
Coelostoma Brullé, 1835
|
publication ID |
https://doi.org/10.11646/zootaxa.3887.3.4 |
|
publication LSID |
lsid:zoobank.org:pub:ABD688ED-D7F6-40F1-8821-0C339293A2A5 |
|
DOI |
https://doi.org/10.5281/zenodo.6140230 |
|
persistent identifier |
https://treatment.plazi.org/id/297C0906-F21B-FFB2-0CBA-820490EAE117 |
|
treatment provided by |
Plazi |
|
scientific name |
Coelostoma Brullé, 1835 |
| status |
|
Coelostoma Brullé, 1835 View in CoL
( Figs. 1–32 View FIGURES 1 – 8 View FIGURES 9 – 14 View FIGURES 15 – 23 View FIGURES 24 – 31 View FIGURES 32 – 33 )
Cercydium Klug, 1833: 160 (partim) (nomen nudum).
Coelostoma Brullé, 1835: 293 View in CoL . Type species: Hydrophilus orbicularis Fabricius, 1775 (by monotypy). = Cyclonotum Erichson, 1837: 212 View in CoL . Type species: Hydrophilus orbicularis Fabricius, 1775 (by monotypy; junior objective synonym).
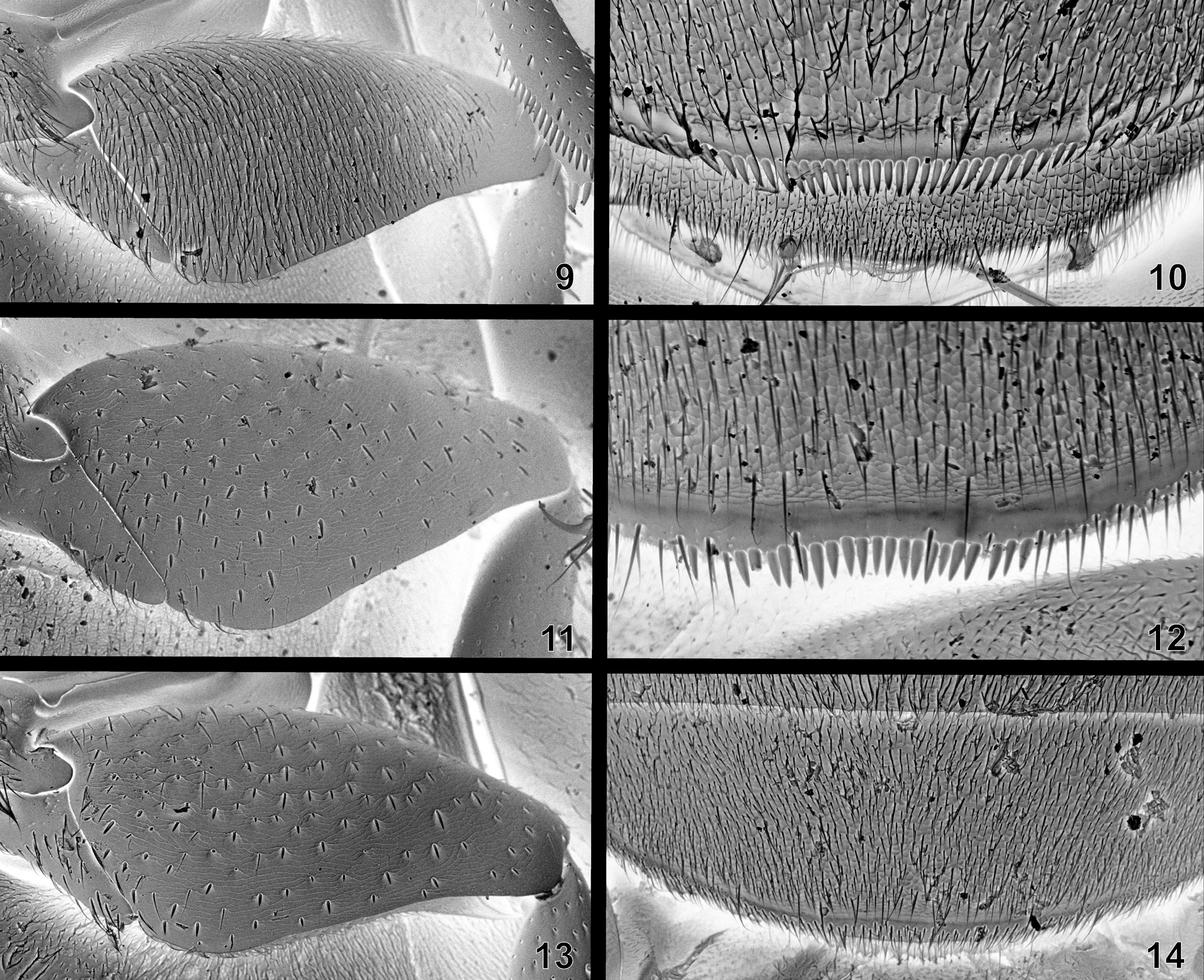
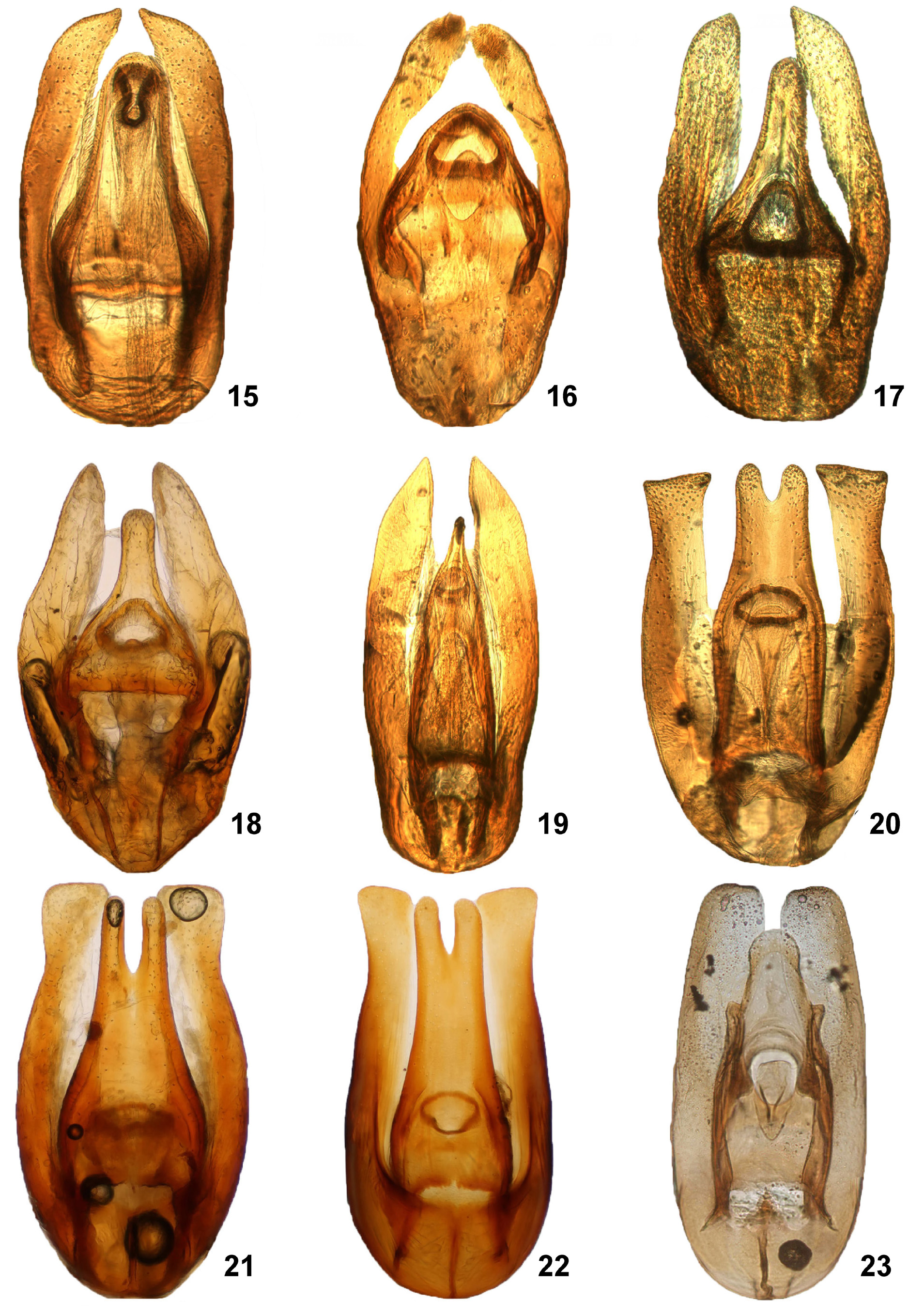
Diagnosis. Body broadly oval, more or less uniformly brown to black ( Figs. 1–2 View FIGURES 1 – 8 ); clypeus covering bases of antennae; antenna with 9 antennomeres, club loosely segmented; prosternum more or less bulging medially, often with medially dentiform anterior margin; mesoventrite at least partly fused to mesepisterna, strongly raised posteriorly to form an arrowhead-shaped process ( Fig. 6 View FIGURES 1 – 8 ); anteromedian pit-like groove of mesoventrite present ( Fig. 6 View FIGURES 1 – 8 ); metaventrite with raised middle portion; metaventral process strongly projecting anteriorly between mesocoxae, abutting mesoventral elevation ( Fig. 6 View FIGURES 1 – 8 ); meso- and metatarsus with first tarsomere clearly longer than second tarsomere; elytra with sharply impressed sutural stria in posterior half ( Fig. 1 View FIGURES 1 – 8 ); elytra without striae or serial punctures, sometimes with traces of serially arranged punctures laterally; first abdominal ventrite not carinate mesally, sometimes except the extreme base; apical part of abdominal ventrite 5 entire or emarginate ( Figs. 10, 12, 14 View FIGURES 9 – 14 ); phallobase of aedeagus extremely reduced ( Figs. 15–33 View FIGURES 15 – 23 View FIGURES 24 – 31 View FIGURES 32 – 33 ).
Differential diagnosis. The majority of species of Coelostoma may be easily distinguished from other genera of the Coelostomatini by the combination of uniformly blackish dorsal coloration, loosely segmented antennal club, absence of clear elytral series, presence of sutural stria, and abdominal ventrite 1 carinate at most at extreme base. These characters allow the safe identification of all Chinese species treated below. Coelostoma gentilii sp. nov. has weak traces of serially arranged punctures at lateralmost parts of each elytron, but it is otherwise extremely similar to all other species and easy-to-recognize as a member of Coelostoma .
Comments. When a wider spectrum of Asian coelostomatines is examined, it is evident that several species treated contemporarily as “atypical” representatives of the genus Dactylosternum Wollaston, 1854 (e.g., D. arabicum Balfour-Browne, 1951 , D. coelostomoides Orchymont, 1923 and D. indicum Orchymont, 1923 ) may in fact belong to Coelostoma . These species do not match the current diagnosis of Coelostoma in presence of elytral series or the carinate first abdominal ventrite, which may indicate that these two characters are not good for delimiting coelostomatine genera. Additional studies of mentioned species is necessary to understand their generic assignment and the definition of Coelostoma as a genus.
Species-level identification. Most species of Coelostoma are extremely similar to each other externally, and the external characters (especially the pubescence of mesofemora and presence/absence of stout setae on abdominal apex) are mostly useful only to distinguish the subgenera. In few cases, dorsal coloration and character of dorsal punctuation of pronotum and elytra may allow to identify the specimens to species, but in most cases the morphology of male genitalia is the only character which allows reliable species identification.
The morphology of the aedeagus is very variable within the genus, and provides easy-to-observe characters for identification of species. Important characters include: (1) the general form of the aedeagus, (2) the form of the paramere and its apical portion, (3) the form of the median lobe and its length compared to the parameres, (4) the position and shape of the gonopore, and (5) the presence and shape of the internal median sclerite projecting towards gonopore. We recommend examination of the aedeagus in a “wet” state (i.e. in drop of water or glycerin or mounted in hydantoin, euparal or other mounting medium) and in dorsal view. In dry-mounted and/or ventrally observed genitalia, several critical characters are not seen or are deformed, which may lead to incorrect identification.
Biology. All species of Coelostoma for which biology is known are aquatic, collected either directly among submerged plants at the edge of water, or from wet places along streams, rivers and standing water. Many Chinese species are night-active: they hide themselves outside of water during the day (usually under moss or roots of plants growing next to the watercourse, Fig. 4 View FIGURES 1 – 8 ) and may be found feeding on wet and submerged surfaces including of wet rocks and artificial concrete surfaces at night ( Fig. 5 View FIGURES 1 – 8 ). Some species may be collected from mud or from under wet leaf litter, few species are only found in interstitial habitats under stones and among gravel at sides of stony rivers. Some species may be attracted at light, mostly just after the sunset (i.e. circa between 7 and 9 pm during summer in southern China, very rarely they are collected after 10 pm).



No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Sphaeridiinae |
Coelostoma Brullé, 1835
| Jia, Fenglong, Aston, Paul & Fikáček, Martin 2014 |
Coelostoma Brullé, 1835 : 293
| Erichson 1837: 212 |
| Brulle 1835: 293 |
Cercydium
| Klug 1833: 160 |