
Polycarpa urmeli, Sanamyan, Karen & Hissmann, Karen, 2008
|
publication ID |
https://doi.org/ 10.5281/zenodo.181612 |
|
DOI |
https://doi.org/10.5281/zenodo.6234497 |
|
persistent identifier |
https://treatment.plazi.org/id/3143879F-FFE9-3116-FF7D-3412FF54B110 |
|
treatment provided by |
Plazi |
|
scientific name |
Polycarpa urmeli |
| status |
sp. nov. |
Polycarpa urmeli sp. nov.
( Figures 1 View FIGURE 1 , 2 View FIGURE 2 , 3 View FIGURE 3 )
Material examined. One specimen (holotype): JAG" dive # 660, 18 November 1999 277 m on rocky ground off the southeast coast of the volcanic island Sangihé, northeast Indonesia (03°22.08´N, 125°31.19´E). This specimen is deposited in the Kamchatka Branch of the Pacific Institute of Geography ( KBPIG), KBPIG 1/ 1356.
Second specimen: from JAGO dive # 659, 17 November 1999 217 m on rocky ground off the northeast coast of Sangihé Island (03°44.75´N, 125°25.19´E).
Further material: In addition to the collected samples, numerous observations of live specimens of this species were made during a series of submersible dives performed along the North Indonesian Sangihé Islands and the northern part of Sulawesi. A video camera mounted on the submersible´s manipulator arm provided detailed in situ video close-ups of the animals.
The specimens were collected alive with the manipulator arm of the submersible and stored in a sampling tube for the rest of the dive. At the surface, they were transferred to a glass aquarium with fresh sea water for initial examination and measurement. They were then narcotized with magnesium-chloride and preserved in 3% formalin mixed with sea water. Only one specimen (the holotype) has been examined anatomically, the second specimen kept intact.
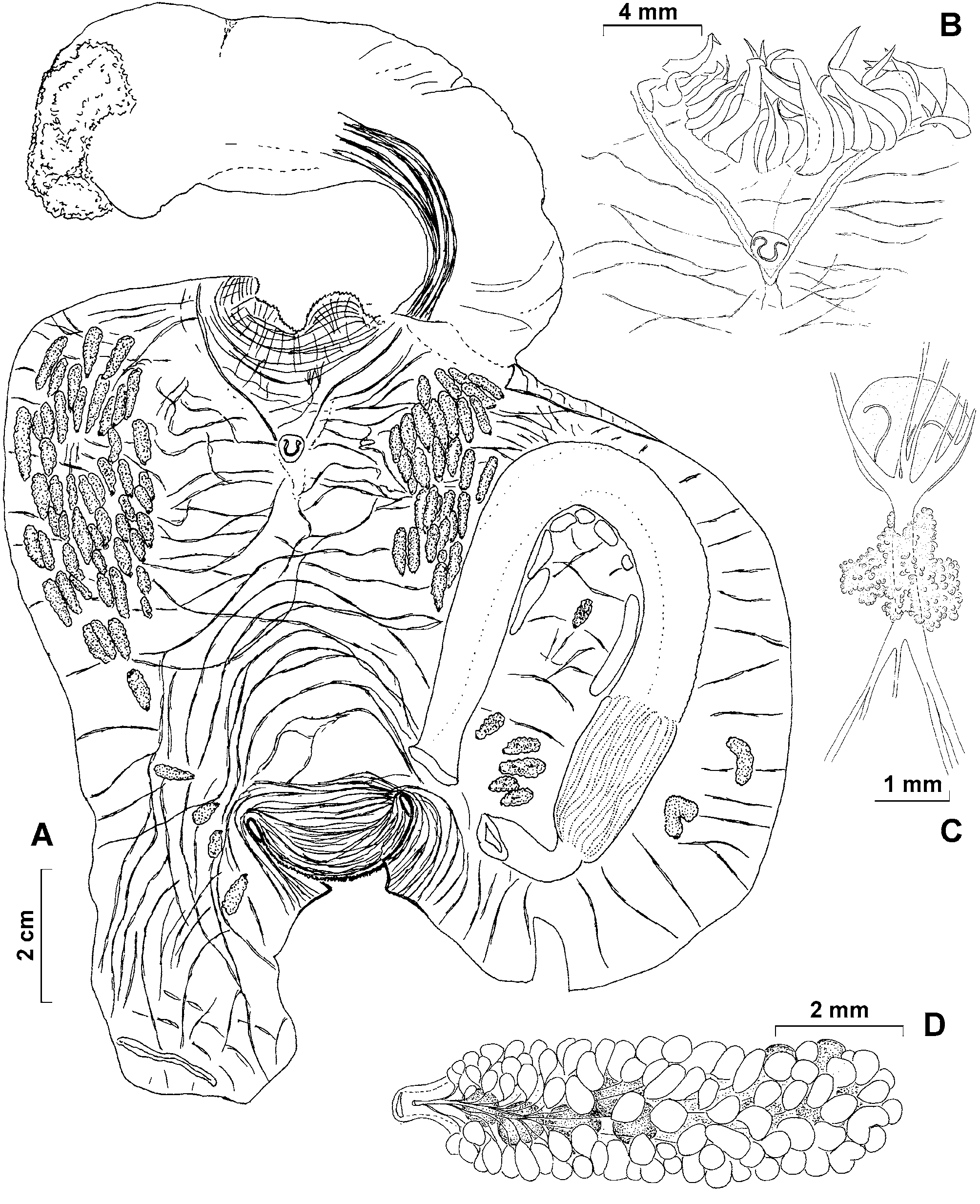
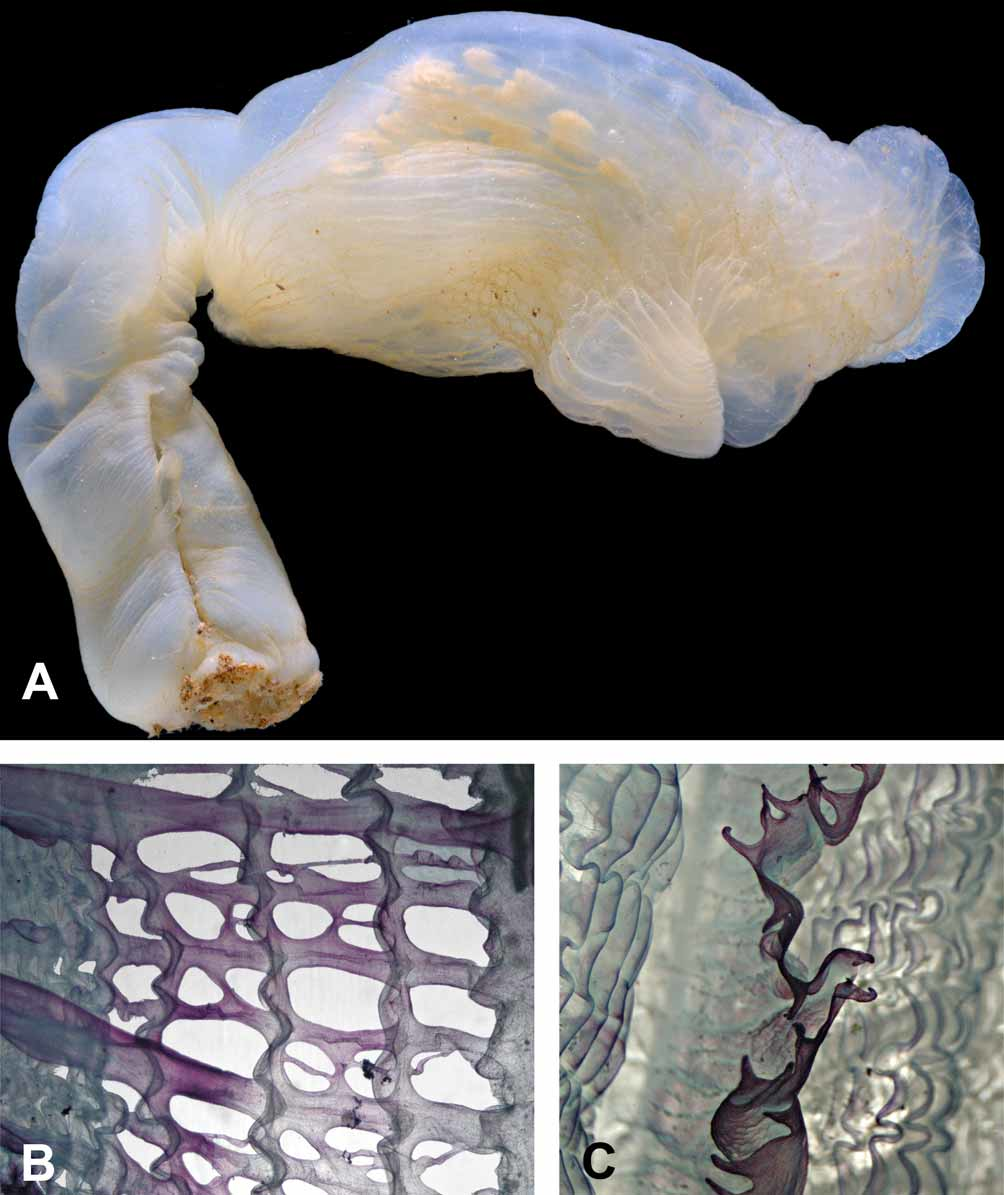
Description. Living specimens have an elongate and almost cylindrical body and are supported on a fleshy stalk arising from the antero-ventral part of the body ( Fig. 1 View FIGURE 1 A). Body and stalk are of about the same length. The body of the living holotype is 12 cm long and the stalk 14 cm. In preservative the body is about 10 cm long and 5 cm wide and the stalk 11.5 cm long and 2 cm diameter at the upper end just below the body, increasing to about 3 cm at the base. The stalk is attached to the substrate by a flat attachment area. In the living specimens, the body is orientated horizontally on the stalk with the dorsum uppermost. In preserved specimens, the stalk is bent dorsally (probably by contraction of its thick dorsal muscles) ( Fig. 2 View FIGURE 2 A). The tunic is thick, soft and transparent with a yellowish tint and a glassy transparent inner layer. The surface of the tunic is smooth ( Fig. 1 View FIGURE 1 B), but has shallow wrinkles when contracted ( Fig. 3 View FIGURE 3 A). Sparse minute elongate structures embedded in the superficial layer of the tunic are visible to the naked eye as black dots ( Fig. 1 View FIGURE 1 D). These are assumed to be epibionts (see below). Otherwise the surface of the tunic is naked, lacking either attached particles or hairs or other outgrowths.
The free end of the body (its posterior end) is rounded. It has a well defined thick solid transverse posteroventral crest (resembling those of some species of Culeolus ), that is visible on the photos of the living specimens and is conspicuous on the preserved specimen. The circular, branchial aperture is at the anterior end of the body close to the top of the stalk. Its margin is minutely dentate but it is not lobed ( Fig. 2 View FIGURE 2 A). The large transverse atrial aperture is halfway along the dorsal side of the body. It exposes a large part of the branchial sac when widely open in life ( Fig. 1 View FIGURE 1 C). It is bilabiate with an anterior and a posterior lip ( Fig. 3 View FIGURE 3 A).
The body wall is thin and transparent and an extension of it filled by loose parenchymatous tissue (part of the haemocoele) projects down to the base of the stalk. The body muscles are composed of thick, primarily transverse bands forming a loose network. Some of the muscles that encircle the anterior end of the body continue into the stalk, where they form a wide ribbon running along its dorsal side and ending half way to its base. Circular muscles are crowded around the short siphons at both openings of the body. The holotype has about 40 large, laterally flattened, tapering tentacles of two different sizes arising just beyond the base of the thin branchial velum ( Figs. 1 View FIGURE 1 E, 2B). The prepharyngeal band, consisting of two thick lamellae close to the ring of tentacles and the branchial sac, forms a dorsal V around the dorsal tubercle ( Figs. 2 View FIGURE 2 A, B). The dorsal tubercle has a U-shaped opening with two re-curved horns. The elongate ganglion is only slightly thicker than the pairs of anterior and posterior nerves that extend from it. The ganglion together with the neural gland is just posterior to the dorsal tubercle ( Fig. 2 View FIGURE 2 B). The dorsal lamina is a high membrane that has numerous long languets on its margin ( Fig. 3 View FIGURE 3 C). The four high branchial folds on each side, have numerous internal longitudinal vessels crowded on them. The branchial formula of the right side of the branchial sac is E4(12)6(12)5(13)6(16)DL. The perforations of the branchial wall are rather irregular, oval or rectangular and usually slightly elongate transversely ( Fig. 3 View FIGURE 3 B). Ciliated epithelium has not been detected lining these perforations.
The simple wide gut loop occupies much of the left side of the body ( Fig. 2 View FIGURE 2 A) and is clearly visible through the transparent tunic in the living specimens. The oesophagus is of moderate length and bent at a right angle before it opens into the stomach. The elongate stomach occupies less than half of the ascending limb of the gut loop, is slightly wider than the intestine and has numerous internal longitudinal folds. The intestine is isodiametric along the whole length, terminating in an anus with a smooth border. The holotype emptied its intestine when it was transferred into the aquarium and now contains neither mud nor other particles.
Gonads are numerous, about 60 on the right and 40 on the left, and are firmly attached to the body wall but not embedded in it. Most are in the anterior halves of each side of the body, where they form rather compact clusters, and often overlap each other ( Fig. 2 View FIGURE 2 A). On the left, six gonads are inside the primary gut loop and two are between the gut loop and the mid-ventral line of the body. Each gonad consists of a central ovary and numerous (sometimes more than 100) male follicles spread over the sides and mesial surface, but not beneath the ovary. Male ducts curve around each side of the ovary, run along its mesial surface and occasionally join together to meet the vas deferens of variable length ( Fig. 2 View FIGURE 2 D) that opens at the base of the short oviduct. The gonads are aligned towards the atrial aperture.
Several low fleshy endocarps are attached to the body wall along the intestine inside the gut loop ( Fig. 2 View FIGURE 2 A). Endocarps are not present outside the gut loop.
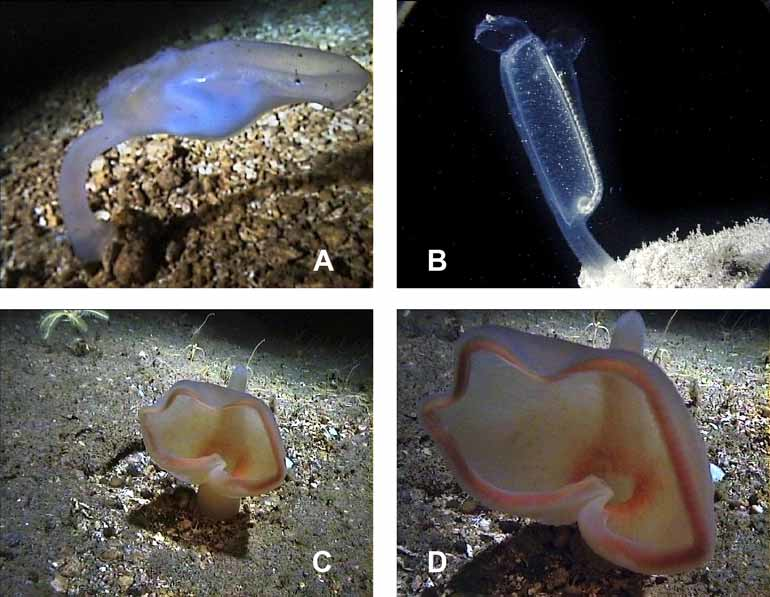
Observations on the living animals. Habitat and abundance: Specimens of Polycarpa urmeli sp. nov. were sighted during submersible dives to between 50 and 400 m water depth around north Indonesia in November 1999. They occurred only sporadically and scattered over large areas. Only one aggregation of 10 individuals, separated by several meters from each other, was found at 237 m depth off Bunaken island. Specimens were present off the islands Sangihé, Mahengetang, Siau, Manado Tua, Bunaken and off the north coast of the northern arm of Sulawesi at water depths between 217 and 277 m and in water temperatures ranging from 12–16°C. They were not found in the Gulf of Tomini along the south coast of the northern arm of Sulawesi. Exploratory dives to determine the upper and lower depth limits of the species were not carried out. Most animals were sighted between 220 and 230 m. They were seen to have settled on open slopes mainly with rocky substrate, and were exposed to bottom currents that could reach 1–1.5 m/s, sometimes as strong surges that swayed the animals back and forth. Polycarpa urmeli lives in an environment with constant or frequent bottom currents and seems to be well adapted to these conditions. When the large branchial aperture is facing towards the current and both apertures are open wide, a large amount of water can pass through the animal without needing cilia to create a flow. This implies that species like P. u r m e l i with non-ciliated stigmata and large branchial perforations only occur in areas of continuous and strong bottom currents. Many deepwater species lack cilia and many of these are large, solitary and typically possess a peduncle arising close to the large branchial opening. The lack of food in their deep water habitats appears to be compensated for by the large amount of water passing through the animal to maximize the volume of microparticles filtered from it by the fine mucus net on the branchial sac. Monniot and Monniot (1978, p.200) suppose that "the energy required for this form of nutrition would be minimal".
Behaviour: Polycarpa urmeli sp. nov. is highly flexible on its fleshy stalk, which lifts the animal about 10 cm above the sediment and into the current. All animals observed had their large branchial apertures facing the current. Strong surging currents that transport a lot of sediment particles, sometimes caused the animals to close both apertures by contraction, and to lower the free (posterior) end of the body towards the substrate. However, this posture was also sometimes observed when there was no current. Individuals also responded to direct tactile contact by contraction of the apertures.
Commensals: A diversity of commensals were observed on the outside of the tunic and inside the peribranchial cavity of several P. u r m e l i individuals. Small specimens of sea cucumbers (Holothuroidea), brittle- (Ophiuroidea) and feather-stars ( Crinoidea ), topshells (Trochidea) and spider crabs (Maiidae) moved around on the ascidians and their stalks ( Fig. 1 View FIGURE 1 H). The filter-feeders among these animals used their host to gain elevation for better exposure to the current, others probably fed on detritus particles and micro-organisms that occurred on the tunic. A tiny benthic ctenophore with long extended tentacles (Order Platyctenidae , probably Lyroctenidae ) was also found on one P. u r m e l i specimen. Another P. u r m e l i carried piggyback between the two body openings a transparent sessile ascidian with cylindrical siphons ( Fig. 1 View FIGURE 1 F). Several P. u r m e l i individuals contained a single or a pair of shrimps. The small shrimps moved around on the outside of the branchial sac, climbed in and out of the peribranchial cavity at the artial aperture, and seemed to be well adapted to their host. One P. u r m e l i specimen contained a small eel (most likely a young conger eel) that probably got trapped in the peribranchial cavity by accident ( Fig. 1 View FIGURE 1 G). The holotype contained in its peribranchial cavity a symbiotic nemerthean worm which appeared to be an undescribed species belonging to the genus Gononemertes (pers. comm. Alexei Chernyshev, Valdivostok). Two Gononemertes species are known from ascidians of the genus Pyura and Phallusia (see Dalby 1995). The minute hairs visible on the tunic of some living specimens of P. u r m e l i ( Fig. 1 View FIGURE 1 E) are assumed to be epibionts. Microscopic examination of the tunic revealed these to be short cylindrical bodies completely embedded in the peripheral layer of the tunic. They appear to contain tentacle-like structures which in life could be expanded to the exterior giving an impression of hairs.
Remarks. Several features of this Polycarpa species, including the absence of the ciliated stigmata and the toothed dorsal lamina, have not been reported for Polycarpa before. The structure of the pharyngeal wall is intermediate between the branchial wall of typical Styelidae and the deep-water genus Bathyoncus . However, the type species of the genus Bathyoncus , B. mirabilis Herdman, 1882 , has only one voluminous gonad on each side of the body and may have affinities with the styelid genus Cnemidocarpa . Nevertheless its branchial wall, like that of Culeolus , is lost, leaving large rectangular meshes formed by longitudinal branchial vessels crossing transverse sinuses. In the present Polycarpa species the branchial wall persists and is perforated by irregular unciliated perforations probably derived from true stigmata by losing cilia and uniting adjacent stigmata into larger perforations. This may be an adaptation to deep water conditions and feeding by 'passive' filtration (see below). Such unciliated perforations are known in Pterygascidia mirabilis Sluiter, 1904 (see Kott in press) and in several other deep-water species belonging to genera in which most other species have typical ciliated stigmata (e.g. Distomus pacificus Monniot and Monniot, 1991 , Monandrocarpa abyssa Sanamyan and Sanamyan, 1999 ). Bathyoncus arafurensis Monniot and Monniot, 2003 , has a denser branchial sac and, unlike B. mirabilis , numerous polycarps and its assignment to the genus Bathyoncus is not well established.
The present species is not closely related to any of the more than 120 Polycarpa species. The preserved specimens resemble superficially P. c l a v a t a Hartmeyer, 1919 (especially as figured by Kott 1985, Fig. 71a), which also has a thick stalk inserted to the anterior end of the body. However, endocarps of P. clavata are spread over the whole body wall, and gonads and the shape of the gut loop differ significantly from those of P. urmeli . Living specimens of P. clavata are brillant orange to yellow in colour and the tunic of the species is thick and quite firm.
Sympatric species. Four other stalked solitary ascidians were observed during the JAGO dives in northern Indonesia. The first was a transparent species of similar size and body shape like Polycarpa urmeli , except the free (posterior) end of its body was slightly flattened and rectangular and the atrial aperture was not in the middle of the body but slightly closer to the branchial aperture ( Fig. 4 View FIGURE 4 A). This transparent species occurred in the same habitat as P. u r m e l i. The second ascidian species appeared to be Pterygascidia mirabilis Sluiter, 1904 (family Ciallusiidae , see Kott in press) and was encountered at 253 m at the north coast of Sulawesi ( Fig. 4 View FIGURE 4 B). The branchial and atrial apertures of this species were both close together at the free end of the transparent elongated body. The siphon of the dorsal branchial aperture had the shape of a hood. The opening of the hood was facing the current that flows from the stalk along the elongated body. The atrial aperture had a ventral lobe and was facing down-current. The genus Pterygascidia is monotypic and known from the Philippines, Indonesia and the northwestern Australian continental shelf (Kott in press). The third ascidian species belonged to the genus Culeolus , probably C. herdmani Sluiter, 1904 . Members of this genus have an extraordinarily long thin stalk, much longer than the body itself and a wide-meshed branchial sac which is reduced to the network of longitudinal and transverse vessels, the branchial wall having disappeared. The Culeolus specimen was encountered at 220 m depth at the Mahengetang submarine volcano. The fourth species belonged to the family Octacnemidae which is exclusively known from deeper waters and mainly characterized by the hypertrophy of the oral siphon. One specimen of this family was found at 270 m depth off Lembeh Island at the south coast of northern Sulawesi. The oral siphon of the pale yellowish species was greatly enlarged to form a large basket or hood ( Fig. 4 View FIGURE 4 C,D). The outer edge of this basket was undulated and rose-colored allaround like roughed lips. Also the inside of the basket around the opening to the branchial sac was rose-colored. An elongated prominent atrial siphon pointing vertically upwards opened at the antero-dorsal part of the body. The short thick stalk extended at the antero-ventral corner. The animal oriented the aboral surface of the oral basket to the on-coming bottom current, i.e. the oral opening of the body was facing down-current, as also observed for Megalodicopia hians Oka 1918 ( Okuyama et al. 2002; pers. comm. Edward Seidel, Monterey Bay Aquarium). Members of this family, like Megalodicopia hians , are macrophagous and engulf large particles and organisms such as small invertebrates, whereas others, like the two genera Dicopia and Situla , have probably a mixed diet and feed on both large and tiny particles ( Monniot & Monniot 1975).
Ethymology. The species name refers to the shape of the main body that resembles the head of a puppet dinosaur called “Urmel”, the main character of a famous puppet show broadcasted in the German Television in 1969.

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.