
Physalaemus claptoni, Leal & Leite & Da Costa & Nascimento & Lourenço & Garcia, 2020
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4766.2.3 |
|
publication LSID |
urn:lsid:zoobank.org:pub:385FD3DD-9E4E-4FE4-A3D9-7952F012A963 |
|
DOI |
https://doi.org/10.5281/zenodo.3803675 |
|
persistent identifier |
https://treatment.plazi.org/id/82FFC419-8465-4403-ADE9-289FDED82D92 |
|
taxon LSID |
lsid:zoobank.org:act:82FFC419-8465-4403-ADE9-289FDED82D92 |
|
treatment provided by |
Carolina |
|
scientific name |
Physalaemus claptoni |
| status |
sp. nov. |
Physalaemus claptoni , new species
urn:lsid:zoobank.org:act:82FFC419-8465-4403-ADE9-289FDED82D92
Holotype ( Figs. 1–3 View FIGURE 1 View FIGURE 2 View FIGURE 3 ; Table 1 View TABLE 1 ). UFMG 16784 View Materials adult male, November 15, 2014, Serra do Cipó , Fazenda do Chiquito (19.166917° S, 43.529000° W; 1321 m above sea level; datum WGS84), Municipality of Santana do Riacho, State of Minas Gerais, Brazil, F. Leal and H. Thomassen. GoogleMaps
Paratypes ( Figs. 4–6 View FIGURE 4 View FIGURE 5 View FIGURE 6 , Table 1 View TABLE 1 ). Eighteen adult males: UFMG 16773, 16774, 16776–16783, 16785–16792; one adult female: UFMG 16775, all collected with the holotype by F. Leal and H. Thomassen. Six adult males: UFMG 5418–5423, collected at the type locality on November 1, 2010 by P. C. A. Garcia and F. Natali. One adult male: UFMG 5585, collected at the type locality on November 3, 2010 by F. Leal and P. Taucce.
Referred specimens. Two adult males: UFMG 12999–13000, collected at the type locality on December 1, 2011 by P. C. A. Garcia; the individuals were subjected to cytogenetic analyses. One adult male: UFMG 10120, collected at the type locality on January 28, 2012 by P. C. A. Garcia, J. Thompsom, and L. Durães.
Diagnosis. The new species is distinguished from all the congener species by the following combination of character states: (1) presence of an arrow-shaped blotch on the dorsum of the body ( Figs. 1A View FIGURE 1 , 3 View FIGURE 3 , 4 View FIGURE 4 , 6A View FIGURE 6 ); (2) presence of a median longitudinal light stripe over the urostyle region ( Figs. 1A View FIGURE 1 , 3 View FIGURE 3 , 4 View FIGURE 4 , 6A View FIGURE 6 ); (3) belly and ventral surface of thighs marbled with dark gray irregularly shaped blotches, on a bluish background ( Fig. 5 View FIGURE 5 ); (4) absence of reddish coloration over the axillar and inguinal regions in live individuals; (5) bright orange dots scattered on the dorsum of the head, upper lip, and body, in live individuals ( Figs. 3 View FIGURE 3 , 4 View FIGURE 4 ); (6) ventral surface of hand and foot red in live individuals; (7) small size (adult male SVL= 16.2–18.2 mm, Table 1 View TABLE 1 ); (8) presence of a brown, non-divided, nuptial pad in males ( Fig. 7A View FIGURE 7 ); (9) END/ED in males ranging from 0.85–0.93 ( Table 1 View TABLE 1 ); (10) supernumerary tubercles on foot absent ( Fig. 2D View FIGURE 2 ); (11) tarsal fold absent ( Fig. 1B View FIGURE 1 ); (12) tarsal tubercle absent; (13); texture of posterior region of belly and ventral surface of thigh smooth ( Fig. 1B View FIGURE 1 ); (14) advertisement call composed of two note types (note A + B); (15) advertisement call duration of 0.80– 1.28 s ( Fig. 8A View FIGURE 8 ; Table 2 View TABLE 2 ); (16) note A with ascending amplitude until mid-note then descending towards the end of the note; (17) note B with pulses arranged into 5–7 groups; (18) dominant frequency of note A from 1734.4–2765.6 Hz; and (19) dominant frequency of note B from 1507.3–2859.4 Hz ( Fig. 8 View FIGURE 8 ).
Comparison with other species. The presence of an arrow-shaped blotch on the dorsum of the body distinguishes Physalaemus claptoni sp. nov. from all the species of the Physalaemus cuvieri Clade (sensu Lourenço et al. 2015). Additionally, the absence of tarsal tubercle distinguishes P. claptoni sp. nov. from the species of the Physalaemus cuvieri Clade, except P. lateristriga , P. olfersii , P. orophilus , and P. maximus of the Physalaemus olfersii species group, from which the new species is promptly distinguishable by the smaller size of adult males at 16.2–18.2 mm ( P. lateristriga , P. olfersii , P. orophilus , and P. maximus combined SVL of adult males= 23.1–44.8 mm). Within the P. signifer Clade, the presence of a longitudinal light stripe over the urostyle region distinguishes P. claptoni sp. nov. from all the other species, except from P. rupestris ( Fig. 9 View FIGURE 9 , S 1 View FIGURE 1 ), which also has this stripe over the urostyle region ( Fig. 9A View FIGURE 9 ). By having belly and ventral surface of thigh marbled with dark gray irregularly shaped blotches on a bluish background in live specimens, P. claptoni sp. nov. is set apart from P. angrensis , P. atlanticus , P. nanus , and P. spiniger (belly with flashy orange blotches in those species), from P. maculiventris (which has a pale chest and belly, with posterior region of belly and ventral surface of thigh showing bold black blotches), and from P. obtectus (which has belly with flashy red blotches). The absence of reddish coloration over the axillar and inguinal regions in live individuals distinguishes P. claptoni sp. nov. from P. deimaticus and P. erythros (reddish coloration present over the axillary and inguinal regions on those species). The bright orange dots scattered on the dorsum of the head, upper lip, and body in live specimens of P. claptoni sp. nov. distinguishes it from all the other species within the P. signifer Clade. Ventral surfaces of hand and foot are red in live individuals, which distinguishes P. claptoni sp. nov. from P. angrensis , P. atlanticus , P. maculiventris , P. spiniger (ventral surface of hand and foot orange in these species), and from P. bokermanni , P. caete , P. camacan , P. irroratus , P. moreirae , P. nanus , and P. nattereri (which lack aposematic coloration on ventral surfaces of hand and foot). The small size of P. claptoni sp. nov. adult males at 16.2–18.2 mm promptly distinguishes it from P. atlanticus , P. caete , P. camacan , P. crombiei , P. deimaticus , P. erythros , P. irroratus , P. maculiventris , P. moreirae , P. nattereri , and P. obtectus (combined SVL= 18.9–50.6 mm). The presence of a non-divided nuptial pad in males distinguishes P. claptoni sp. nov. from P. angrensis , P. atlanticus , P. bokermanni , P. caete , P. camacan , P. crombiei , P. deimaticus , P. erythros , P. moreirae , P. nanus , P. obtectus , P. rupestris ( Fig. 7B View FIGURE 7 ), P. signifer , and P. spiniger (nuptial pad divided in those species). Additionally, P. claptoni sp. nov. is distinguished from P. rupestris by having brown nuptial pad (nuptial pad white-cream in P. rupestris ; Fig. 7B View FIGURE 7 ) and by the proportion between END/ED in males ranging from 0.85–0.93 (END/ED of P. rupestris males ranges from 0.75–0.82). The absence of supernumerary tubercles on foot distinguishes P. claptoni sp. nov. from P. angrensis , P. caete , P. camacan , P. crombiei , P. irroratus , P. moreirae , P. signifer , and P. spiniger (supernumerary tubercles present on foot of those species). The absence of a tarsal fold distinguishes P. claptoni sp. nov. from P. angrensis , P. atlanticus , P. bokermanni , P. caete , P. camacan , P. crombiei , P. irroratus , P. moreirae , P. nanus , P. obtectus , P. signifer , and P. spiniger (tarsal fold present in those species). The smooth texture of posterior region of belly and ventral surfaces of thigh in P. claptoni sp. nov. distinguishes it from P. camacan and P. irroratus (posterior region of belly and ventral surface of thigh granulated in those species). The advertisement call of P. claptoni sp. nov. is composed of two note types (notes A and B), setting it apart from P. atlanticus , P. angrensis , P. bokermanni , P. camacan , P. crombiei , P. erythros , P. maculiventris , P. moreirae , P. obtectus , and P. signifer (advertisement call has only one note type in those species). The advertisement call of P. claptoni sp. nov. and P. rupestris are both composed of two note types (A and B), however are promptly distinguishable from each other by its duration, which ranges from 0.80– 1.28 s in P. claptoni sp. nov. and from 1.51– 2.20 s in P. rupestris . Additionally, note A of P. claptoni sp. nov. has pulses with an ascending amplitude until mid-note and then descends towards the end of the note, while note A of P. rupestris has only descending amplitude modulation; note B of P. claptoni sp. nov. has pulses arranged in 5 to 7 pulse groups, whereas note B of P. rupestris has pulse grouping only at the onset and final portion of the note. Dominant frequency of note A in P. claptoni sp. nov. varies from 1734.4–2765.6 Hz and in P. rupestris from 3046.9–3421.9 Hz. Dominant frequency of note B in P. claptoni sp. nov. ranges from 1507.3–2859.4 Hz and in P. rupestris from 3046.9–3421.9 Hz.
Description of the holotype. Adult male, SVL 17.3 mm ( Fig. 1–3 View FIGURE 1 View FIGURE 2 View FIGURE 3 ). Head slightly longer than wide. Head width (32.4% SVL) slightly shorter than head length (34.7% SVL). Snout short, rounded in dorsal and lateral views ( Fig. 2 View FIGURE 2 ). Canthus rostralis distinct, rounded; loreal region slightly concave. Snout protruding beyond lower jaw. Nostril dorsolaterally oriented, faintly protruding. Internarial region slightly concave; top of the head flat. Eye slightly prominent, antero-laterally oriented, its diameter 11% bigger than END. Tympanum indistinct externally. Supratympanic fold distinct, thick, extending from the posterior corner of the eyes to the shoulders. Dentigerous process of vomer absent. Premaxillary and maxillary teeth absent. Choanae rounded, separated from each other by a distance as large as four times its diameter. Tongue elliptical, elongated, slightly constricted in its middle, free around lateral and posterior margin. Vocal slit present, longitudinal, originating on the sides of the tongue in its anterior third, and extending towards the corner of the mouth. Vocal sac single and subgular, weakly differentiated externally, slightly folded over the chest (in preservative). Dorsolateral fold present, weakly distinct, from the posterior corner of the eye to the inguinal region. Forearm very hypertrophied in relation to upper arms; upper arms slender, short. Fingers thick, without webs, relative lengths I=II=III <IV; fingers tip slightly dorsal-ventrally dilated, not expanded laterally. Proximal subarticular tubercles very large, simple, prominent and rounded; distal subarticular tubercle present on fingers III and IV, smaller and less prominent than the proximal tubercles; supernumerary tubercles large, rounded, present at base of fingers II, III and IV. External metacarpal tubercle ovoid, large and prominent. Inner metacarpal tubercle ovoid, large and prominent. Nuptial pad not divided, presenting dark keratinized spicules densely covering the dorsal and lateral surfaces of the thumbs (except for the distal phalange) and ventrally cover the external half of the internal metacarpal tubercles ( Fig. 7A View FIGURE 7 ). Tibia length 40% SVL; foot length 64% SVL. Toes thick, without webs, relative lengths I <II=V <III <IV; toes tip slightly dorsal-ventrally dilated, not expanded laterally. Subarticular tubercles distinct, simple, prominent and rounded; supernumerary absent. Tarsal fold absent; tarsal tubercle absent. Inner metatarsal tubercle distinct, ovoid; outer metatarsal tubercle distinct, rounded. Inguinal gland well developed, oval. Cloacal opening directed posteriorly at upper level of thigh. Region below the cloaca very granular, tubercles relatively large and slightly square-shaped, reaching the thighs at its posteroventral edge, where tubercles become increasingly smaller and lower. In preservative, the texture of the hidden and ventral surfaces are smooth, the dorsal surfaces of the head, body and limbs are slightly rugose, the edge of the eyelid bears a few very low tubercles, and the flank has small tubercles scattered; in life, the flank and dorsal surfaces of the head, body and thigh are granulated.
Measurements (in mm) of the holotype: SVL 17.3; HL 6.0; HW 5.6; ED 1.6; AMD 3.1; ESD 2.5; END 1.4; IND 1.4; THL 7.6; TL 7.0; FL 11.0; HAL 4.1.
Color of the holotype in life. Dorsal background color of the head, body, upper arms, and forearms light orange. Interorbital blotch leaden gray colored, irregularly shaped, curved towards the body, extending from the superior edge of one eyelid to the other; anterior margin well-defined, bordered by a very thin white line; posterior margin diffuse. Dorsum of the body with an arrow-shaped blotch medially located, gray leaden, and outlined by a very thin white line; its posterior portion extends transversely toward the inguinal gland, covering the sacrum region; a small white spot is displayed on the middle of the arrow-head and there is a longitudinal light stripe over the urostyle region. Black ocelli cover 75% of the inguinal gland on its posterior portion. The dorsal and hidden surfaces of thigh, tibia and tarsus leaden gray colored; dorsal surface of the toes is light orange; transversal black bar on the dorsal surfaces of thigh, tibia, and foot, aligned to each other, slightly invading the posterior surface of the thigh and ventral surface of the tibia. Irregularly shaped black blotches on the heel, knee, and ventral surface of tarsus. Longitudinal elongated black blotch on the posterior surface of upper arm, extending slightly dorsally to the elbow. Hand and distal half of the forearm with small black spots and scattered orange dots. Dorsolateral gray leaden-colored blotch, extending from the posterior margin of the eye, passing over the supratympanic fold and almost reaching the inguinal region; stained by black spots on the supratympanic fold and on the posterior half of the flank. Canthus rostralis, upper half of the loreal region and lateral surfaces of the snout stained by small black spots; supralabial region bright orange colored. Black blotches under the eye separated from each other by an orange surface. Dorsum of the body and head with scattered bright orange dots. Ventral surfaces of the limbs marbled with dark gray and bluish irregular shaped spots; black blotch over the glandular tissue on the ventrolateral edge of forearm. Black bar above the cloaca, curved down toward the posterior surface of the thighs. Gular region and chest dark gray, stained with scattered bluish dots. Belly and ventral surface of thigh marbled with dark gray irregularly shaped blotches, on a bluish background. Ventral surface of the hand red; toes I and II, external metatarsal tubercles, internal metatarsal tubercle and subarticular tubercles of the foot red; plantar region and region between subarticular tubercles of the toes III, IV and V black with scattered irregular shaped whitish spots. Iris orange, adorned with black vermiculation; pupil black and horizontal.
Color of the holotype in alcohol 70%. In preservative, all the dorsal coloration took on dark gray shades; the dorsal blotches become barely distinguishable; all the orange coloration faded. Ventrally, the black and dark gray colorations become brownish dark gray, bluish coloration become pale white; and the red coloration of the hand and foot become pale white.
Variation. Measurements of 26 adult males and one adult female are presented in Table 1 View TABLE 1 . The adult female ( Fig. 6 View FIGURE 6 ) differs from adult males by having slender forearm; vocal slit absent; nuptial pad absent; and HL and HW smaller in relation to SVL. The left hand of the female paratype is malformed, with smaller fingers. In life, color of dorsal background of head, body, upper arm, and forearm may vary from pale yellow to bright orange; color of interorbital, dorsal and dorsolateral blotches as well as dorsal and hidden surfaces of thigh, tibia and tarsus may be brown or dark gray; the orange dots on the dorsum of head and body vary in number and intensity of color, being pinkish in some individuals; the area with orange coloration over upper lips and canthus rostralis varies in size ( Fig. 4 View FIGURE 4 ). Interorbital blotch may be M shaped; tip of the arrow-shaped blotch may be in contact with the interorbital blotch. Median longitudinal light stripe over the urostyle region varies in size and thickness. Irregularly shaped blotches of belly and ventral surface of thigh may vary in size and number. Iris varies from pale yellow to orange. In preservative, the current (2019) general coloration of specimens collected in 2010 is brownish, whereas the ones collected on 2014 remain grayish. Tympanum may be slightly discernible externally in some individuals. Head may be longer than wide or as long as wide. Dark superficial keratinized layer of nuptial pad may be peeled, but nuptial pad remains visible.
Etymology. The specific epithet is used as a noun and honors Eric Clapton, the English rock and blues guitarist, singer, and songwriter. Clapton is widely recognized as one of the best guitarists of all time. Over his career, Clapton’s work has made an outstanding contribution to music, influencing many musicians all around the world.
Advertisement call. The advertisement call of Physalaemus claptoni sp. nov. ( Fig. 8A, B View FIGURE 8 ; Table 2 View TABLE 2 ; n=45 calls from five males; Audio S1) consists of two types of multipulsed notes (herein referred as notes A and B) released in sequence. It is formed by 65–93 pulses, has duration ranging from 0.80– 1.28 s (n=45 advertisement calls), interval between notes A and B from 0.10– 0.19 s (n=45 intervals) and between advertisement calls varying from 0.14– 1.38 s (n=41 intervals). Note rate within advertisement call varied from 1.56–2.51 notes/s (n=90 notes). Note A (n=154 notes) is shorter than B, with duration varying from 0.11– 0.16 s, is formed by 12–17 pulses emitted at a rate of 86.7–119.3 pulses/s. Note A was released at a rate of 0.53–0.63 notes/s. Pulses of note A are homogeneously distributed and have ascending amplitude until mid-note then descending towards the end of the note, being fairly symmetric (peak amplitude from 0.44–0.59). Notes A were also observed being released in sequences without note B, and, sometimes, between advertisement calls. In this case, we considered each note A as a single call. The interval between single A note calls emitted in sequences ranged from 0.7– 1.9 s (n=113 intervals). Note B (n=45 notes, from four males) duration varies from 0.67– 1.01 s and is formed by 53–81 pulses emitted at a rate of 67.5–92.1 pulses/s. Note B was released at a rate of 0.32–0.61 notes/s. Pulses of note B are arranged in 5–7 pulses group, with relative amplitude lower than that of note A. Within those groups, pulses are also homogeneously distributed and with ascending amplitude until its midpoint, then descending towards the end of the pulse group. The first and second pulse groups are generally closer and even in contact to each other than are the others (See Fig. 8 A View FIGURE 8 ). The other pulse groups of note B may have a complete amplitude modulation in between groups, or not. In the latter case, there are a few isolated and low-amplitude pulses in between groups. Frequency bandwidth of note A (n=154 notes) is between 1453.1–3140.6 Hz and of note B (n=45 notes) is between 1335.1–3375.0 Hz. Dominant frequency of note A (n=154 notes) ranged from 1734.4–2765.6 Hz and of note B (n=45 notes) from 1507.3–2859.4 Hz.
Phylogenetic inferences and genetic distances. The phylogenetic analysis recovered one most parsimonious tree, with 5853 steps. In the inferred cladogram, Physalaemus claptoni sp. nov. was nested inside the P. deimaticus species group, a clade that was highly supported (bootstrap value of 100%; Fig. 10 View FIGURE 10 ). In this analysis, the P. signifer Clade and the P. cuvieri Clade were recovered, the former with high support ( Fig. 10 View FIGURE 10 ). Physalaemus rupestris was inferred to be the sister species of P. claptoni sp. nov., but with no bootstrap support, and the clade composed of P. claptoni sp. nov. and P. rupestris was the sister group of a clade that included P. deimaticus and P. erythros ( Fig. 10 View FIGURE 10 ).
Uncorrected p -distances between the species of the Physalaemus deimaticus group ranged from 3.5% to 5.9% and from 2.4% to 5.6% based on the mitochondrial H1 fragment and the 16Sar-16Sbr fragment, respectively ( Table 3 View TABLE 3 ). The genetic distances between P. claptoni sp. nov. and each of the other species in the P. deimaticus group were high, with the minimum value being 4.9% for the H1 fragment and 4.6% for the 16Sar-16Sbr segment ( Table 3 View TABLE 3 ). The intraspecific distances were very low for both analyzed fragments and all the species of the P. deimaticus group ( Table 3 View TABLE 3 ).
Distribution and natural history. Physalaemus claptoni sp. nov. is so far only known from its type locality in the highlands of the Serra do Cipó, southern Espinhaço Range, State of Minas Gerais, southeastern Brazil ( Fig. 11 View FIGURE 11 ). The new species reproduces mostly on small still water collections associated with the edge of a hilltop forest fragment surrounded by campo rupestre (or rupestrian grasslands) that becomes slightly flooded during the rainy season. When collected on the outside border of the forest fragment, males were observed calling from the base of dense grass. When captured inside the forest fragment, males were hidden on the leaf litter, generally close to or inside small puddles. Due to the small species size and difficult access to calling sites, we struggled to collect the specimens to make the description, with at least seven field trips from 2010 to 2017. Most of the type series were collected in 2014 when the open area around the forest fragment was burned right before the rainy season, strongly decreasing the density and size of the grass, making it easier to see and collect the calling males. Moreover, this occasion was the one with the densest fog and was when more calling males could be identified, with approximately 30 individuals. During the other expeditions to the same site, the number of calling males varied from zero to approximately 12 individuals.
Advertisement call of Physalaemus rupestris . The advertisement call of Physalaemus rupestris ( Fig. 8C, D View FIGURE 8 ; Table 2 View TABLE 2 ; n=44 calls from three males; Audio S2) consists of two types of multipulsed notes (herein referred as notes A and B) released in sequence. It is formed by 52–85 pulses, has duration ranging from 1.51– 2.2 s (n=44 advertisement calls), interval between notes A and B from 0.22– 0.42 s (n=44 intervals) and between advertisement calls varying from 0.26– 1.75 s (n=24 intervals). Note rate within advertisement call varied from 0.91–1.32 notes/s. Note A (n=93 notes, from three males) is shorter than B, with duration varying from 0.05– 0.10 s, is formed by 8–11 pulses emitted at a rate of 115.4–183.7 pulses/s. Note A was released at a rate of 0.42–0.76 notes/s. Pulses of note A present descending amplitude modulation and are homogeneously distributed, with the exception of the last two or three, which are increasingly separate to each other. Notes A were also observed being released in sequences without note B, and, sometimes, between advertisement calls. In this case, we are considering each note A as a single call. The interval between single A note calls emitted in sequences ranged from 0.22– 5.27 s (n=64 intervals). Note B (n=44 notes, from three males) duration varies from 1.14– 1.78 s and is formed by 41–75 pulses emitted at a rate of 31.4–43.1 pulses/s. Note B was released at a rate of 0.20–0.34 notes/s. Pulses of note B present descending amplitude modulation and are homogeneously distributed, with the exception of the first three to five, which are usually closer to each other, and the last eight to fifteen, which are increasingly close to each other. In the middle of note B, some pulses may be closer to each other than others, but it occurs with just a few pulses and seemingly at random. Frequency bandwidth of notes A (n=93 notes) and B (n=44 notes) are from 1875.0–3750.0 Hz. Dominant frequency of notes A and B ranged from 3046.9–3421.9 Hz.






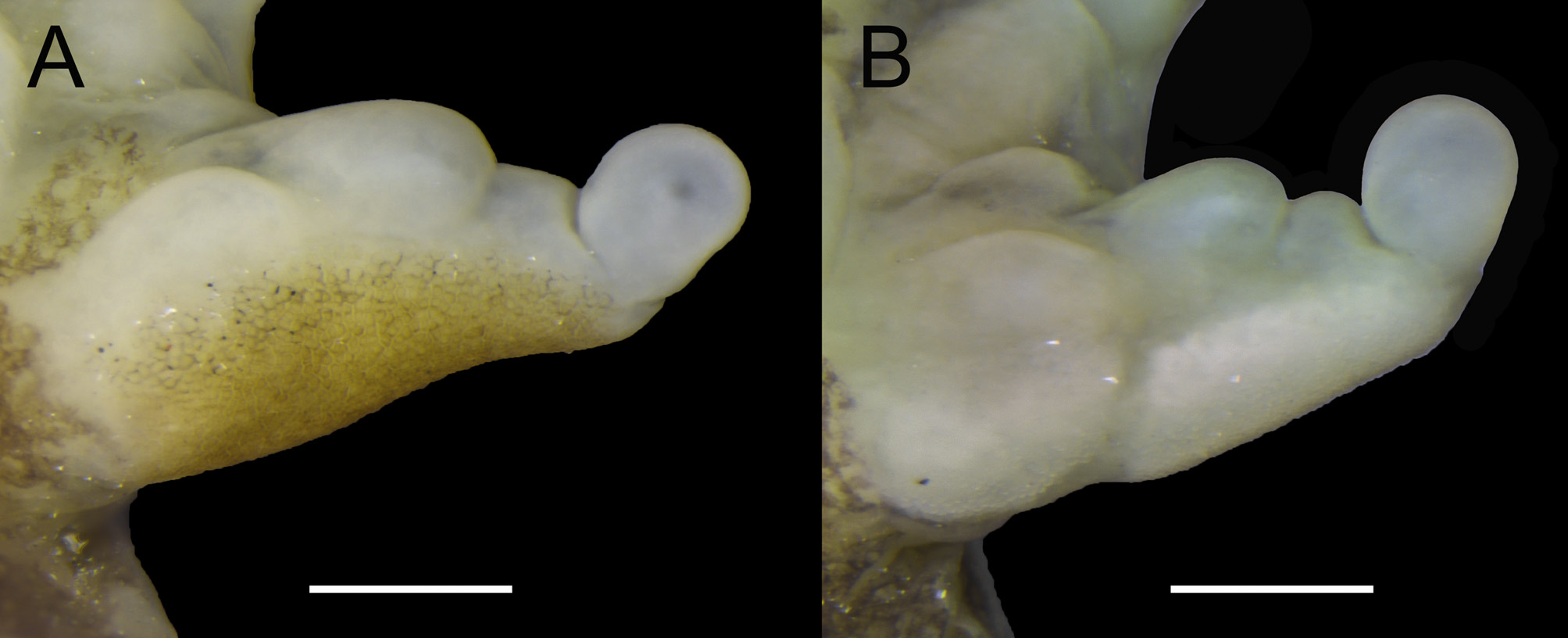
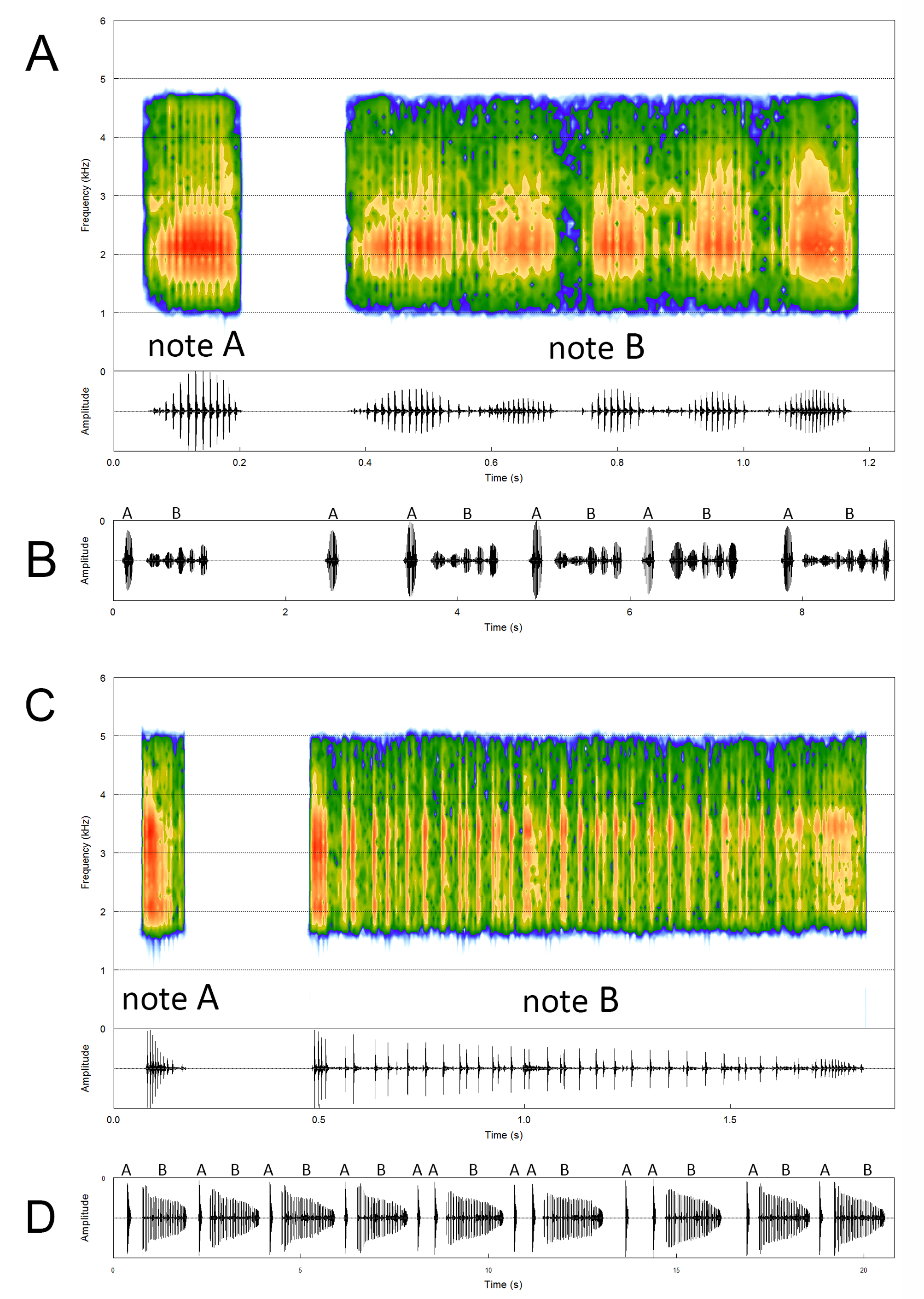

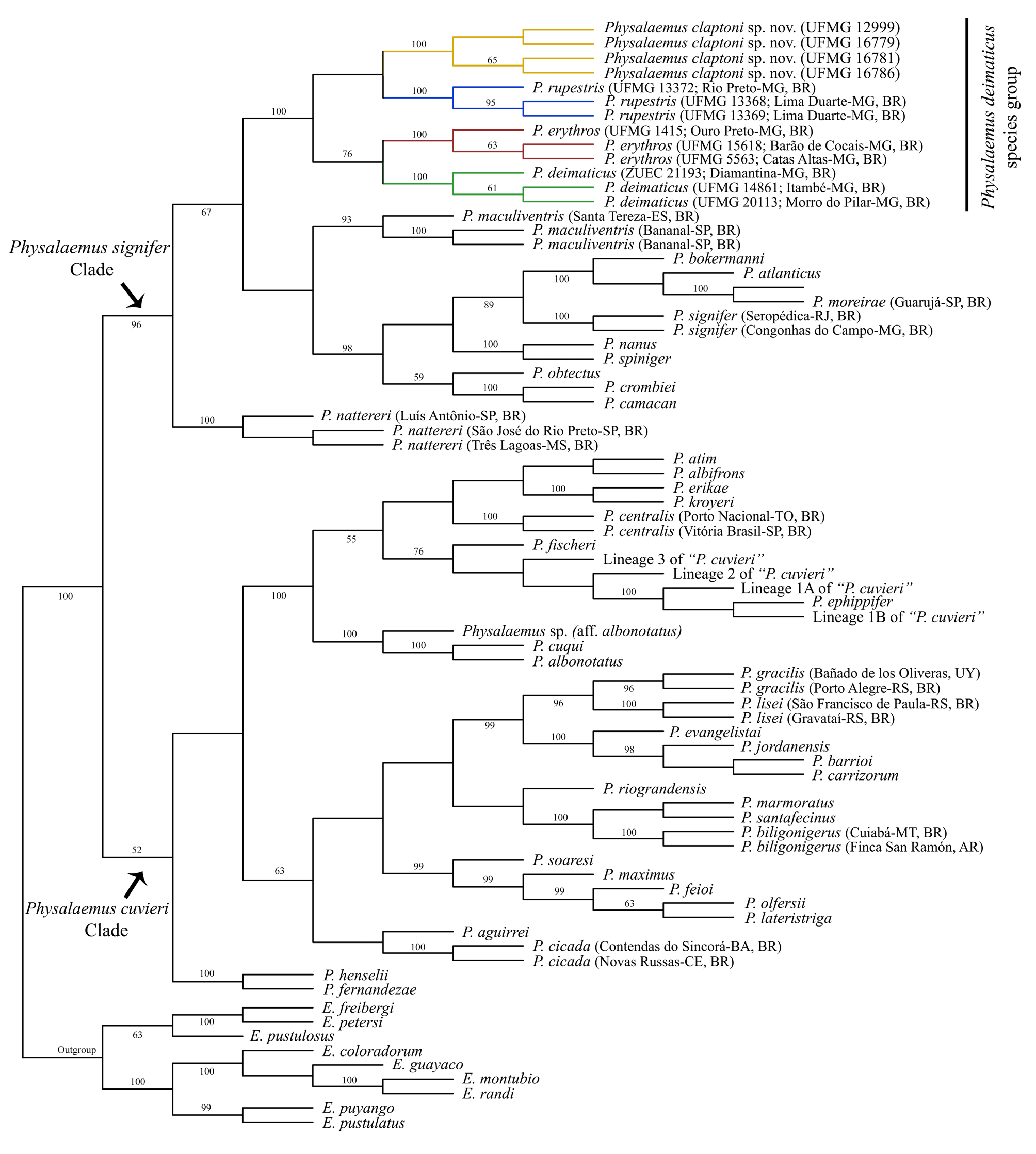
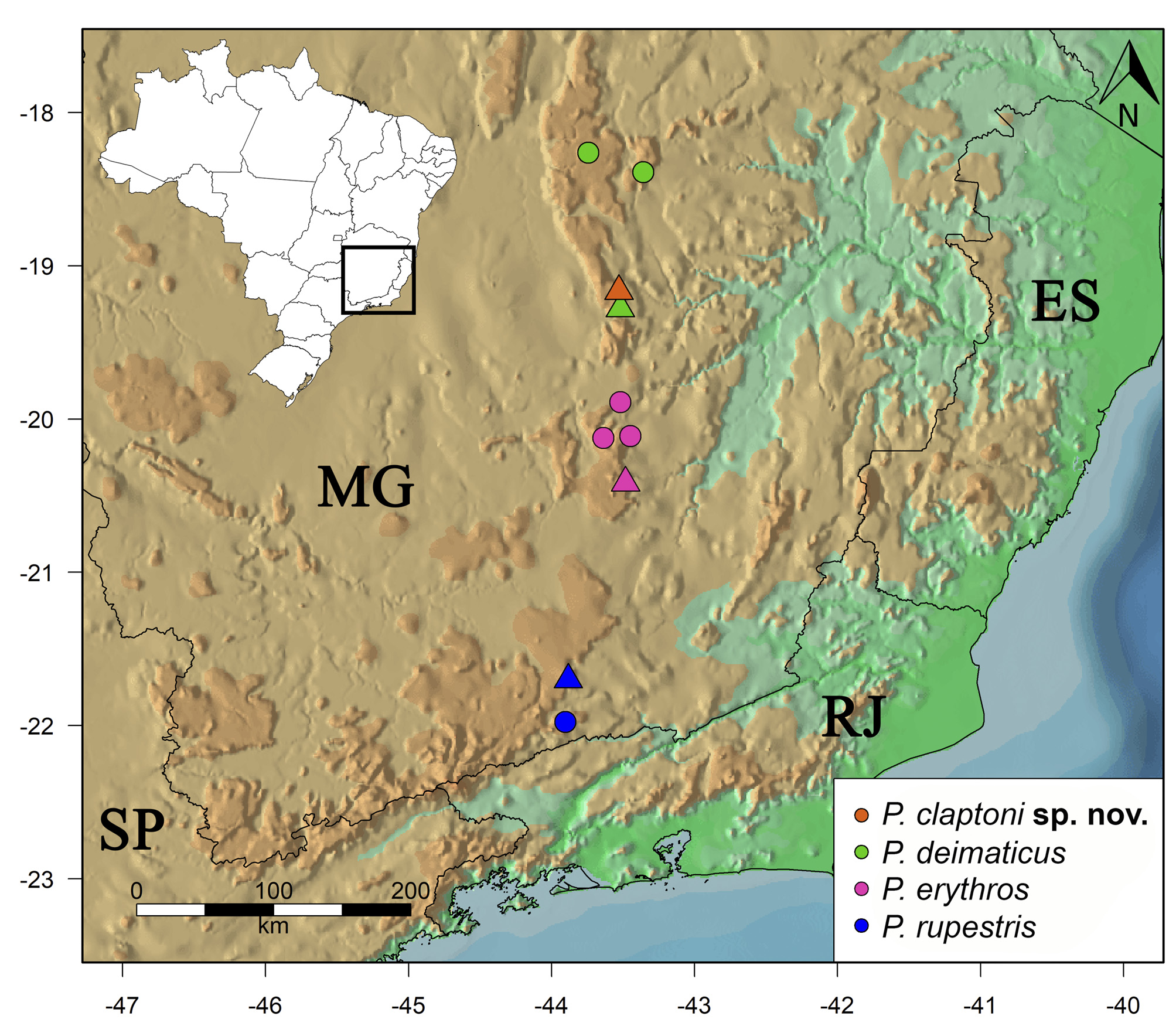
TABLE 1. Measurements (in mm) and proportions of the Phsalaemus claptoni sp. nov. type series and from P. rupestris type series and topotypes.
| Physalaemus claptoni sp. nov. | Physalaemus rupestris | |||
|---|---|---|---|---|
| Male (n=26) | Female (n=1) | Male (n=16) | Female (n=6) | |
| SVL | 16.2–18.2 (17.5 ± 0.5) | 18.2 | 16.4–18.4 (17.3 ± 0.6) | 17.4–18.9 (18.1 ± 0.6) |
| HL | 5.4–6.3 (6.0 ± 0.2) | 5.9 | 5.8–6.5 (6.1 ± 0.2) | 6.1–6.4 (6.2 ± 0.1) |
| HW | 5.3–6.0 (5.7 ± 0.2) | 5.6 | 5.6–6.5 (6.0 ± 0.3) | 5.8–6.1 (5.9 ± 0.1) |
| ED | 1.6–1.7 (1.6 ± 0.02) | 1.6 | 1.6–1.8 (1.7 ± 0.1) | 1.7–1.9 (1.8 ± 0.1) |
| ESD | 2.3–2.6 (2.5 ± 0.1) | 2.5 | 2.4–2.7 (2.6 ± 0.1) | 2.6–2.8 (2.7 ± 0.1) |
| END | 1.4–1.5 (1.4 ± 0.04) | 1.4 | 1.2–1.4 (1.3 ± 0.1) | 1.4–1.5 (1.5 ± 0.05) |
| AMD | 3.1–3.3 (3.2 ± 0.1) | 3.1 | 3.0–3.3 (3.1 ± 0.1) | 3.2–3.4 (3.3 ± 0.1) |
| IND | 1.4–1.5 (1.4 ± 0.04) | 1.4 | 1.6–1.7 (1.6 ± 0.04) | 1.6–1.8 (1.7 ± 0.1) |
| THL | 7.2–7.8 (7.6 ± 0.2) | 7.8 | 6.9–7.7 (7.2 ± 0.2) | 7.5–7.9 (7.7 ± 0.2) |
| TL | 7.0–7.6 (7.3 ± 0.2) | 7.5 | 6.5–7.1 (6.8 ± 0.2) | 7.0–7.3 (7.2 ± 0.2) |
| FL | 10.9–12.2 (11.6 ± 0.5) | 12.2 | 10.4–11.7 (11.1 ± 0.3) | 11.4–11.8 (11.7 ± 0.2) |
| HAL | 3.8–4.4 (4.1 ± 0.1) | 4.4 | 3.9–4.5 (4.2 ± 0.2) | 4.2–4.5 (4.3 ± 0.1) |
| HL/SVL | 0.33–0.35 (0.34 ± 0.01) | 0.32 | 0.34–0.38 (0.35 ± 0.01) | 0.33–0.36 (0.34 ± 0.01) |
| HW/SVL | 0.32–0.34 (0.32 ± 0.01) | 0.31 | 0.33–0.37 (0.35 ± 0.01) | 0.32–0.33 (0.33 ± 0.01) |
| HL/HW | 1.02–1.09 (1.05 ± 0.02) | 1.06 | 1.0–1.07 (1.02 ± 0.02) | 1.03–1.06 (1.04 ± 0.01) |
| THL/SLV | 0.41–0.45 (0.43 ± 0.01) | 0.43 | 0.4–0.43 (0.41 ± 0.01) | 0.42–0.44 (0.43 ± 0.01) |
| TIL/SLV | 0.4–0.43 (0.42 ± 0.01) | 0.41 | 0.38–0.41 (0.39 ± 0.01) | 0.38–0.41 (0.40 ± 0.01) |
| END/ED | 0.85–0.93 (0.89 ± 0.02) | 0.87 | 0.75–0.82 (0.79 ± 0.02) | 0.78–0.85 (0.81 ± 0.03) |
TABLE 2. Advertisement call parameters of Physalaemus claptoni sp. nov. and Pysalaemus rupestris, both recorded at their respective type localities.
| Parameters | P. claptoni sp. nov. n=5 males | P. rupestris n=3 males |
|---|---|---|
| Advertisement call - duration (s) | 0.80–1.28 (1.09 ± 0.11) n=45 | 1.51–2.20 (1.76 ± 0.18) n=44 |
| Advertisement call - number of pulses | 65–93 (78 ± 7) n=45 | 52–85 (61 ± 6) n=44 |
| A/B internotes interval (s) | 0.10–0.19 (0.13 ± 0.02) n=45 | 0.22–0.42 (0.28 ± 0.05) n=44 |
| Inter Advertisement call interval (s) | 0.14–1.38 (0.39 ± 0.27) n=41 | 0.26–1.75 (0.39 ± 0.3) n=24 |
| Advertisement call - note rate (notes/sec) | 1.56–2.51 (1.86 ± 0.21) n=45 | 0.91–1.32 (1.15 ± 0.11) n=44 |
| Note A - duration (s) | 0.11–0.16 (0.12 ± 0.01) n=154 | 0.05–0.10 (0.08 ± 0.01) n=93 |
| Note A - number of pulses | 12–17 (14 ± 1) n=154 | 8–11 (10 ± 1) n=93 |
| Note A - Pulse rate (pulses/sec) | 86.7–119.3 (105.8 ± 6.7) n=154 | 115.4–183.7 (133.7 ± 15.3) n=93 |
| Note A Rate (notes/sec) | 0.53–0.63 (0.59 ± 0.05) n=154 | 0.42–0.76 (0.58 ± 0.17) n=93 |
| Inter AA calls interval (s) | 0.70–1.90 (1.40 ± 0.30) n=113 | 0.22–5.27 (1.47 ± 1.30) n=64 |
| Note B - duration (s) | 0.67–1.01 (0.85 ± 0.09) n=45 | 1.14–1.78 (1.4 ± 0.17) n=44 |
| Note B - number of pulses | 53–81 (65.7 ± 7.4) n=45 | 41–75 (51.0 ± 6.4) n=44 |
| Note B - pulse rate (pulses/sec) | 67.5–92.1 (79.0 ± 6.5) n=45 | 31.4–43.1 (37.6 ± 2.8) n=44 |
| Note B - number of pulses group | 5–7 (6 ± 1) n=45 | __ |
| Note B Rate (notes/sec) | 0.32–0.61 (0.46 ± 0.2) n=45 | 0.20–0.34 (0.27 ± 0.07) n=44 |
| Note A - frequency bandwidth (Hz) | 1453.1–3140.6 (2247.0 ± 590.2) n=154 | 1875.0–3750.0 (2747.5 ± 743.3) n=93 |
| Note A - dominant frequency (Hz) | 1734.4–2765.6 (2131.7 ± 431.0) n=154 | 3046.9–3421.9 (3175.7 ± 130.9) n=93 |
| Note B - frequency bandwidth (Hz) | 1335.1–3375.0 (2435.3 ± 666.1) n=45 | 1875.0–3750.0 (2816.5 ± 793.2) n=44 |
| Note B - dominant frequency (Hz) | 1507.3–2859.4 (2505.5 ± 426.9) n=45 | 3046.9–3421.9 (3153.3 ± 115.7) n=44 |
| UFMG |
Universidade Federal de Minas Gerais |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |