
Dioscorea buckleyana Wilkin, 2009
|
publication ID |
https://doi.org/ 10.5252/a2009n2a2 |
|
persistent identifier |
https://treatment.plazi.org/id/2C42E241-FFDA-FF92-FF56-CD44FF2BD6D6 |
|
treatment provided by |
Carolina |
|
scientific name |
Dioscorea buckleyana Wilkin |
| status |
|
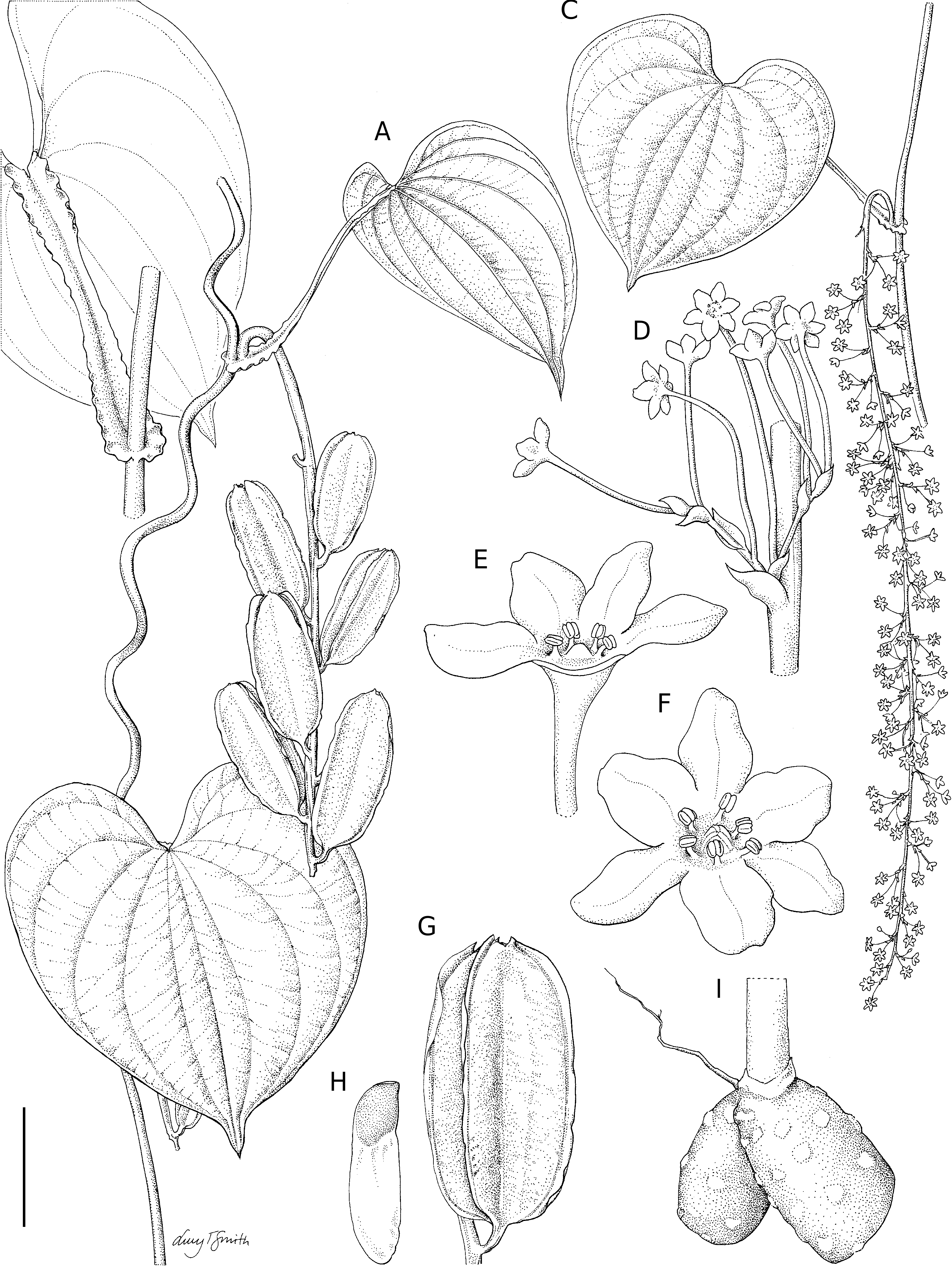
1. Dioscorea buckleyana Wilkin View in CoL , sp. nov. ( Fig. 1 View FIG )
Dioscorea pteropodae affinis sed petiolo 24-76 mm (nec 9-26 mm) longo, lamina folii chartacea vel crasse chartacea (nec coriacea ), pedicello primo cymularum in inflorescentia masculina ut minimum 4.8 mm longo (nec ut maximum 2.9 mm tantum longo), capsula ad apicem obtuse acuta vel truncata vel non profunde retusa (nec rotundata ) tristrata (nec bistrata) mesocarpio tenui medullosoque circumcirca axem proviso (nec mesocarpio carentia), exocarpio evoluto 1.5-3 mm (nec c. 1.5 mm tantum) ultra marginem incrassatam endocarpi extenso differt.
TYPUS. — Madagascar. Antsiranana (Diégo Suarez) Province, Antsiranana II District, Montagne des Français , 13.IV.1970, ♀ fr., Veyret 1301 (holo-, P! [P00496391]) .
PARATYPES. — Madagascar. Antsiranana (Diégo Suarez) Province , Antsiranana II District, bord de la route vers Ramena , 30.I.2004, ♀ immat. fr., Hladik 6820 (P!). — Bord de la route vers Ramena, 30.I.2004, ♂ fl., Hladik 6821 (P!). — Environs de Antsiranana (Diégo Suarez), Montagne des Français, end XII.1924, ♂ fl., Humbert 3956 (P!). — Montagne des Français près d’Antsiranana (Diégo Suarez), I.1926, ♂ fl., Perrier de la Bâthie 17511 (P!). — Mahavanona , Mahagaga , Ambohimarina , à 3 km au nord-est du village Mahagaga , 12°24’35”S, 49°22’19”E, 10.II.2005, ♀ immat. fr., Rakotondrafara, Randrianasolo, Be, Benjara & Jaovoavy 286 (MO!). — Collines et plateaux calcaires de l’Analamera, I.1938, ♂ fl., Humbert 19119 (P!). — Ambilobe District , Anivorano, Maraotoalana, Réserve spéciale d’Ankarana, forêt du Mahoro entrée est, traversée en direction du nord-ouest, 12°49’S, 49°15’E, 7.XI.1997, ♂ fl., Bardot-Vaucoulon 934 (K!, MO, P!) GoogleMaps .
DESCRIPTION
A twining vine to c. 6 m, stems annual from a fleshy tuber. Tubers (only apical part seen) two, probably one per rainy season, to c. 3 cm in diam., vertically oriented, divergent, epidermis brown with irregularly distributed flat, orbicular to elliptic, cream to brown pustules c. 5 mm in diam., crown not clearly differentiated. Indumentum wholly absent, even from young buds. Stems left-twining, to c. 6 mm in diameter, terete and unarmed, white to pink below ground, above pale green to purplish, drying coppery red-brown.
Leaves alternate, blade (30-)41-77 × 23-82 mm, broadly or very broadly ovate to orbicular, sometimes broader than long, chartaceous to thickly so and firm when dry, especially in fruiting specimens, dull green when fresh, drying olive-brown above, paler below, veins (5 or) 7 to the apex, with a bifid vein to each basal lobe, margins entire, base cordate, with a broad, open, shallow sinus 2-13 mm deep, apex rounded and abruptly narrowed into a 6.5-14 mm long acumen bearing a 2.5-3.5 mm long, deltoid to narrowly so forerunner tip, drying concolorous to brown, margins sometimes curling upwards when dry; primary and secondary venation prominent on lower leaf blade surface; petiole ( Fig. 1B View FIG ) 24-76 mm long, 1.7-2.5 mm wide when dry and flat, colour as stem, flattened, channeled on upper surface, margins winged (often folded in herbarium specimens), wing undulate towards base and apex (only clearly visible in living material); cataphylls to 8 mm long, ovate, acuminate, thickly chartaceous, colour as stem, with lateral nodal flanges on either side at base; lateral nodal flanges (“stipules” of Burkill 1960) amplexicaul around node, semicircular, to c. 6 mm in diameter together, continuous with petiole base and undulate in fresh material; bulbils absent. Inflorescences 1 per axil, simple, pendent, lax, axes ridged and angled. Male inflorescences 28-205 mm long (including 6-20 mm long peduncle), axis 0.5-2.1 mm wide, colour as stem, sometimes present on stems bearing only cataphylls, (not true leaves) in November and December, leaves developing later in the rainy season; flowers in cymules at nodes or solitary or in pairs in early stages of development or towards inflorescence apex, nodes fairly regularly spaced c. 5-8 mm apart, cymules either solitary or appearing to be one cymule above the other ( Fig. 1D View FIG ); cymule bracts (1.0-)1.2-2.1 × 0.7-1.3 mm, ovate, membranous, short-acuminate; bracteoles similar but smaller and inserted at 90° to cymule bract; cymule primary branch to 1.8 mm long, only visible in later stages of cymule development. Female inflorescences only known from infructescences, 50- 94 × 1.2-2 mm, peduncle 17-27 mm long, spicate, colour as stem, axis bearing (1-)3-9 capsules. Male flowers rotate at anthesis but tepals ascending to erect before and after anthesis, on (2.5-) 4-9 mm long pedicels (first pedicel of cymule 4.8-9 mm long), angled, thickened only immediately under flower, subtending 0.5-1.3 × 0.2-0.7 mm long, ovate to elliptic or lanceolate, acute to short-acuminate floral bracts; tepal whorls undifferentiated to weakly differentiated (outer narrower), inserted on a flat, discoid to weakly concave, thin torus 0.6-1.1 mm in diam., translucent except in centre where pistillodes are inserted; tepals 6, 0.8-2.0 × 0.6-1.6 mm, free, elliptic to ovate or broadly so or rarely orbicular, pale green, turning brown after anthesis, apex obtuse to acute, thinly chartaceous; stamens erect, filaments 0.1-0.25 mm long, stout, anthers 0.15- 0.4 × 0.15-0.5 mm, broadly orbicular-oblongoid, dorsifixed, making a T-shape with filament, pale yellow; pistillodes c. 1 mm long, 3 conical, erect fleshy structures in centre of torus, inner margins contingent. Female flowers unknown. Capsules 23-37 × 13-18 mm, ascending at c. 30-5° to axis at dehiscence on a 1.5-4.2 × c. 1.5-2 mm, (weakly) clavate, angled capsular stipe, oblong-elliptic to obovate in outline, base rounded to cuneate (immature), apex bluntly acute to truncate or shallowly retuse, tepals persistent in immature capsules but lost by dehiscence; wall divided into three layers (only visible at maturity) exocarp externally pale brown with dense red-brown mottling, extending beyond endocarp on wing margins giving an appearance like a submarginal vein, though the 1.5-3 mm wide exocarp wing tends to curl up at maturity giving the whole capsule a more oblong appearance, mesocarp thin and pithy when dry, only present around axis, endocarp tough, smooth and pale tan inside. Seeds winged primarily at base, but with a c. 1 mm wide, undulate extension to the wing on the outer side and apex of the seed (thus the side of the seed adjacent to the axis is the only unwinged part), seed 7.0-8.5 × 5.0- 6.5 mm excluding wing, flattened ovoid to flattened ovoid-reniform, dark brown, wing 13-15 × 5.5-6.5 mm, oblong, apex obtuse to rounded, membranous, golden-brown.
REMARKS
Bardot-Vaucoulon 934 has 1-1.3 mm long floral bracts, 1.5-2 × 1.1-1.6 mm tepals and 0.3-0.4 × 0.3-0.5 mm anthers. Thus it falls in the high end of the range for all these characters. This variation
B
needs review when more material is available. Male plants of D. buckleyana can flower before their leaves develop, e.g., Bardot Vaucoulon 934, Humbert 3956.
DISTRIBUTION AND ECOLOGY
As Figure 3 shows, D. buckleyana has been encountered at the Réserve spéciale d’Ankarana, Analamera and in and near the Montagne des Français. It appears to be restricted to limestone substrates, with at least six of the nine specimens seen by the authors being from areas of calcareous rock. It is found in deciduous forest at altitudes from 50 to 300 m.
ETYMOLOGY
Dioscorea buckleyana is named for Dr George Buckley to recognise his lifetime of dedication to helping children with disabilities, in particular in the United Kingdom and Romania.
VERNACULAR NAME
Dioscorea buckleyana is known as Taravy or Oviala in the Antsiranana region, or Ovy at Analamera.
USES
Tuber edible.
CONSERVATION STATUS
The extent of occupancy (EOO) of D. buckleyana is 916 km 2, and its area of occurrence (AOO) is 181 km 2 using a cell width of 6.01 km; it occupies five cells (see Willis et al. 2003 and Callmander et al. 2007 for discussion of the importance of cell size). Both of these measures suggest that the appropriate IUCN red list assessment is EN ( IUCN 2001). Rapoport Analysis ( Rapoport 1982) gives an area of 2023 km 2 and three subpopulations. AOO also indicates that there are three subpopulations, as does Figure 3. The exploitation of wild yams in Northern Madagascar is extensive, based on the authors’ field experience there, so decline in numbers of mature plants is likely. As Figure 3 shows, the known populations of this species are in or near protected areas. A provisional IUCN red list assessment of EN B1ab(v); B2ab(v) is proposed.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |