
Conopeum seurati ( Canu, 1928 )
|
publication ID |
https://doi.org/ 10.1080/00222933.2020.1771452 |
|
publication LSID |
lsid:zoobank.org:pub:4FED6730-1C70-4420-B1DA-F1D9046221DF |
|
persistent identifier |
https://treatment.plazi.org/id/2D0787B7-E53C-FFF2-FEA0-FDCDEFD8FD33 |
|
treatment provided by |
Carolina |
|
scientific name |
Conopeum seurati ( Canu, 1928 ) |
| status |
|
Conopeum seurati ( Canu, 1928) View in CoL ( Figures 3 View Figure 3 , 4 View Figure 4 )
Nitscheina spiculata Canu, 1928: 262 , pl. 30, figs 1, 2.
Nitscheina seurati Canu, 1928: 263 , pl. 30, figs 3–6.
Nitscheina fluviatilis Canu, 1928: 263 , pl. 30, fig. 7.
Conopeum seurati: Bobin and Prenant 1962: 381 View in CoL , figs 2, 3; Prenant and Bobin 1966: 127, figs 33, 34 (cum syn.); Jebram 1968: 90, fig. 3, Jebram, 1969a: 225; Jebram, 1969b: 341; Ryland and Hayward 1977: 62, fig. 21; Jebram and Rummert 1978: 502; Poluzzi 1980: 101; Occhipinti Ambrogi 1981: 80, fig. 26; Occhipinti Ambrogi and d’Hondt 1981: 192; Pisano and Matricardi 1983: 311; Poluzzi and Forti 1983: 54, pl. 1, fig 2, pl. 2, figs 1, 2; Occhipinti Ambrogi 1985: 223; Poluzzi and Sabelli 1985: 269, figs 5, 8, 9; Zabala 1986: 244, fig. 55; Poluzzi and Agnoletto 1988: 7, fig. 4A; Zabala and Maluquer 1988: 77, fig. 66; Read and Gordon 1991: 270, fig. 1A; Hayward and Ryland 1998: 124, fig. 24C, D, 25; Rosso 2003: 243; De Blauwe 2009: 172, figs 150–152; Rosso et al. 2010: 596; Ayari and Taylor 2014: 18; Chimenz Gusso et al. 2014: 89, fig. 28a–d; Rosso and Di Martino 2016: 573; Rouse et al. 2018: 143.
Material examined
Mediterranean : NHMUK 2020.1 About NHMUK . 3.1, 36.7505° N, 10.5317° E, 0 m, topotypic specimen from Wadi Bezirk , Tunisia, collected by Rakia Ayari; NHMUK 2020.1 About NHMUK GoogleMaps . 3.2, Wadi Tinja , Tunisia, collected by Rakia Ayari; NMNH A66832 View Materials , labelled Nitscheina spiculata, Wadi Abid , Tunisia, collected by Prof . Léon Gaston Seurat (University of Algiers); NIWA 134508 View Materials , Stn Z18077, 42.9495° N, 10.5439° E, 0 m, Piombino, Tuscany, Italy, coll GoogleMaps . 26 July 2016; NIWA 134512 View Materials , Stn Z18078, 44.8435° N, 12.2898° E, 0 m, on Xenostrobus securis, Goro, Po River delta, Italy, coll GoogleMaps . 23 September 2016; NIWA 134513 View Materials , Stn Z18079, 44.8190° N, 12.3468° E, 0 m, on Xenostrobus securis, Gorino, Po River delta, Italy, coll GoogleMaps . 23 September 2016 by Agnese Marchini and Jasmine Ferrario.
New Zealand: NIWA 98801 View Materials , Stn Z18071, 35.3124° S, 174.1210° E, 0 m, Opua, Bay of Islands, New Zealand, coll GoogleMaps . 31 July 1988 by D.P. Gordon and S .F. Mawatari ; NIWA 98802 View Materials , Stn Z18614, ~ 43.6108° S, 172.7048 E, Lyttelton, coll GoogleMaps . 17 August 1988 by D.P. Gordon and S .F. Mawatari from vessel being cleaned; NIWA 98946 View Materials , Stn Z18081, ~ 40.5569° S, 172.6315° E, Whanganui Inlet, 0 m, on Xenostrobus securis , coll GoogleMaps . January 1963 by O.J. Marston . MITS 1863 , 2 WRE105, 35.7321° S, 174.3258° E, no depth data, Whangarei , 3 November 2005; MITS 1925 , 2 WRE116, 35.7255° S, 174.3271° E, no depth data, Whangarei , 3 November 2005; MITS 66480 , WRE048, 35.7233° S, 174.3255° E, 2 m, Whangarei , 14 November 2002; MITS 66498 , WRE050, 35.7238° S, 174.3253° E, 1 m, Whangarei , 14 November 2002; MITS 66516 , WRE053, 35.7233° S, 174.3255° E, 2 m, Whangarei , 14 November 2002; MITS 66550 , WRE061, 35.7242° S, 174.3270° E, 2 m, Whangarei , 14 November 2002; MITS 66578 , WRE069, 35.7254° S, 174.3285° E, 2 m, Whangarei, 14 November 2002; MITS 66588 , WRE072, 35.7254° S, 174.3285° E, 3 m, Whangarei, 14 November 2002; MITS 71244 , OPX20201, 35.3143° S, 174.1223° E, 6 m, Opua, 18 February 2015 GoogleMaps .
Description
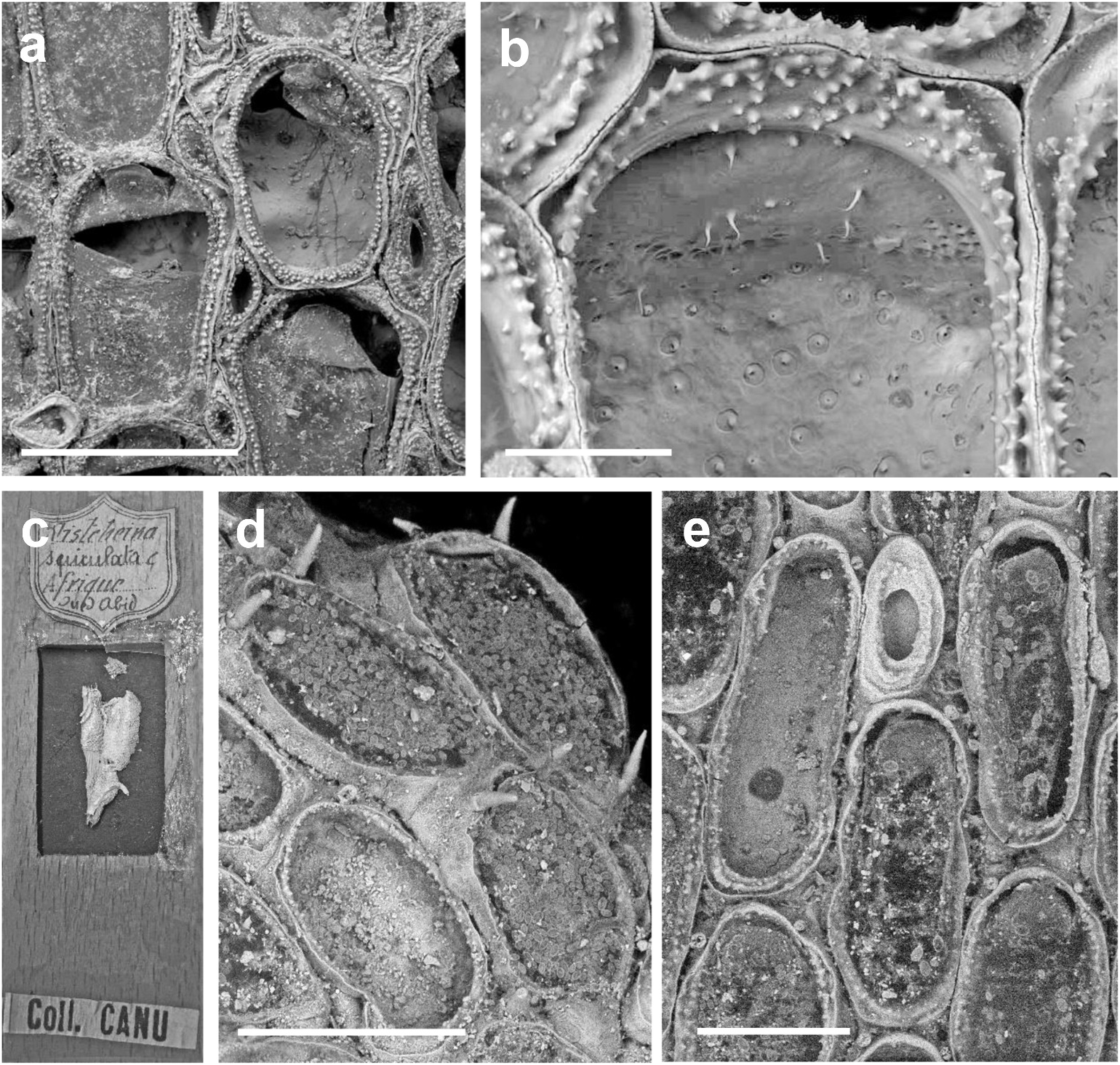
Colony encrusting, typically unilamellar but sometimes self-overgrowing and multiserial. Mature autozooids in zone of astogenetic repetition arranged quincuncially, often twice as long as wide and rounded-subrectangular to elongate-oval, but shape variable depending on position in colony; lateral walls of adjoining zooids closely abutted. Entire frontal area covered by membrane-like cuticularised frontal wall; proximal gymnocyst absent ( Figure 3 View Figure 3 (a)) to little-developed ( Figure 3 View Figure 3 (d,e)), 1–2 rare spine bases in distolateral corners of some zooids ( Figure 3 View Figure 3 (d)). No marginal spines. Basal wall wholly calcified, no window. Operculum at distal end of frontal wall, differentiated within it as a distally rounded flap a little longer than semicircular.
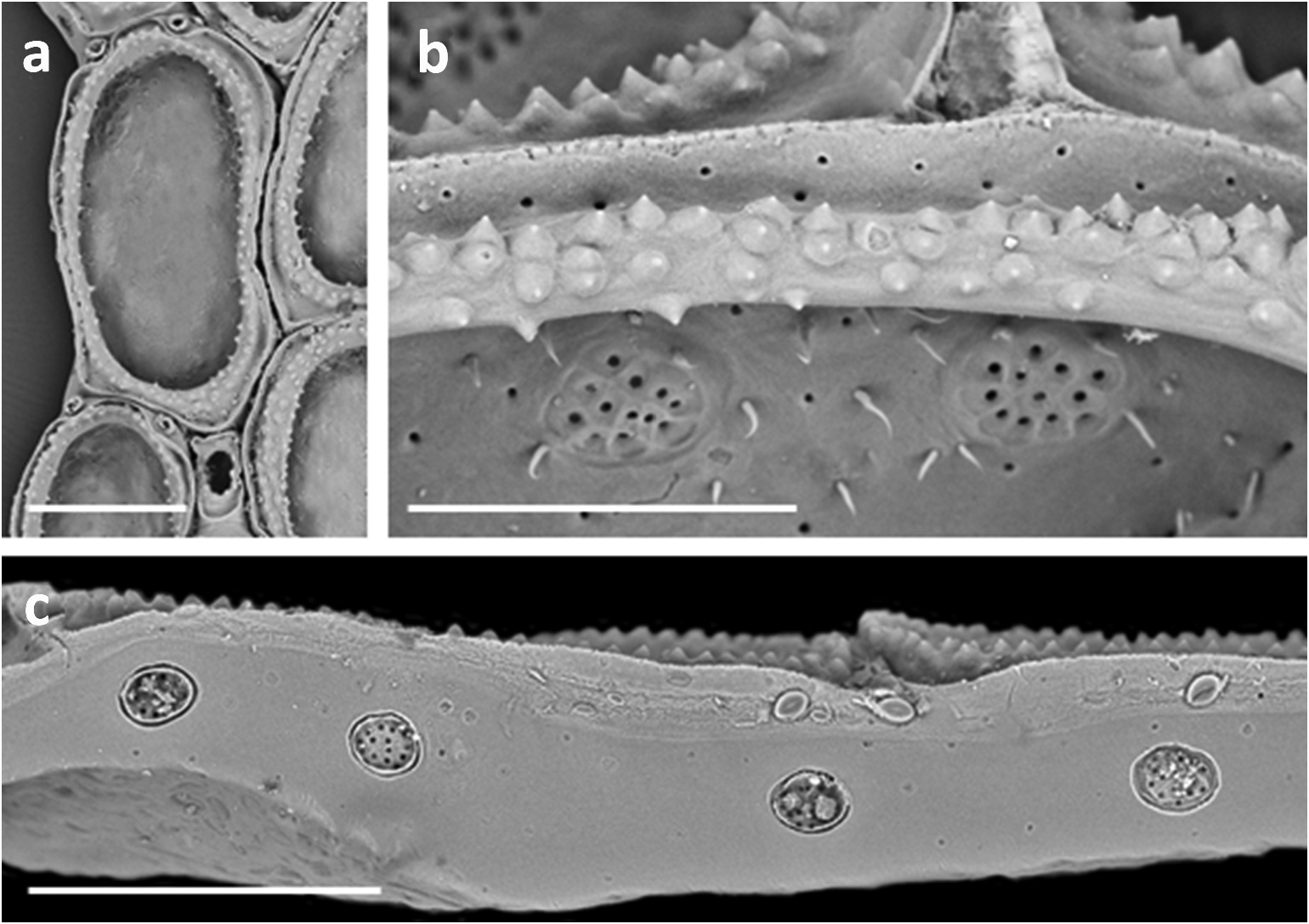
Opesia entirely surrounded by narrow sloping cryptocyst ( Figure 3 View Figure 3 (b)) that is generally of more or less equal width throughout, its surface with conspicuous rounded or pointed granules in 1–4 discontinuous series depending on cryptocyst width; granular part of cryptocyst almost completely surrounded by very narrow furrow ( Figures 3 View Figure 3 (b). 4(a, b)) inside thin zooid rim; furrow constricted or absent mid-distally. In older (ephebic) zooids, granular cryptocyst thicker, with more pustules, and furrow more obvious.
Kenozooids sporadic, small and adventitious ( Figures 3 View Figure 3 (a), 4(a)) or large ones interzooidal ( Figure 3 View Figure 3 (e)), typically inserted in corners between autozooids, small, round, elongate-oval, bean-shaped or spindle-shaped, opesia unevenly surrounded by granular cryptocyst and either no gymnocyst or a tiny exposure.
Lateral interzooidal communications via three multiporous mural septula along each side ( Figure 4 View Figure 4 (c)); larger septula occur in lower half of transverse walls, more or less as a crescentic cluster of pores. Sparse, thin calcareous spinules on inner face of distal transverse wall above septula ( Figure 3 View Figure 3 (b)) or lateral walls either side of septula ( Figure 4 View Figure 4 (b)), often associated with tiny pores; these pores can also occur within the cryptocystal furrow ( Figure 4 View Figure 4 (b)) and in basal walls.
Measurements
Separate measurements were made of specimens from Wadi Bezirk (type locality), the Po River Delta ( Italy) and Whanganui Inlet ( New Zealand):
Wadi Bezirk: ZL 432–722 (574) [N = 28]; ZW 150–501 (331) [N = 36]; OpL 350–653 (520) [N = 18]; OpW 101–357 (266) [N = 24]; PCrL 3–62 (28) [N = 18]; KL 149–471 (289) [N = 10]; KW 84–182 (120) [N = 10]
Po Delta: ZL 342–580 (480) [N = 49]; ZW 260–373 (313) [N = 38]; OpL 274–480 (400) [N = 43]; OpW 181–278 (227) [N = 34]; PCrL 3–62 (28) [N = 18]; ZD 112–184 (143) [N = 4]
Whanganui Inlet: ZL 412–533 (478) [N = 31]; ZW 223–361 (295) [N = 30]; OpL 296–445 (382) [N = 31]; OpW 167–268 (225) [N = 32]; PCrL 20–65 (40) [N = 11]; ZD 133–156 (145) [N = 4]
Remarks
The topotypic specimen described above lacked an ancestrula, but a well-developed specimen from Wadi Tinja has a well-preserved ancestrular region. The ancestrular opesia is elongate-oval with a thin non-granular cryptocystal rim and there are no spines. The ancestrula is surrounded by five zooids. There is a mid-distal daughter zooid, which itself has budded one mid-distal and two distolateral zooids. The original daughter zooid and its two distolateral zooids are connected to two other zooids, directed proximally, that flank the ancestrula; they in turn have produced two proximal zooids that abut the proximolateral corners of the ancestrula. Bobin and Prenant (1962) show the isolated ancestrula (based on a specimen from Essex, England) as subcircular with a pair of distolateral spines, having a more-developed cryptocystal rim than in C. antipodum n. sp.
In comparing C. seurati and C. antipodum , the most obvious distinction, especially evident in SEM micrographs, is the cryptocystal furrow in C. seurati ( Figure 3 View Figure 3 (b)). This furrow is consistent in all Mediterranean specimens that we have seen but completely absent in C. antipodum . The furrow is mostly not apparent in previously published line drawings, exceptions being Jebram (1968, fig. 3) and Poluzzi (1980, fig. 1 inset; reproduced in Poluzzi and Sabelli 1985, fig. 1 inset). It is seen, but has not been especially highlighted, in published SEM micrographs (e.g. Poluzzi 1980, pl. 2, figs 5, 6; Poluzzi and Agnoletto 1988, fig. 4A). It is apparent in specimens from New Zealand ( Figure 4 View Figure 4 (a,b)), confirming the presence of naturalised populations. Excluding material collected from a vessel hull in Lyttelton, C. seurati has been found on anthropogenic substrata at Opua in Northland, in the Town Basin of Whangarei Harbour and in Whanganui Inlet, South Island. Conopeum antipodum n. sp. is by far the commoner species in New Zealand.
The types of the three species from Wadi Bezirk ( N. seurati , N. spiculata , N. fluviatilis ) described by Canu (1928) seem to be no longer extant in the collection of Recent Bryozoa in the MNHM, Paris. Inasmuch as Bobin and Prenant (1962) did not mention having examined them when accepting Borg’s (1931) conclusion that they were all the same species,they may have been missing by that time. However,one of us (EDM) found a historic specimen labelled Nitscheina spiculata ( Figure 3 View Figure 3 (c)) from Wadi Abid, Tunisia, in the palaeontological collection. This was scanned by SEM for the present study ( Figure 3 View Figure 3 (d,e)).
Canu’s (1928) illustrations (light micrographs) are very poor, and inadequate to see much in the way of discriminating details. Borg (1931, p. 24) requested, and was sent by Canu, all three species of Nitscheina (presumably all topotypic) and concluded that they constituted the same variable species. Borg noted that the specimen labelled N. spiculata had zooids in the same colony with and without lateral spines. It is unclear whether Borg (1931) was including the distolateral spines in his description of ‘here and there also rather long lateral spines’. He described N. seurati as being ‘only sparsely provided with lateral spines’, whereas N. fluviatilis was ‘provided with lateral spines that are rather variable in length and sometimes in their upper part more or less curved, but many of the zoids [sic] lack spines’. Borg (1936) redescribed a specimen of N. spiculata from Wadi Abid (probably the one examined for this study, collected in June 1928 by Prof. Seurat). Again, he treated all three species as synonymous, calling them ‘ Membranipora (Electra) crustulenta (Pallas) var. spiculata Canu’. Bobin and Prenant (1962) showed conclusively that this entity ( spiculata / seurati / fluviatilis ) was not conspecific or congeneric with Electra crustulenta (now Einhornia crustulenta ( Pallas, 1766)) . Bobin and Prenant (1962) were the first to use the combination Conopeum seurati , applying it to all three species described by Canu (1928). They noted that N. spiculata had page priority in Canu’s paper (published in November 1928) but rejected use of this species name on the grounds that the combination Nitscheina spiculata was preoccupied – ”le même nom N. spiculata avait été utilise déjà par Canu et Bassler (1923, p. 21) pour une éspèce fossile du Tertiaire américain”. In fact, Canu and Bassler (1923) named the species Membranipora spiculata (which, judging from their photos, is not Membranipora but a species of Electridae or perhaps Calloporidae ). Borg (1931, 1936) was correct in retaining spiculata for the Tunisian- Mediterranean species but so many authors have followed Bobin and Prenant (1962) and Prenant and Bobin (1966) that the combination Conopeum seurati is entrenched in the literature and there seems little point in changing it. [Note that the synonymy given above is deliberately not exhaustive for such a well-known species; additional earlier literature is cited in Prenant and Bobin (1966).]
The contemporary topotypic material from Wadi Bezirk examined by SEM shows no obvious frontal gymnocyst or spines ( Figure 3 View Figure 3 (a)), whereas a historic museum specimen from Wadi Abid labelled Nitscheina spiculata has in the same colony many zooids identical to those from Wadi Bezirk. There are also many zooids in which the cystid walls are raised frontalwards around all or part of the zooid, being developed more distolaterally and/or proximolaterally. A distolateral pair of spines is frequently present, being either very short, having the form of pointed tubercles, or much longer and tapering, with well-developed bases that remain when the spines are broken off. Neither the specimens from Wadi Bezirk nor Wadi Abid seen by us have lateral spines, but they were present on some of the original specimens described from this locality, as elsewhere in the Mediterranean and Western European coasts today. Following Borg (1931, 1936), Bobin and Prenant (1962) and Prenant and Bobin (1966), we accept the synonymy of the three species described by Canu (1928), a conclusion supported by gene sequencing of morphologically variable colonies from elsewhere (see below).
Variations in Mediterranean C. seurati have been documented by different authors, the most comprehensive studies being those of Poluzzi and Forti (1983) and Poluzzi and Sabelli (1985), who described self-overgrowth and boundary interactions, with the formation of erect lobes, some bilamellar. Colonies could also grow around thin algal filaments, forming hollow tubes. Poluzzi and Sabelli (1985) termed some expressions of self-overgrowth as ‘celleporoid layers’ – in which regenerated zooids act as pseudoancestrulae, budding heaps of zooids in loose massive layers of randomly orientated polygonal zooids with smaller polypides. Additionally, both basal and celleporoid layers can include autozooids with closure plates – non-feeding zooids filled with what appeared to be nutritive material.
It is noteworthy that Conopeum seurati in New Zealand has been found associated with Xenostrobus securis , as in Whanganui Inlet; this association was reported for the Italian species sent from the Po River delta for our genetic analysis. Equally, some C. seurati in the Town Basin, Whangarei Harbour, was found encrusting tubes of the naturalised-alien polychaete Ficopomatus enigmaticus ( Read and Gordon 1991) , an association also seen in Tunisia ( Canu 1928), France ( Prenant and Bobin 1966), Italy ( Pisano and Matricardi 1983) and Turkey ( Gugel 1997).
| NMNH |
Smithsonian Institution, National Museum of Natural History |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Conopeum seurati ( Canu, 1928 )
| Gordon, Dennis P., Sutherland, Judy E., Perez, Brenda A., Waeschenbach, Andrea, Taylor, Paul D. & Di Martino, Emanuela 2020 |
Conopeum seurati:
| Rouse S & Loxton J & Spencer Jones ME & Porter JS 2018: 143 |
| Rosso A & Di Martino E 2016: 573 |
| Ayari R & Taylor PD 2014: 18 |
| Chimenz Gusso C & Nicoletti L & Bondanese C 2014: 89 |
| Rosso A & Chimenz C & Balduzzi A 2010: 596 |
| De Blauwe H 2009: 172 |
| Rosso A 2003: 243 |
| Hayward PJ & Ryland JS 1998: 124 |
| Read GB & Gordon DP 1991: 270 |
| Poluzzi A & Agnoletto A 1988: 7 |
| Zabala M & Maluquer P 1988: 77 |
| Zabala M 1986: 244 |
| Occhipinti Ambrogi A 1985: 223 |
| Poluzzi A & Sabelli B 1985: 269 |
| Pisano E & Matricardi G 1983: 311 |
| Poluzzi A & Forti P 1983: 54 |
| Occhipinti Ambrogi A & d'Hondt J-L 1981: 192 |
| Poluzzi A 1980: 101 |
| Jebram D & Rummert H-D 1978: 502 |
| Ryland JS & Hayward PJ 1977: 62 |
| Jebram D 1969: 225 |
| Jebram D 1969: 341 |
| Jebram D 1968: 90 |
| Prenant M & Bobin G 1966: 127 |
| Bobin G & Prenant M 1962: 381 |
Nitscheina spiculata
| Canu F 1928: 262 |
Nitscheina seurati
| Canu F 1928: 263 |
Nitscheina fluviatilis
| Canu F 1928: 263 |