
Chorocaris variabilis, Komai & Tsuchida, 2015
|
publication ID |
https://doi.org/ 10.1080/00222933.2015.1006702 |
|
publication LSID |
lsid:zoobank.org:pub:4BFC8C31-C006-4738-BED7-B13D3296A7CA |
|
DOI |
https://doi.org/10.5281/zenodo.4329042 |
|
persistent identifier |
https://treatment.plazi.org/id/287587FD-481B-FFDB-6C9B-FAD586A5FD4F |
|
treatment provided by |
Carolina |
|
scientific name |
Chorocaris variabilis |
| status |
sp. nov. |
Chorocaris variabilis View in CoL sp. nov.
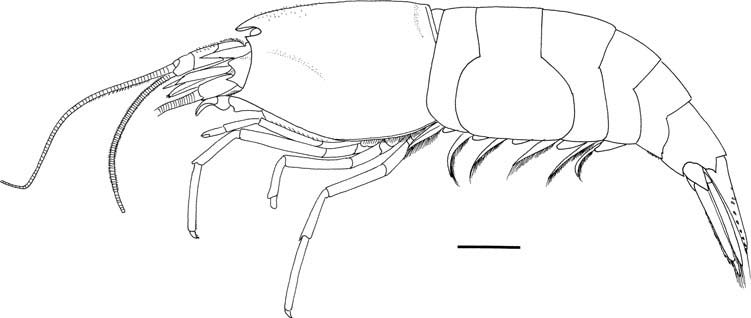
( Figures 6–10 View Figure 6 View Figure 7 View Figure 8 View Figure 9 )
Chorocaris View in CoL sp. – Zelnio and Hourdez 2009: 52.
Material examined
Holotype. Wave Mercury 2007 (Luk Luk) Campaign, dive 28, South Su , Manus Basin , Bismarck Sea , 03°08.09 ʹ S, 152°10.5 ʹ E, 1310 m, 10 April 2007, female (cl 13.6 mm), CBM-ZC 11,945. GoogleMaps
Paratypes
Manus Basin. Wave Mercury 2007 (Luk Luk) Campaign, same data as holotype, 10 males (cl 5.2–9.1 mm), CBM-ZC 11946; same data as holotype, 14 females (cl 6.5–12.9 mm), 6 ovigerous females (cl 10.7–13.5 mm), CBM-ZC 11947; dive 30, South Su , 3°08.09 ʹ S, 152°10.5 ʹ E, 1305 m, 11 April 2007, 9 males (cl 10.6– 11.9 mm), CBM-ZC 11948; same data, 6 females (cl 13.3–15.0 mm), 1 ovigerous female (cl 12.8 mm), CBM-ZC 11949; dive 34, same site, 1310 m, 14 April 2007, 13 males (cl 6.6–9.0 mm), 9 females (cl 5.0–12.0 mm), 1 ovigerous female (cl 9.6 mm), CBM-ZC 11950; dive 53, Solwara 1, 03°07.91 ʹ N, 152°09.1 ʹ E, 1570 m, 25 April 2007, 27 males (cl 5.1–7.5 mm), CBM-ZC 11951; same data, 26 females (cl 4.4–10.4 mm), 14 ovigerous females (cl 6.7–9.4 mm), CBM-ZC 11952; same data, 22 males (cl 4.8–7.2 mm), 14 females (cl 5.2–12.0 mm), 3 ovigerous females (cl 9.6–14.0 mm), OUMNH. ZC GoogleMaps .2014–01-015. RV Yokosuka, YK 06-13 cruise, DS Shinkai 6500, dive #978, PACMANUS site, 03°43.60 ʹ S, 151°40.32 ʹ E, 1683 m, 16 September 2006, 1 male (cl 8.2 mm), 1 female (cl 7.6 mm), 1 ovigerous female (cl 10.8 mm), JAMSTEC 066720–066722; DS Shinkai 6500, dive #981, Vienna Woods , 03°09.813 ʹ S, 151°16.737 ʹ E, 2480 m, 14 September 2006, 1 ovigerous female (cl 10.3 mm), JAMSTEC 068873; DS Shinkai 6500 dive #982, PACMANUS site, 03°43.608 ʹ S, 151°40.328 ʹ E, 1684 m, 20 September 2006, 3 specimens (not measured; DNA extracted, preserved in frozen condition), JAMSTEC 070103–070105. North Fiji Basin. RV GoogleMaps Yokosuka, YK 06-13 cruise, DS Shinkai 6500, dive #985, Mussel Hill site, 16°54.444 ʹ S, 173°54.895 ʹ E, 1988 m, 2 October 2006, 8 males (cl 4.9–7.1 mm), 2 females (cl 5.4, 7.0 mm), JAMSTEC 070543–070552; same data, 1 specimen (not measured; DNA extracted, preserved in 99% ethanol), JAMSTEC 070543. Vanuatu. RV GoogleMaps Sonne, SO 229-060-N201, Nifonea vent field, 18°07.735 ʹ S, 169°31.112 ʹ E, 1873 m, 14 July 2013, 9 males (cl 6.6–9.3 mm), 3 females (cl 6.7–7.7 mm), 1 ovigerous female (cl 7.6 mm), CBM- ZC 12 , 481 GoogleMaps .
Non-type. Vanuatu: same data as CBM-ZC 12481, 2 juveniles (cl 3.8, 3.9 mm), CBM- ZC 12 , 482 .
Description
Females. Body integument smooth, shiny, fairly thin, with scattered short setae, more numerous on dorsal side.
Rostrum ( Figure 7A–C View Figure 7 ) falling slightly short of midlength of first segment of antennular peduncle, triangular in dorsal view; tip varying from blunt (ovigerous females) to acute or subacute (non-ovigerous females); dorsal surface smooth. Carapace ( Figures 6 View Figure 6 , 7A–C View Figure 7 , 10G–J) compressed laterally; dorsal surface sloping down anteriorly to rostrum; antennal tooth triangular, acute (non-spawning females) or acute to blunt (spawning females); pterygostomial angle strongly produced anteriorly in acute or subacute tooth extending as far as dorsolateral process of antennal basicerite.
Third pleonal pleuron ( Figure 6 View Figure 6 ) broadly rounded, always unarmed; fourth pleuron rounded or bluntly angular posteroventrally, without sharp tooth ( Figures 7D View Figure 7 , 10K); fifth pleuron usually with small posteroventral tooth and with zero to four tiny denticles on sinuous to convex posterolateral margin ( Figures 7D View Figure 7 , 10K). Sixth pleomere ( Figure 7D View Figure 7 ) about 1.2 times longer than fifth pleomere and about 1.1 times longer than high, having acute or subacute posteroventral tooth, posterolateral process terminating in sharp tooth. Telson ( Figure 7E View Figure 7 ) about 2.2 times longer than anterior width, falling short of posterior margins of uropods, very slightly narrowed posteriorly, normally armed with six to eight dorsolateral spines arranged in sinuous row; posterior margin convex, occasionally with minute median denticle or tooth and with two pairs of spines at lateral angles and 20–24 long plumose setae ( Figure 7F View Figure 7 ).
Eyes ( Figure 7A, B View Figure 7 ) lacking setae on anterior surface. Antennular peduncle ( Figure 7A, B View Figure 7 ) stout, reaching distal margin of antennal scale; first segment with moderately large distolateral and small distomesial teeth and prominent, blunt proximolateral tubercle; stylocerite slender, nearly straight, falling slightly short of distal margin of second segment; second segment 1.0–1.1 times as long as wide, with distomesial tooth subequal in size to corresponding tooth on first segment; flagella moderately slender. Antennal peduncle ( Figure 7A, B, G View Figure 7 ) stout; basicerite with ventrodistal tooth overreaching dorsodistal projection; fifth segment (= carpocerite) slightly overreaching midlength of antennal scale; antennal scale suboval, about 0.4 times as long as carapace, 1.6 times longer than wide, lateral margin almost straight, but convex near distolateral tooth, distolateral tooth acute or subacute, clearly separated or approximated to lamella.
Mouthparts generally similar to those of C. parva sp. nov., as figured ( Figures 8A–E View Figure 8 , 9A View Figure 9 ). Second maxilliped ( Figure 8E View Figure 8 ) relatively stout. Epipod of third maxilliped nearly trapezoidal, slightly unequally bilobed ( Figure 9B View Figure 9 ).
First pereopod ( Figure 9C–E View Figure 9 ) reaching or slightly overreaching midlength of antennal scale, moderately slender to robust, polymorphic as in C. vandoverae and C. paulexa . Second to fifth pereopods relatively stout. Second pereopod ( Figure 9G View Figure 9 ) reaching as far as first pereopod; chela 0.6–0.8 times as long as carpus; dactylus 1.1–1.2 times longer than palm ( Figure 9H View Figure 9 ). Third pereopod ( Figure 9I View Figure 9 ) overreaching antennal scale by half to full length of propodus; dactylus somewhat compressed laterally, 0.15–0.20 times as long as propodus, terminating in strong, curved unguis, flexor surface with 10–16 accessory spinules arranged in three or four irregular rows ( Figure 9J View Figure 9 ); propodus with spinules arranged in two separate rows or double row on flexor surface ( Figure 9J View Figure 9 ); carpus 075–0.90 times as long as propodus; merus unarmed; ischium usually unarmed, but rarely with one tiny spine. Fourth pereopod ( Figure 9K View Figure 9 ) similar to third pereopod in structure; ischium always unarmed. Fifth pereopod ( Figure 9L View Figure 9 ) generally similar to third and fourth; propodus with both lateral and mesial rows of spinules, each arranged into double row (spinules more numerous in lateral row than mesial row) ( Figure 9M View Figure 9 ); propodus and carpus combined distinctly longer than merus and ischium combined; carpus 0.6–0.7 times as long as propodus; ischium always unarmed.
Appendices internae on second to fourth pereopods simple, on fifth pereopod normally developed, bearing terminal cluster of coupling hooks. Uropod ( Figure 7E View Figure 7 ) with protopod ( Figure 7H View Figure 7 ) bearing subacute to acute posterolateral process; rami both overreaching posterior margin of telson; exopod slightly longer than endopod, with subequal posterolateral spines.
Males. Body less robust than in females. Rostrum ( Figure 10A, B) acutely pointed, occasionally faintly carinate on dorsal midline. Carapace with sharp antennal tooth ( Figure 10B); pterygostomial process narrower than in females, sharply pointed ( Figure 10B). Pleon with fourth pleuron armed with small acute posteroventral tooth; fifth pleuron also with sharp posteroventral tooth and additional one to three small teeth or denticles on posterolateral margin. Antennal scale 1.8–1.9 times longer than wide; distolateral tooth more slender than in females, always clearly separated from lamella, acute ( Figure 10A). Pereopods more slender than in females; ischium of third pereopod unarmed or armed with one spine, of fourth pereopod usually unarmed, but rarely armed with one spine ( Figure 10C, D), and of fifth pereopod always unarmed. Endopod of first pleopod ( Figure 10E) bilobed distally, mesial lobe prominent, with three or four long spiniform setae mesially, lateral lobe rather obsolete, also with row of spiniform setae extending onto lateral margin, proximal four or five of these setae bearing setules; mesial margin with row of spiniform setae. Appendix masculina of second pleopod ( Figure 10F) stout, slightly longer than appendix interna, armed with about 10 spiniform setae distally. Posterolateral process of uropodal protopod acuminate.
Juveniles. Generally similar to adults, but body more compressed laterally. Rostrum rounded distally. Eyes each with minute tubercle on anterior surface dorsomesially.
Size
Males cl 4.8–11.9 mm; females cl 4.4–15.0 mm, ovigerous females cl 6.7–13.5 mm.
Coloration in life
Not known.
Distribution
Hydrothermal vents in southwestern Pacific. Manus Basin: South Su, 1305–1310 m; Solwara 1, 1570 m; PACMANUS, 1683–1684 m; Vienna Woods, 2480 m. North Fiji Basin: Mussel Hill, 1988 m. Vanuatu: Nifonea vent field, 1873 m.
Remarks
The material of this new species came from three hydrothermal vent sites in the southwestern Pacific, i.e. Manus Basin, North Fiji Basin and Vanuatu. The specimens from the three locations are morphologically very similar. Of the four specimens used in the molecular phylogenetic analysis (see below), three specimens came from the Manus Basin, and one specimen was from the North Fiji Basin. The maximum genetic divergence among the four specimens is 1.8% ( Table 1). Consequently it is reasonable to consider that these specimens represent a single species.
Chorocaris variabilis sp. nov. exhibits a substantial degree of variation in the shape of the rostrum and of the carapace teeth (see above). Males and non-spawning females differ from spawning females in having an acuminate rostrum with an occasionally faintly carinate dorsal surface (males), a sharply pointed antennal tooth and branchiostegal tooth on the carapace, and a clearly separated distolateral tooth on the antennal scale. Spawning females are characterized by the bluntly pointed rostrum, and antennal and branchiostegal teeth on the carapace.
Chorocaris variabilis sp. nov. is morphologically very similar to C. paulexa and C. vandoverae particularly in the prominent pterygostomial tooth on the carapace and the possession of 10–16 accessory spinules arranged in three or four rows on the flexor faces of the dactyli of the third to fifth pereopods. Nevertheless, C. variabilis differs from these two species in the possession of a conspicuous posteroventral tooth and additional denticles on the fifth pleonal pleuron and in the absence of setae on the anterior face of each eye. In C. paulexa and C. vandoverae , the posteroventral angle of the fifth pleonal pleuron is rounded or at most angular and the eye has several stiff setae on the anterior face. Furthermore, although somewhat variable in C. variabilis , the ventral surface of the rostrum is usually less convex than in C. paulexa and C. vandoverae .
Chorocaris vandoverae does not exhibit strong variation between males and nonspawning females on the one hand, and spawning females ( Komai and Segonzac 2008). Hence, males and non-spawning females of this new species are readily distinguished from those of C. vandoverae by the acuminate rostrum and sharply pointed antennal and pterygostomial teeth on the carapace. In C. vandoverae , the rostrum is rounded distally, the pterygostomial tooth is more bluntly pointed, and the posterolateral angle of the fifth pleonal pleuron is at most subacute and devoid of additional denticles on the posterolateral margin. Furthermore, the armature of the appendix masculina of the second pleopod may be different between C. variabilis and C. vandoverae . In the new species, spiniform setae are restricted to the distal half of the appendix masculina, while similar spiniform setae form a row extending proximally beyond the midlength of the appendix masculina in C. vandoverae . With regard to spawning females, C. variabilis is distinguished from C. vandoverae in possessing a sharp posteroventral tooth on the fifth pleonal pleuron and a more sharply pointed protopod of the uropod.
No information on the male is available for C. paulexa (cf. Martin and Shank 2005). Therefore variability between males and females in this species remains unknown. Nevertheless, the rostrum is proportionally narrower in C. variabilis than in C. paulexa (see Martin and Shank 2005; Komai and Segonzac 2008). In addition, the third to fifth pereopods seem to be more slender in this new species than in C. paulexa , but the significance of this character may be questionable because of intraspecific variation (as seen in other confamilial or congeneric species; see Komai and Segonzac 2003, 2004, 2008).
Records of C. vandoverae from hydrothermal vents on the North Fiji Basin ( Desbruyères et al. 1994) and the Lau Basin ( Desbruyères et al. 1994; Zelnio and Hourdez 2009) are referred to C. variabilis on the basis of the close morphological similarity between C. vandoverae and this new species, and their geographical distribution. Chorocaris sp. 2 reported from the Manus Basin by Zelnio et al. (2010) corresponds to C. variabilis . Consequently the occurrence of C. vandoverae outside the Mariana Back Arc Basin is unconfirmed.
Etymology. From the Latin variabilis (= variable) in reference to the substantial morphological variation seen in this new species.



| ZC |
Zoological Collection, University of Vienna |
| RV |
Collection of Leptospira Strains |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Chorocaris variabilis
| Komai, Tomoyuki & Tsuchida, Shinji 2015 |
Chorocaris
| Zelnio K & Hourdez S 2009: 52 |