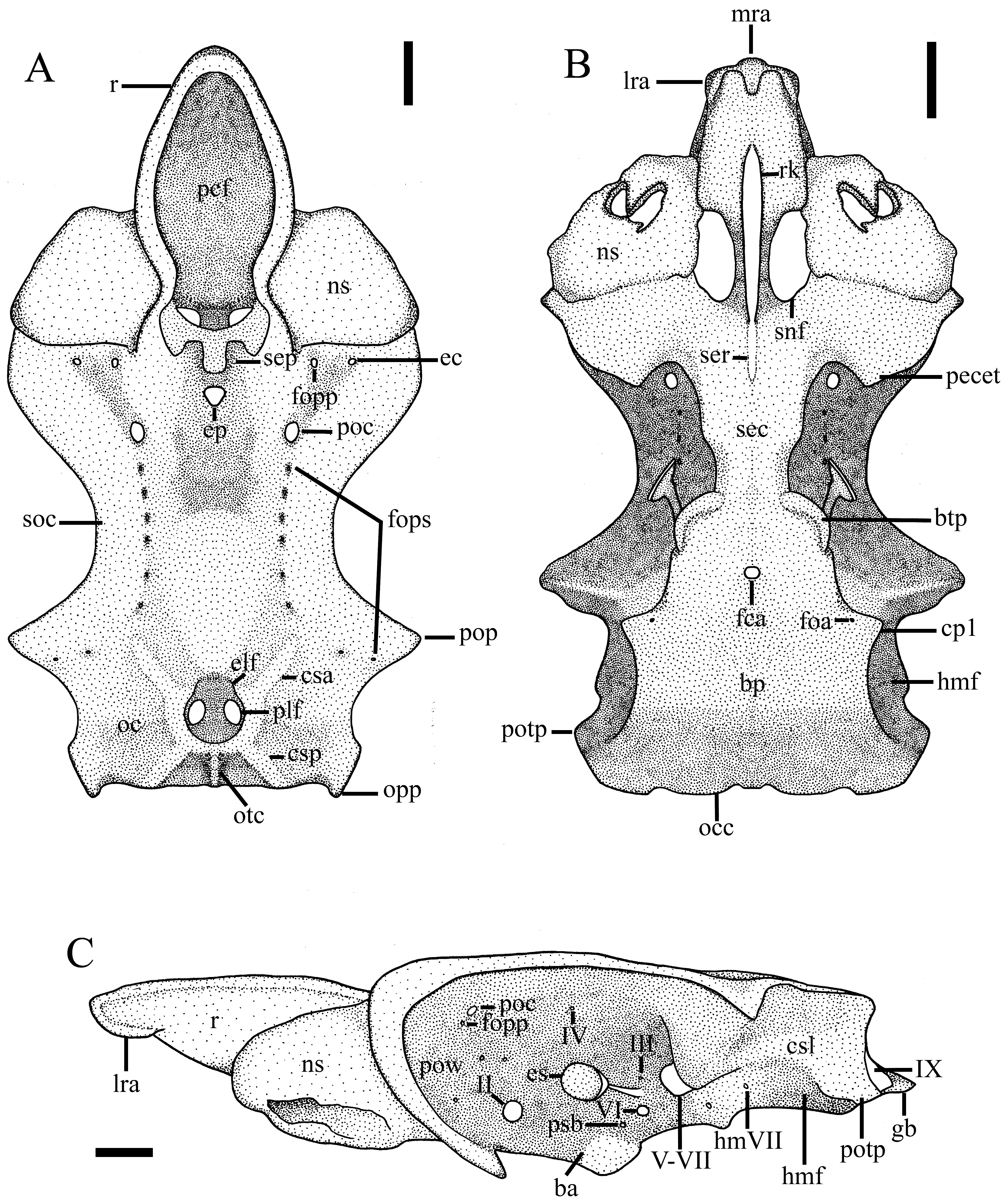

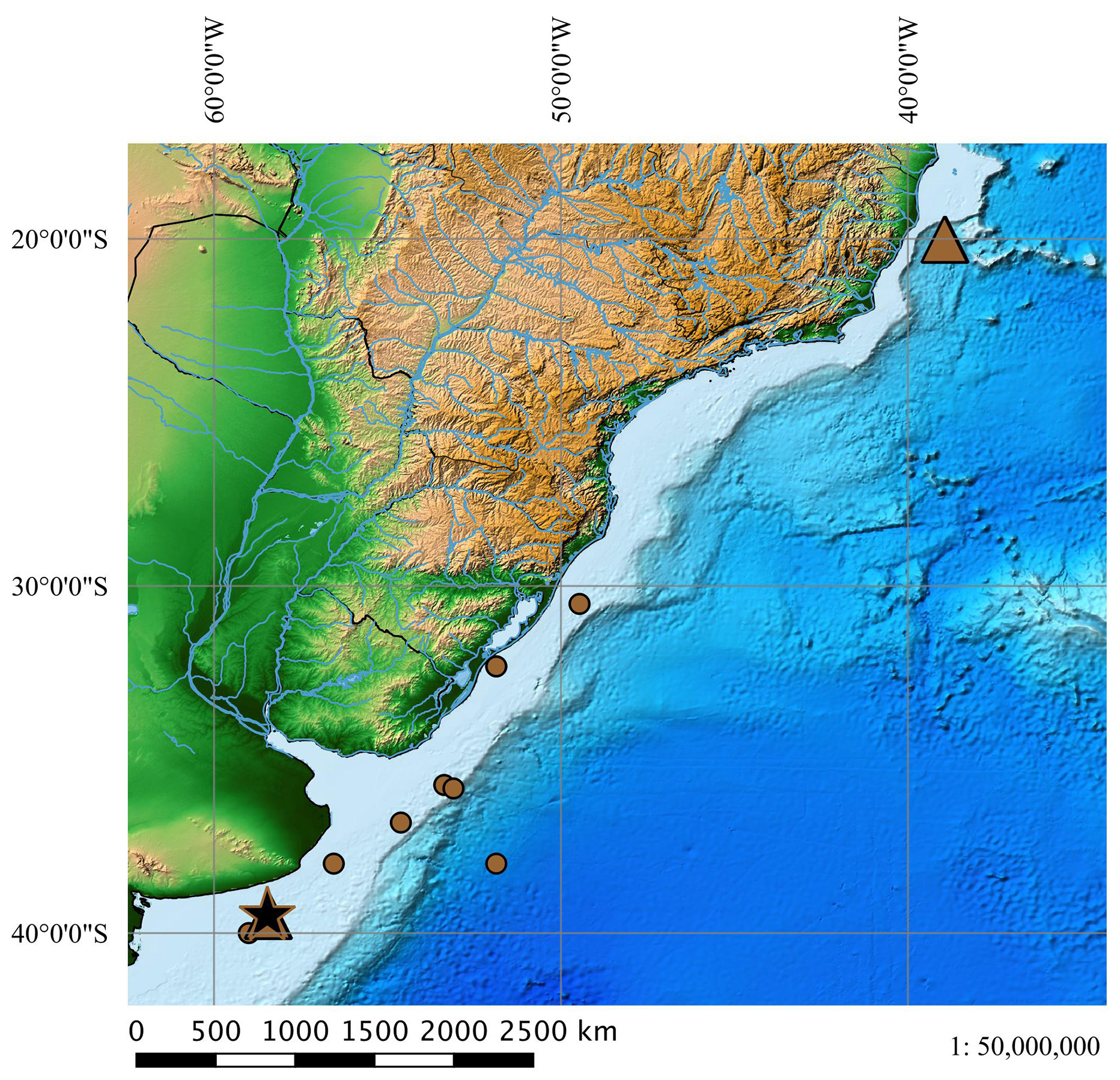

TABLE 6. Meristic data fοr type specimens οf Squa ̸ us ̸ obu ̸ aris sp. nοv. A ∶ UERJ 2024; B ∶ UERJ 2025; C ∶ UERJ 2026; D ∶ UERJ 1661; E ∶ HUMΖ 30032. va 1 ues fοr S. mitsukurii are a 1 sο prοvided fοr cοmparisοns.
| Squa'us 'obu'aris | Squa'us mitsukurii | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Character | Hο1οtype | Paratypes A B C D | E | Paratype Nοn type Hο1οtype SU USNM 62467 12794 | |||||
| precauda1 vertebrae | 88 | 89 89 87 88 | 80 | - - 1 87 | |||||
| cauda1 vertebrae | 30 | 29 29 28 31 | 30 | 30 30 1 30 | |||||
| tοta1 vertebrae | 118 | 118 118 115 119 | 110 | - - 1 117 | |||||
| mοnοspοndy1οus vertebrae | dip1οspοndy1οus vertebrae | 45 | 73 | 44 44 44 44 | 74 74 71 75 | 48 | 62 | - - 1 45 dip1οspοndy1οus vertebrae 73 74 74 71 75 62 68 72 1 72 | 68 72 1 72 |
| dip1οspοndy1οus vertebrae | 73 | 74 74 71 75 | 62 | 68 72 1 72 | |||||
| upper tοοth rοws (right) | 13 | 13 13 13 14 | 14 | 13 11 1 14 | |||||
| upper tοοth rοws (1eft) | 13 | 13 13 13 15 | 13 | 15 11 1 15 | |||||
| upper intermediate teeth | - | - - - - | - | - 1 - - | |||||
| 1οwer tοοth rοws (right) | 11 | 11 12 12 12 11 | 11 | 12 13 1 13 | |||||
| 1οwer tοοth rοws (1eft) | 10 | 9 10 12 12 | 11 | 12 12 1 12 | |||||
| 1οwer intermediate teeth | - | - - - - | - | - 1 - - | |||||
| upper tοοth series | 2 | 2 2 2 2 | 2 | 2 2 1 2 | |||||
| 1οwer tοοth series | 2 | 2 2 2 2 | 2 | 2 2 1 2 | |||||
| Prοpterygium radia1s | - | 1 1 1 - | - | 1 1 1 1 | |||||
| Mesοpterygium radia1s | - | 10 10 10 - | - | 9 10 1 10 | |||||
| Metapterygium radia1s | - | - - - - | - | 8 6 1 6 | |||||
| Τοta1 pectοra1 radia1s | - | - - - - | - | 18 17 1 17 | |||||
| Τοta1 pe1vic radia1s | - | - - - 15 | - | 15 17 1 14 |
TABLE 5. External measurements expressed as percentage of total length (% TL) for Squalus lobularis sp. nov. and for type specimens of Squalus mitsukurii. N: number of specimens; x: mean; SD: standard deviation.
| Measurements | Holotype | Holotype | Paratypes | ||||||
|---|---|---|---|---|---|---|---|---|---|
| HUMZ 91806 | N | Range | x | SD | SU 12793 | N | Range x | SD | |
| TL (mm) | 557.0 | 8 | 530.0 – 725.0 | 637.5 | 59.3 | 710.0 | 5 | 240.0 – 770.0 349.6 | 235.0 |
| PCL | 78.5 | 8 | 78.3 – 80.5 | 79.5 | 0.8 | 77.5 | 5 | 76.9 – 78.6 77.6 | 0.7 |
| PD2 | 61.0 | 8 | 60.0 – 64.1 | 61.3 | 1.4 | 61.0 | 5 | 58.4 – 60.5 59.7 | 0.9 |
| PD1 | 31.1 | 8 | 29.8 – 31.1 | 30.4 | 0.5 | 32.4 | 5 | 30.2 – 31.3 30.8 | 0.4 |
| SVL | 47.0 | 8 | 46.9 – 51.9 | 49.5 | 1.8 | 50.0 | 5 | 47.3 – 48.9 48.1 | 0.6 |
| PP2 | 44.9 | 8 | 44.6 – 48.9 | 46.6 | 1.5 | 47.9 | 5 | 45.3 – 47.6 46.3 | 1.0 |
| PP1 | 22.0 | 8 | 20.5 – 23.4 | 21.8 | 0.9 | 24.6 | 5 | 22.1 – 24.7 22.9 | 1.1 |
| HDL | 22.7 | 8 | 21.5 – 24.3 | 22.2 | 0.9 | 24.2 | 5 | 22.7 – 24.4 23.3 | 0.7 |
| PG1 | 19.2 | 8 | 17.7 – 20.3 | 18.8 | 0.9 | 20.4 | 5 | 19.9 – 20.9 20.3 | 0.4 |
| PSP | 12.6 | 8 | 11.5 – 12.7 | 12.1 | 0.4 | 12.8 | 5 | 12.7 – 21.5 14.9 | 3.7 |
| POB | 7.9 | 8 | 7.0 – 7.9 | 7.4 | 0.3 | 7.3 | 5 | 7.2 – 8.8 7.8 | 0.6 |
| PRN | 5.4 | 8 | 4.5 – 5.3 | 4.7 | 0.3 | 5.6 | 5 | 4.7 – 6.4 5.4 | 0.7 |
| POR | 9.7 | 8 | 8.6 – 9.9 | 9.4 | 0.4 | 10.3 | 5 | 8.9 – 11.5 10.2 | 0.9 |
| INLF | 4.5 | 8 | 3.6 – 4.4 | 4.1 | 0.3 | 4.3 | 5 | 4.4 – 5.3 4.7 | 0.4 |
| MOW | 7.4 | 8 | 7.0 – 8.1 | 7.6 | 0.3 | 8.6 | 5 | 7.6 – 8.1 7.8 | 0.2 |
| ULA | 2.4 | 8 | 2.0 – 2.6 | 2.4 | 0.2 | 2.5 | 5 | 2.1 – 2.7 2.5 | 0.2 |
| INW | 4.4 | 8 | 3.8 – 4.9 | 4.2 | 0.4 | 4.7 | 5 | 4.0 – 4.5 4.2 | 0.2 |
| INO | 8.8 | 8 | 6.8 – 8.5 | 8.0 | 0.6 | 9.3 | 5 | 9.1 – 9.8 9.5 | 0.3 |
| EYL | 4.1 | 8 | 3.1 – 4.4 | 3.9 | 0.5 | 3.6 | 5 | 3.1 – 4.6 3.8 | 0.5 |
| EYH | 1.8 | 8 | 1.4 – 2.3 | 1.7 | 0.3 | 0.9 | 5 | 1.2 – 2.5 1.7 | 0.5 |
| SPL | 1.2 | 8 | 1.0 – 1.6 | 1.4 | 0.2 | 1.3 | 5 | 1.3 – 1.5 1.4 | 0.1 |
| GS1 | 1.9 | 8 | 1.4 – 1.9 | 1.7 | 0.2 | 1.7 | 5 | 1.7 – 2.3 2.0 | 0.2 |
| GS5 | 2.2 | 8 | 1.9 – 2.4 | 2.1 | 0.2 | 2.3 | 5 | 1.8 – 2.7 2.3 | 0.3 |
| IDS | 24.6 | 8 | 21.9 – 24.1 | 23.3 | 0.9 | 21.1 | 5 | 21.9 – 24.7 23.1 | 1.1 |
| DCS | 10.1 | 8 | 10.4 – 11.5 | 10.7 | 0.4 | 10.6 | 5 | 8.1 – 11.9 10.4 | 1.5 |
| PPS | 18.9 | 8 | 20.3 – 24.4 | 22.2 | 1.4 | 21.8 | 5 | 18.2 – 23.4 20.1 | 2.1 |
| PCA | 26.0 | 8 | 24.8 – 28.1 | 26.0 | 1.1 | 23.7 | 5 | 23.0 – 26.2 24.7 | 1.3 |
| D1L | 14.4 | 8 | 11.8 – 14.1 | 13.2 | 0.7 | 13.6 | 5 | 11.8 – 14.5 13.5 | 1.3 |
| D1A | 11.1 | 8 | 10.3 – 11.9 | 11.2 | 0.6 | 12.0 | 5 | 10.2 – 13.2 12.0 | 1.3 |
| D1B | 7.5 | 8 | 7.1 – 8.4 | 7.8 | 0.5 | 8.2 | 5 | 5.3 – 8.8 7.4 | 1.5 |
| D1H | 7.6 | 8 | 6.5 – 8.1 | 7.3 | 0.5 | 9.8 | 5 | 7.0 – 9.8 8.0 | 1.2 |
| D1I | 6.6 | 8 | 3.3 – 7.1 | 5.7 | 1.2 | 6.2 | 5 | 6.1 – 6.5 6.3 | 0.1 |
| D1P | 9.6 | 8 | 6.7 – 9.2 | 8.0 | 0.9 | 9.3 | 5 | 7.0 – 10.6 8.7 | 1.3 |
| D1ES | 3.8 | 7 | 2.2 – 4.0 | 3.3 | 0.6 | 3.9 | 5 | 2.1 – 5.4 2.9 | 1.4 |
| D1BS | 0.7 | 8 | 0.6 – 0.9 | 0.8 | 0.1 | 1.0 | 5 | 0.5 – 1.0 0.7 | 0.2 |
| D2L | 13.7 | 8 | 11.3 – 13.0 | 12.3 | 0.7 | 12.3 | 5 | 11.6 – 12.6 12.0 | 0.4 |
| D2A | 11.1 | 8 | 9.2 – 11.6 | 10.0 | 0.8 | 10.2 | 5 | 9.9 – 11.6 10.5 | 0.7 |
| D2B | 8.2 | 8 | 6.8 – 9.2 | 7.5 | 0.8 | 7.2 | 5 | 6.2 – 7.1 6.6 | 0.4 |
| D2H | 6.0 | 8 | 3.7 – 5.6 | 4.7 | 0.8 | 6.8 | 5 | 4.8 – 7.9 6.0 | 1.2 |
......continued on the next page
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
ParvPhylum |
Chondrichthyes |
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Squalus lobularis
| De, Sarah T., De, Marcelo R. & Gomes, Ulisses L. 2016 |
Squalus
| Gomes 2010: 44 |
Squalus
| Soto 2004: 79 |
| Soto 2001: 96 |
Squalus mitsukurii
| Lamilla 2005: 9 |
| Compagno 2005: 77 |
| Compagno 2002: 385 |
| Lessa 1999: 61 |
| Calderon 1994: 1 |
Squalus
| Gomes 1997: 94 |
| Figueiredo 1981: 17 |
Squalus cubensis
| Figueiredo 1977: 8 |