
Gobius xanthocephalus Heymer and Zander, 1992
|
publication ID |
https://doi.org/10.1080/00222930400001533 |
|
publication LSID |
lsid:zoobank.org:pub:4FAC0A48-45C5-40F8-A77D-36922E22B21F |
|
persistent identifier |
https://treatment.plazi.org/id/2E558667-C60B-8E72-FE82-B157FB2EFD5B |
|
treatment provided by |
Felipe |
|
scientific name |
Gobius xanthocephalus Heymer and Zander, 1992 |
| status |
|
Gobius xanthocephalus Heymer and Zander, 1992 View in CoL
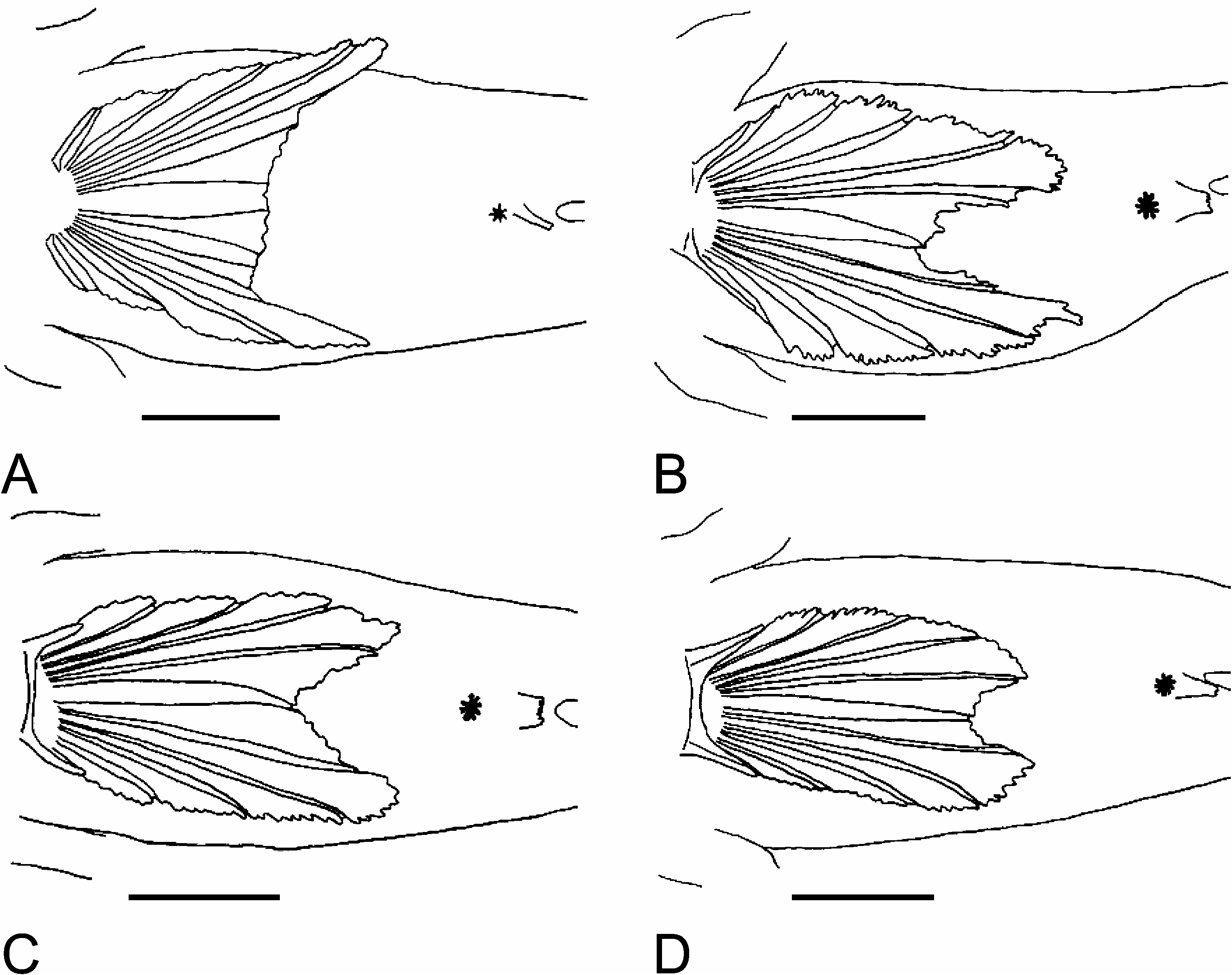
( Figures 2D View Figure 2 , 3D View Figure 3 )
Material
Western Mediterranean: France ( n 53): one juvenile, 26.1 + 7.3 mm, Banyuls-sur-Mer, Ile Grosse , August 1982, R. Patzner ; one juvenile, 28.8 + 9.2 mm, and 1♀, 60.0 + 17.8 mm, Banyuls-sur-Mer, Ile Grosse , September 1992, R. Patzner. Italy ( n 52) : 1♀, 54.0 + 14.7 mm ( NMW 84864, part), and 1 „, 53.7 + d mm ( NMW 84864, part), Gulf of Genova, Savona , 2 August 1984, G. A. C. Balma. Atlantic Ocean: Portugal ( n 54) : 3 „„, 44.4 + 12.0 to 62.1 + 17.2 mm ( NMW 94853), and 1♀, 69.1 + 18.4 mm, Arrabida , south of Lisboa, 2001, E. Gonçalves. Canary Islands ( n 51) : one juvenile 26.2 + d mm ( SMNS 15430 About SMNS ), Tenerife, Playa Paradiso , July 1994, P. Wirtz .
Compared material
Western Mediterranean: France ( n 53): 1♀, 51.9 + 14.1 mm (ZMH 7328, holotype by Heymer and Zander 1992), 1♀, 44.6 + 11.8 mm, and 1 „, 52.7 + 15.2 mm (both ZMH 6195, paratypes by Heymer and Zander 1992), Banyuls-sur-Mer, Ile Grosse, 30 July 1974, A. Heymer.
Identification
Maximum total length about 9 cm. The life coloration of this species is similar to colour morph 2 of G. auratus , but with a pale trunk and red dots forming fewer distinct longitudinal lines along the body. A yellow coloration is characteristic for dorsal parts of the head. Detailed descriptions on the coloration pattern are provided by Heymer and Zander (1994). V short, not reaching anus, with disc only slightly emarginate. C rounded. Important meristics are D2 I/15 (14–15), A I/14 (13–15) and LL 47–50 (44–50).
General morphology
The morphometrics of western Mediterranean and Atlantic material, compared with the types designated by Heymer and Zander (1992) are given in Table II. Detailed morphological descriptions are provided by Heymer and Zander (1992, 1994). Body moderately elongate, trunk laterally compressed. Head large, but length less than 30% of SL, with rather straight ventral side due to relatively straight mouth. Latter short (UJ 10% in SL), not reaching middle of pupil posteriorly. Snout oblique, slightly shorter or equal to eye diameter. Eyes large, diameter about 28% of H. Caudal fin rounded, shorter than head.
Fins (Table III)
D1 VI; D2 I/15 (14–15); A I/14 (13–15); C 14–15 branched rays; P 19–20; V I/5+5/I. Fin bases or lengths given in Table II. D1 without elongate spines, only fifth projecting. Narrow interdorsal space without a membrane. D2 origin anterior of A origin. Rear tips of D2 and A reaching back to C origin, especially in males. C rounded. Longest tip of P reaching back to below D2-origin or D2 1, when the fin is pressed against the body. Three to four uppermost P-rays clearly free from membrane, and with bifid ends. V short, not reaching anus, and not deeply emarginate (V5l about 83% in V4l, Figure 2 View Figure 2 ). Anterior membrane reduced, with a maximum median extension of one-fifth of spine length.
Scales
LL 46–50 (44–50; Table III); TR 14–15 (13–15). Trunk scales mostly ctenoid, but cycloid anterior of imaginary line from dorsal P-base to about middle of D1-base and from ventral P-base to V-origin. Cheek naked. Predorsal area, including nape, upper corner of opercle and pectoral base with small cycloid scales. Larger cycloid scales on breast.
Coloration ( Figure 3D View Figure 3 )
Typical life coloration of G. xanthocephalus is a yellow head and a pale bluish trunk with numerous red to brown dots forming horizontal rows on both. The large pectoral spot, also typical for this species, may not be evident in living specimens. Branchiae not well visible through the opercle. Most distinct row of dots along lateral midline. About three not very distinct rows dorsally, with the median one most evident and originating from the orbit. Two somewhat irregular rows run ventral of the lateral midline row. About four rows forming longitudinal lines on D1 and D2 and vertical lines on C. Predorsal area with many irregularly distributed red dots. Head with one evident row running from the lower orbit to the dorsal P-base, anteriorly continued between orbit and upper lip. Six radial stripes on rim of the eye, confluent on the upper rim, and a V-shaped stripe on the snout, reaching from each orbit to the middle of the upper lip. Three large brown dots between jaw angle and preopercle. Four dark geniohyoid spots and one on the lower lip near jaw angle. After preservation in 70% ethanol, the basic coloration remains a bright brown. The red dots on head and body become substituted by pale mottling. Sometimes rows of small brown dots visible on trunk—one in lateral midline, one above and one below. Dots on median fins sometimes remain grey. Nine to 10 darker and narrow vertical stripes on trunk between Pbase and C-origin, slightly oblique and zigzag-formed. Large black pectoral spot edged white. Cheek spots, geniohyoid spots and dots on lips dark brown and evident. Some small dark dots usually visible on opercle.
Lateral line system
The head canal system is fully developed with anterior and posterior oculoscapular canal and preopercular canal, with pores Ơ, λ, Κ, ω, α, β, ρ; ρ1, ρ2, and γ, δ, E, respectively. The counts of head neuromasts are given in Table IV. Arrangement of superficial head neuromasts resembles that of Miller (1986: as G. auratus ) and Heymer and Zander (1992, 1994). Suborbital row 2 sometimes, row 3 usually reaching longitudinal row d, dividing latter into two or three sections. Row b relatively short, rarely exceeds row 5 anteriorly, therefore latter often confluent. When divided, inferior section 5i relatively long. Row 6i always extends to below level of row d. Row ‘‘ α ’’ always present with at least one, rarely with two papillae near pore α. Last papilla of oculoscapular rows x1 and u sometimes with ventral extension, indicating row tr, but with uncertain assignation. Left and right anterior dorsal row o usually well separated from each other, rarely confluent. Trunk rows as ld1 (11–19 papillae), ld2 (3–9), ld3 (4–8), lv1 (18–30), lv2 (8–20), lv3 (7–18), and as 25–31 ltm (up to 18 papillae, longest rows below D1 and on caudal peduncle). Three longitudinal rows lc (20–46) on caudal fin.
Biology
Gobius xanthocephalus shows a strong benthic behaviour. It is abundant on gently sloping bedrock coasts at Banyuls-sur-Mer, observed in depths between 3 and 12 m. The specimens can be found resting on the rocks or on small sandy patches in between.
Molecular genetic analysis
Of 379 bp sequenced, 344 characters were constant, 21 were parsimony-uninformative, and only 14 were parsimony-informative. The observed genetic variation turned out to be very low among all ingroup taxa (zero to eight mutations) and showed no insertion or deletion events, neither within nor between morphologically defined taxa. Thus, all taxa share common ancestry in the recent past. Parsimony analysis resulted in 125 most parsimonious trees of a length of 45 mutations (consistency index excluding uninformative sites, 0.7083; retention index, 0.7879, trees not shown). The neighbour-joining tree ( Figure 6A View Figure 6 ) was identical to one of the most parsimonious trees. The latter was used to construct the minimum spanning tree ( Figure 6B View Figure 6 ). Within the 30 specimens analysed of G. fallax , G. auratus and G. xanthocephalus , 18 different haplotypes were found, 15 of which are each represented by only one specimen ( Figure 6 View Figure 6 ). The remaining three occurred in more than one specimen, but only one of them was shared intraspecifically. More than onethird of all specimens investigated shared the same haplotype. This haplotype occurred not only in seven specimens of G. fallax but also in four of the colour morph 2 specimens of G. auratus . These G. fallax specimens originated from the Gulf of Trieste (Piran), from Pula and the Kvarner region (Cres), while all of the four G. auratus stem from the Kvarner region (Cres and Krk). The remaining specimens of G. fallax were placed in two other haplotype clusters. Apart from the most common haplotype, G. auratus was irregularly distributed over the minimum-spanning tree and also showed a second haplotype shared with G. fallax . Four out of five specimens of colour morph 1 of G. auratus clustered close to the most common haplotype, differing only by one or two base substitutions, but the fifth was placed within the second major haplotype cluster, which is mainly represented by G. fallax from the Kvarner region. The two specimens of G. xanthocephalus from Portugal were separated from each other by one substitution and represented the only intraspecific cluster, which differed by five unique mutations from all other specimens and by seven to eight mutations from the most frequent haplotype found in both G. fallax and G. auratus .


| NMW |
Naturhistorisches Museum, Wien |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.