
Cyrtodactylus nyinyikyawi, Grismer & Wood & Thura & Win & Quah, 2019
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4577.2.3 |
|
publication LSID |
lsid:zoobank.org:pub:5955B151-353F-4FB2-8EEC-A5674F8F0139 |
|
DOI |
https://doi.org/10.5281/zenodo.4324115 |
|
persistent identifier |
https://treatment.plazi.org/id/2A7FCD3A-FFE1-380F-FF0A-86DE3EC5FF12 |
|
treatment provided by |
Felipe |
|
scientific name |
Cyrtodactylus nyinyikyawi |
| status |
sp. nov. |
Cyrtodactylus nyinyikyawi sp. nov.
Shwe Settaw Bent-toed Gecko
( Fig. 4 View FIGURE 4 )
Holotype. Adult female CAS 226139 View Materials collected on 14 September 2002 at 1030 hrs by Thin Thin , Kyi Soe Lwin , and Hla Tun from Shwe Settaw Wildlife Sanctuary, Min Bu Township, Magway Region, Myanmar (20.05972°N, 94.59611°E; 137 m in elevation). GoogleMaps
Diagnosis. Cyrtodactylus nyinyikyawi sp. nov. differs from all other species in the peguensis group by having the unique combination of nine supralabials; eight infralabials; 35 paravertebral tubercles; 20 longitudinal rows of body tubercles; 35 ventral scales; 19 subdigital lamellae on the fourth toe; four rows of enlarged, post-precloacal scales; keeled, conical, body tubercles; top of head bearing dark blotches; five dark, body bands; dark body bands lacking paravertebral elements; and maximum SVL of 64.5 mm ( Table 3).
Description of holotype. Adult female, SVL 64.5 mm; head moderate in length (HL/SVL 0.25), wide (HW/ HL 0.62), somewhat flattened (HD/HL 0.42), distinct from neck, triangular in dorsal profile; lores inflated, prefrontal region concave, canthus rostralis rounded; snout elongate (ES/HL 0.40), rounded in dorsal profile; eye large (ED/HL 0.20); ear opening elliptical, moderate in size (EL/HL 0.09); eye to ear distance greater than diameter of eye; rostral rectangular, partially divided dorsally, bordered posteriorly by large left and right supranasals separated small internasal, laterally by first supralabials; external nares bordered anteriorly by rostral, dorsally by large supranasal, posteriorly by three postnasals (upper largest), ventrally by first supralabial; nine (R) supralabials extending to below midpoint of eye ball; eight rectangular infralabials tapering smoothly to below posterior margin of eye ball; scales of rostrum and lores flat, larger than granular scales on top of head and occiput; scales on top of head and occiput intermixed with slightly enlarged tubercles; dorsal supraciliaries not elongate or keeled; mental triangular, bordered laterally by first infralabials and posteriorly by large, left and right trapezoidal postmentals that contact medially for 70% of their length posterior to mental; one row of slightly enlarged chinshields tapering posteriorly to fourth infralabial; and gular and throat scales small, granular, grading posteriorly into larger, flatter, smooth, subimbricate to imbricate, pectoral and ventral scales.
Body relatively short (AG/SVL 0.52) with weak ventrolateral folds; dorsal scales small, interspersed with larger, semi-regularly arranged, weakly keeled tubercles; tubercles extend from occiput onto base of tail but no farther; tubercles on occiput and nape smaller than those on posterior portion of body; approximately 20 longitudinal rows of dorsal tubercles; approximately 35 paravertebral tubercles; 35 flat, imbricate, ventral scales larger than dorsal scales; seven dimpled, precloacal scales; and four rows of enlarged post-precloacal scales.
Forelimbs moderate in stature, relatively short (FL/SVL 0.13); flat scales of anterior margin of forearm larger than those on body, not interspersed with tubercles; palmar scales raised; digits relatively short, well-developed, inflected at basal, interphalangeal joints, slightly narrower distal to inflections; claws well-developed, sheathed by a dorsal and ventral scale; hind limbs more robust than forelimbs, moderate in length (TBL/SVL 0.17), covered dorsally by granular scales interspersed with slightly larger, weakly keeled tubercles and anteriorly by large, flat, imbricate scales; ventral scales of femora flat, imbricate, larger than dorsals, lacking a distinct row of enlarged femoral scales; small postfemoral scales form an abrupt union with larger, flat ventral scales of posteroventral margin of thigh; subtibial scales flat, imbricate; plantar scales slightly raised; digits relatively short, welldeveloped, inflected at basal, interphalangeal joints, slightly narrower distal to inflections; 19 subdigital lamellae (R,L) on fourth toe; claws well-developed, base of claw sheathed by a dorsal and ventral scale; two enlarged postcloacal tubercles at base of tail; postcloacal scales flat.
Proximal 14.4 mm of tail original, posterior 32.4 mm regenerated, 5.4 mm in width at base, tapering to a point; dorsal scales of original of tail granular rapidly becoming flatter posteriorly; dorsal scales of regenerated tail large, flat, semi-regular in arrangement; and transversely enlarged, median, subcaudal scales twice as wide as long, not extending onto lateral margin of tail in original section.
Coloration in life ( Fig. 4 View FIGURE 4 ). Dorsal ground color of head body, limbs, and tail yellow; top of head bearing large, dark-brown, irregularly shaped blotches edged in yellow; dark-brown, wide, nuchal loop extending from posterior margin of one eye, across occiput, to posterior margin of other eye; nape bearing a wide, dark-brown band edged in yellow; four wide, dark-brown body bands between limb insertions edged in yellow lacking distinct, paravertebral components; large, round, dark-brown markings between body bands two and three and three and four; seven or eight smaller, somewhat diffuse brown blotches along lower margins of flanks; one wide, dark-brown post-sacral band edged in yellow not bearing paravertebral sections; one dark-brown caudal band on original portion of tail; regenerated portion of tail light-colored bearing diffuse, randomly arranged, dark markings; dorsal portion of forelimbs darkly mottled to banded; dorsal portion of hind limbs bearing irregularly shaped, dark-brown blotches edged in yellow. All ventral surfaces generally beige, immaculate.
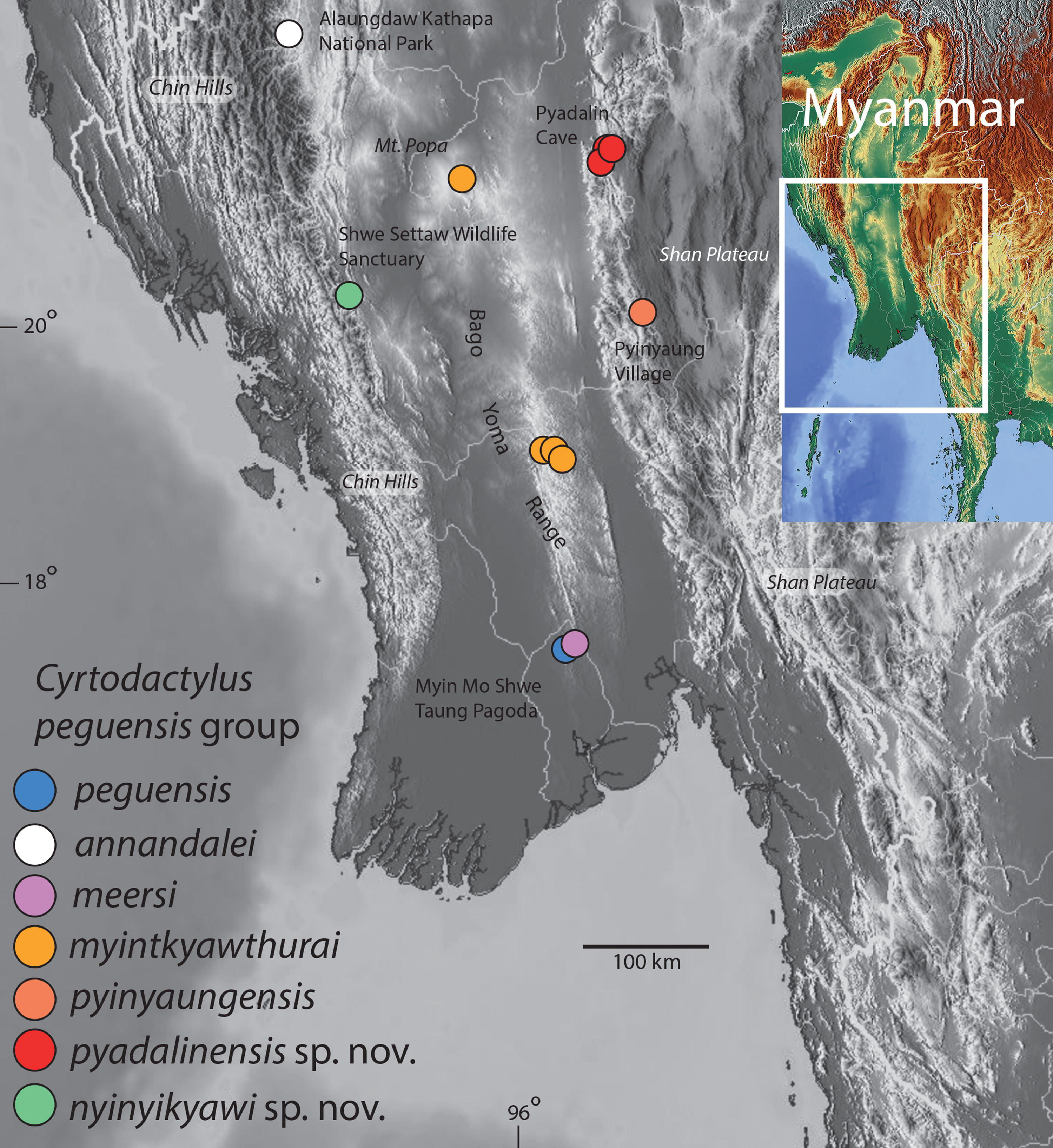
Distribution. Cyrtodactylus nyinyikyawi sp. nov. is known only from the type locality of the Shwe Settaw Wildlife Sanctuary, Min Bu Township, Magway Region, Myanmar ( Fig. 1 View FIGURE 1 ).
Etymology. The specific epithet, nyinyikyawi is a patronym honoring Nyi Nyi Kyaw the Director General of the Forestry Department for his contributions to conservation efforts in Myanmar in general and to our work in particular.
Natural History. The holotype CAS 226139 View Materials is a gravid female collected on the ground in secondary dry deciduous hardwood forest at 1030 hrs along the edges of a small seasonal lake. Being gravid with two eggs indicates that the monsoon month of September falls within this species’ reproductive season .
Comparisons Cyrtodactylus nyinyikyawi sp. nov. is the sister species of a clade that includes the sister species C. peguensis and C. pyinaungensis and C. myinykyawthurai ( Fig. 2 View FIGURE 2 ). It differs from C. peguensis by a 9.3% uncorrected pairwise sequence divergence, from C. myintkyawthurai by an 11.3–12.3% sequence divergence, and from C. pyinaungensis by a sequence divergence of 10.0–10.3%. Outside this clade, C. nyinyikyawi sp. nov. differs from C. meersi by a sequence divergence of 10%, from the Panluang-Pyadalin Cave population if differs by 9.3– 9.7%, and from C. annandalei it differs by 10.3%. It differs from all species of the peguensis group by having a higher number of paravertebral tubercles (35 vs. 25–33, collectively) and differs from all other species except the Panluang-Pyadalin Cave population by the dark dorsal bands lacking distinct, paravertebral elements as opposed to having them ( Figs. 4 View FIGURE 4 , 5 View FIGURE 5 ). It differs further from C. annandalei in that the top of the head is blotched as opposed to being unicolor It differs further from the Panluang-Pyadalin Cave population in having 35 as opposed to 38–40 ventral scale rows. Other differences separating C. nyinyikyawi sp. nov. form other peguensis group species are listed in Tables 2 and 3.
continued. myintkyawthurai , C. pyinyaungensis , and C. pyadalinensis sp. nov.
Remarks. Some (i.e. Dayrat 2005; Thomas Hbrek, in litt, 2018) have grave concerns about descriptions of new species based on only a single specimen, and posit that this should ‘never’ be done because such a description cannot take into account intraspecific variation that could potentially preclude its specific recognition. The myopic nature of this opinion notwithstanding, it is not only incorrect philosophically—as the ontological existence of a species is independent of its diagnosis ( Frost & Kluge 1994)—it is counterproductive in reality. Additionally, such tactics would impede biodiversity studies in general and taxonomy in particular. Estimates have shown that 19% of all new vertebrate species described between 2000 and 2010 were based on a single specimen ( Lim et al. 2012) and that number is likely to have increased in the last seven years—an indication that often, this is the logical first step in constructing species delimitation hypotheses (i.e. integrative taxonomies). Furthermore, with well-supported phylogenetic data such as that herein indicating that the specimen in question is not nested within or a sister species to any other species and shares a 9.3–12.3% uncorrected pairwise sequence divergence from its closest relatives, renders any morphological arguments to the contrary moot—regardless of these arguments’ erroneous conflation and confusion of ontology and epistemology. However, in this case, Cyrtodactylus nyinyikyawi sp. nov. has widely differing morphological and color pattern characters that at this point, distinguish it from all other species in the peguensis group, thus further eclipsing assumptions that it may be conspecific with something else. We are concerned about describing a new species based on a single specimen but only because the diagnosis is incomplete, not because it has anything to do with the reality of the specimen representing a distinct, independently evolving lineage based on the molecular evidence. Given the general ongoing biodiversity crisis throughout Southeast Asia, we felt it prudent to describe this species for potential protective status rather than delay its publication for the sake of a better diagnosis.
The weak part of recognizing this specimen as a distinct species is not the incomplete diagnosis, but that the species was initially delimited on the basis of a single-locus mtDNA phylogeny. It is well-documented that mtDNA phylogenies can reveal significant structure in a data set by recovering sequentially nested monophyletic groups even though within that same data set, nuclear genes can indicate significant gene flow among these groups (e.g. Shaw 2002; Fisher-Reid & Wiens,2011; Toews & Brelsford 2012), thus precluding their species status. This weakens any hypothesis of specific identity based solely on mtDNA data. Nonetheless, given the current data available concerning its phylogenetic relationships and the discrete morphological and color pattern differences separating C. nyinyikyawi sp. nov. from its congeners in the peguensis group, we regard its specific identity as a robust, testable hypothesis.



No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |