
Lynceus tatei ( Brady, 1886 )
|
publication ID |
https://doi.org/10.11646/zootaxa.3702.6.1 |
|
publication LSID |
lsid:zoobank.org:pub:A875F2FF-3DAA-4AC3-9451-773F095A7C82 |
|
DOI |
https://doi.org/10.5281/zenodo.5463045 |
|
persistent identifier |
https://treatment.plazi.org/id/346D87FD-F139-255B-2A90-498EFEFE876C |
|
treatment provided by |
Felipe |
|
scientific name |
Lynceus tatei ( Brady, 1886 ) |
| status |
|
Lynceus tatei ( Brady, 1886) View in CoL
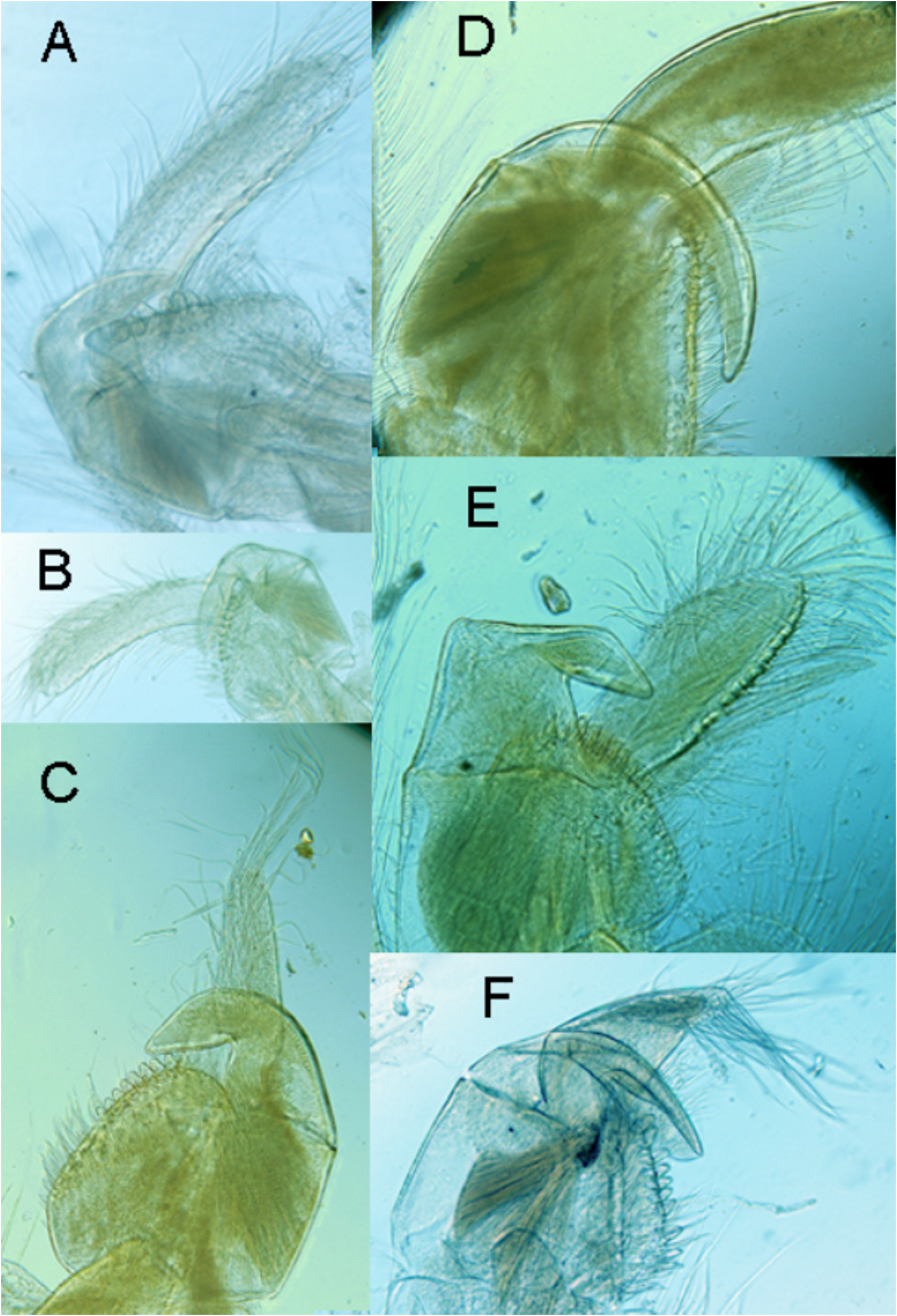
( Figs. 3A,B View FIGURE 3 , 4B View FIGURE 4 , 5 View FIGURE 5 , 6C,D View FIGURE 6 )
Limnetis tatei Brady, 1886: 84–85 View in CoL , fig. A; Sars 1895: 43–46, pl. 8; Henry 1924: 121–122 (list), 135–136 (text).
Lynceus tatei Sayce, 1903: 258–259 View in CoL , pl. 36, fig. E, 1–3; Wolf 1911: 255 (list); Dakin 1914: 295 (list), 303 (text); Daday 1927: 6 (key), 45–49, fig. 155; Glauert 1924: 59–60; Martin & Belk 1988: 452 (list); Brtek 1997: 61 (list); Richter & Timms 2005, 342.
Limnetis eremia Spencer & Hall, 1896: 244 View in CoL , pl. 23, figs 30–32.
Lynceus eremia Sayce, 1903: 258–259 View in CoL , pl. 36, fig. F, 1–3; Wolf 1911: 255 (list); Dakin 1914: 295 (list); Chilton 1917: 481.
Comments. Brady’s (1886) original “description and figures are....very imperfect, and hardly suffice to fully recognize the species” ( Sars 1895). However Sars’ (1895) own redescription is adequate, but the specimens came from 1030 km away from the type locality at no specific site in the Sydney area and they are not available now. The original author did not designate any types and no specimens survive from the type locality. The original type locality was drained decades ago, but fortunately there is a collection in the SAM (C3652) about 50 km away from a temporary swamp which still exists. A neotype has been chosen from this site.
New Type locality. Australia, South Australia, 29.5 km SE of Millicent , pond, 37 o 44’ 44”S, 140 o 37’ 47”E GoogleMaps .
Neotype. Male deposited in the South Australian Museum. Length 3.8 mm, height 3.2 mm Registration number SAM 7641 View Materials .
Other material from new type locality: 3 males, 3 females from South Australia, 29.5 km SE of Millicent , pond, 37 o 44’ 44”S, 140 o 37’ 47”E, 5 September 1971, J. Aslin, SAM C3652 View Materials GoogleMaps .
Other material. New South Wales: 125 km NW of Bourke, Bloodwood Station, pool near Plover Pan , 29 o 31’ 06.7”S, 144 o 49’ 34.8”E, 31 May 1999, S. Richter. AM GoogleMaps P.90482; 125 km W of Bourke, Bloodwood Station, Last Pan , 29 o 29’ 08.3”S, 144 o 49’ 02.7”E, 31 May 1999, S. Richter, AM GoogleMaps P.90483; 12 km NNW of Berridale; pond near road on ‘ Stoneleigh ,’ 36 o 15’ 41.5”S, 148 o 47’ 42.1”E, 1 February 1996, BVT, AM P47130 & P88146 GoogleMaps ; 42 km SSW of Cooma, Bungarby, swamp near Barkers Lake , 36 o 41’ 32”S, 149 o 00’ 14”E, 27 November 1991, BVT, AM P47131 & P88147 GoogleMaps .
South Australia: Gawler District, near Roseworthy Lagoon , 10 October 1991, R . G. Briggs, SAM C7645 View Materials ; 3.2 km S of Salisbury, vernal pools at Parafield Airport , 34 o 48’ 8.3”S, 138 o 37’ 35.2”E, 8 October 2010, P. Coleman, SAM C7640 View Materials GoogleMaps .
Tasmania: Rosstown , 15 September 1900, O.A.Sayce, NMV J54021 View Materials , J62987 View Materials , J62987.1 About NMV .
Victoria: Mansfield , 23 October 1910, no collector recorded, NMV J53993 View Materials , J62982 View Materials , J62982.1 About NMV ; Cheltenham , 22 October 1910, no collector recorded, NVM J46629 View Materials , J62980 View Materials , J62980.1.
Western Australia: 50 km ESE of Perenjori, large central pool on Wanarra Rock , 29 o 31’ 23.5”S, 116 o 47’ 38.1”E, 20 August 2010, BVT, WAM 51608 GoogleMaps ; Bullsbrook, Warbrook Rd, Tortoise Reserve Area , 1963, Zoology Honours Class, WAM , 51313; Bullsbrook, 21 mile peg, pond in Tortoise Reserve Area , 27 September 1963, Zoology Honours Class, WAM 51314 ; Gosnells, a pond, 5 September , 1960, D.H. Edwards, AM P55619 & P88149 ; Oakford, Orton Rd , 24 July 1961, A. R . Main , WAM 51324 ; Serpentine, Halls Rd 0.5 km N of railway level crossing, 32 o 21’ 28”E, 115 o 58’27”E, 15 September 2011, BVT, WAM 51609 ; 22 km SE of Kojonup, Ngopitchup Swamp , 19 October 2003, MZ, WAM 51610 ; Bremer Bay District , 15.5 km SE of South Coast Highway-Bremer Bay Rd junction, 34 o 28’ 11”S, 118 o 39’ 20”E, 4 September 2011, BVT, WAM 51611 GoogleMaps ; 11.3 km NW of Augusta, roadside pool 1.8 km from junction of Caves Rd and Hamelin Bay Rd, 34 o 13’ 11”S, 115 o 04’ 20”E, 6 July 1990, IAEB, WAM 51612 GoogleMaps .
Diagnosis. Adult size < 4 mm. Carapace broadly arcuate anteriorly, narrower posteriorly. Head axis curved through 90 o. Male with endite VI of thoracopod 1 digitiform and covering almost all the spines on the mediolateral corner of the endite III and reaching about half way along its medial surface. Female almost invariably with notch anteriolaterally on rostrum.
Description of Neotype. Male. Head ( Figs.5B,C View FIGURE 5 ) large, subequal to trunk, surface finely punctuate. Fornices broad, angulated and arcuate over second antennal base, and dividing head into a larger upper portion and a smaller lower part. Small mound centroposteriorly, the site of the dorsal organ. Compound eyes close together about midway along the central ridge between the cervical suture and rostrum. A small frontal pore and two small lateral setal fields just anterior to the eyes, and an ocellus deeply embedded beneath the setal fields. Posterior and anterior surfaces of head at similar levels on either side of lateral suture between compound eye and lateral fornix. Rostrum longer than wide with truncated apex about one-third wider than the narrowest part and terminating each side in a minor extended denticular apices. Terminal facet almost triangular and at right angles to head axis and with a citiated arcuate ventral margin. Dorsal carina sharp and convexly curved so that the plane of the compound eyes and setal fields at right angles to the terminal facet of the rostrum.
First antenna ( Fig. 5D View FIGURE 5 ) small, almost reaching to the rostrum apex. Two antennomeres, proximal cylindrical and distal semiclavate, both of equal length. Distal antennomere bearing numerous papillae, mainly dorsally. Each papillae with a delicate digital process.
Second antenna ( Fig. 5E View FIGURE 5 ) biramous well developed, protruding beyond the rostrum by about half its length. Peduncle of three segments, the first with two to three plumose setae, the second with one to two spines, and the distal segment with about five spines attending base of the anterior (dorsal) ramus. Both antennal rami with about 18 antennomeres, almost all with long ventral plumose setae. Anterior ramus with short dorsal setae one per antennomere and ventral ramus with two spaced dorsal setae towards the apex.
Labrum large, linguiform, clothed in small setae particularly apically. Mandible broadly spatulate. First maxilla typical for genus and second maxilla absent.
Carapace: ( Fig. 5A View FIGURE 5 , 6C View FIGURE 6 ) Broadly circular in outline, but in detail, anterior higher than posterior with dorsoposterior and ventroposterior margins sloping steeper than the smooth arcuate curve anteriorly. Hinge line dorsal but sloping anterior to posterior and only just hidden by valves. Anteriorly a slight embayment in the carapace at the position of the compound eye so that the apex of the head visible when valves closed. Valves roundly inflated laterally. Carapace surface finely punctuate. Abductor muscle scar in an anteriolateral position about twice its diameter from the margin.
Thorax: Ten thoracic segments each with a pair of thoracopods. The telsonic posterior plate partly divided centroposteriorly, each half bearing a long seta apically. Somite below enlarged, similarly divided and bearing small denticle apically.
Thoracopod I ( Figs. 3A, B View FIGURE 3 , 5F View FIGURE 5 ) modified as a clasper appendage, right and left claspers equal in size and shape. Endite VI arcuate, but more convex about 2/5ths along its length, and divided into a basal part about as wide as high and a longer digitiform process almost reaching to end of the spine row on distomedial corner of endite III and about half way along medial edge of this endite. Digitiform process part of endite VI tapering but slightly towards its blunt apex and almost twice as long than the width of the basal part where the two meet. Endite V cylindrical shaped, curved and four to five times longer than wide, and narrowing apically.Two areas of setae, one a row of about 12 stout setae extending in an imperfect row about 3/5ths of the distal length of the endite, and the other oval field of dishevelled thinner setae centred about 60 o around from the row but reaching near to the row. Endite IV small, about one seventh the length of endite V with a straight, almost setaeless, margin against endite V and a convex opposite margin with numerous long setae. Endite III vaguely rectangular but wider than long, and with medial margin narrowing markedly distally, so that distal margin about half width of basal margin. A row of about 8 stout spines medodistally, each with rounded apices. A mediolateral field of stout setae, in an indistinct row of shorter setae on the inner surface of the mediolateral field and a field of longer setae marginally and many longer setae clumped distally.
Thoracopod II typical for genus (Martin & Belk 1988).
Thoracopods III to VII similar, though decreasing in size posteriorly.
Thoracopods VIII to X reduced and lacking the epipodite and proximal lobe of the exopodite.
Size. Neotype length 3.8 mm, height 3.2 mm. The other specimens in the collection are slightly bigger: males 4.1 mm long, 3.9 mm high, 4.2 by 3.8, and 4.4 by 3.8; females 3.9 by 3.4, 4.0 by 3.5 and 4.0 by 3.7. Brady’s specimens were slightly bigger, reaching 0.5 mm.
Comments on female. Sars (1895) describes the female in detail and then briefly comments on the male. From this and his figures in plate 8 there is little doubt that the present neotype and the specimens Sars had are of the same species. This is despite the huge distance apart of the original type locality and the site from which Sars’ specimens came and the fact that perhaps L. tatei was never common in the Sydney area. There are no old collections in museums and no modern collections anywhere. Nevertheless the original description, Sars redescription and the description of the neotype all note the unusual carapace shape of a broad anterior and narrower posterior half, the notches on the anteriolateral corners of the female rostrum, and the small size (< 4mm). In addition Sars and the description of the neotype record the cilia of the anterior margin of the rostrum, the bladelike dorsal carina of the rostrum and the unique structure of endite VI of the first thoracopod.
Sars (1895) notes the head shape with its notch on each anteriolateral corner of the rostrum. This rostrum in females from the new type locality is similarly notched and the rostrum is twice as long as wide and it wider at its apex by about 30% from its narrowest part ( Fig. 4G View FIGURE 4 ). The apex is arcuate. The first and second antenna are similar to the male’s (though there are a few dorsal setae on the ventral ramus rather than two), and the mouthparts are standard for Lynceus . As noted by Sars for his material, these females have 12 trunk thoracomeres each with a pair of thoracopods of typical Lynceus structure. The telsonic segment is typical for Lynceus with a divided plate dorsally and each half with a terminal seta. Beneath the telson the last somite is also divided, each half terminating in a denticle. The lamina abdominalis ( Fig. 5J View FIGURE 5 , 6D View FIGURE 6 ) consistes of 3 digitiform lobes anteriorly and laterally and a posterior lobe with another digitiform lobe dorsally. The posterior lobe is probably folded underneath the lamina abdominalis in life ( Fig. 5D View FIGURE 5 ); certainly this would make it more efficient in holding eggs.
While there is no possibility the synonymised species L. eremia Spencer & Hall 1896 is the entity Sars was dealing with, it is just possible that populations of what was once thought to be this separate species L. eremia have been re-encountered and described as one of the new species below. Based on habitat this is ecologically possible, as L.eremia occurred in rock holes in northern South Australia, precisely within the distribution and habitat preference of L. magdaleanae n. sp. However L. eremia’s distinctive carapace shape, notched female rostrum and small size also distinguishes it from this species, as well as promote its synonymy with L. tatei
Resting egg ( Fig. 3B View FIGURE 3 ) is round with an irregular low wide ridges enclosing enlongated irregular depressions. The size 149 ± 9.2 ųm (n = 5).
Variability. Only a few populations have the slight indentation on the anterior margin of the valve, so that the compound eye is readily visible when the valves are closed. Setae on the third peduncle of the second antennae may number as many as seven, instead of the usual five, though the number of antennomeres in each ramus hardly varies around 18. In males the digitifrom process of the endite VI varies in length from being about ¾ covering the mediodistal spines of endite III, to be just longer. Setae on the mediolateral field of endite III vary in number and size in both sexes; similarly for the setae on the palps of thoracopod I in males.The mediolateral spines of endite III vary in number from five to nine.The processes extending from the lamina abdominalis of females vary in proportions between some populations so its structure, except for the single dorsal process, is an unreliable diagnostic character.
Distribution. L. tatei occurs in southern Australia from about Geraldton to Sydney. There are also old records (>100 years old, but no specimens in museums for confirmation) and one by Zofkova (2006) from central Australia (again no specimens available to confirm identification). These may be due to misidentifications or recent loss of habitat, as it is apparently not in the remote inland now, except in the Paroo ( Richter and Timms, 2005). The author and Magdaleana Zofkova (pers. comm. 2006) have only ever found it in temporary ponds. Many early collections were from the vicinity of Sydney ( Sars 1895 examined a collection sent by Mr Whitelegge from the Port Jackson area, but this collection does not exist now) and Melbourne, but all these sites have been lost by urbanisation. Presently its apparent stronghold is in the southwest corner of Western Australia and to a lesser degree in the Monaro in NSW.



No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Lynceus tatei ( Brady, 1886 )
| TIMMS, BRIAN V 2013 |
Lynceus tatei
| Brtek, J. 1997: 61 |
| Daday 1927: 6 |
| Dakin, W. J. 1914: 295 |
| Wolf, E. 1911: 255 |
| Sayce, P. A. 1903: 259 |
Lynceus eremia
| Chilton, C. 1917: 481 |
| Dakin, W. J. 1914: 295 |
| Wolf, E. 1911: 255 |
| Sayce, P. A. 1903: 259 |
Limnetis tatei
| Henry, M. 1924: 121 |
| Sars, G. O. 1895: 43 |
| Brady, G. S. 1886: 85 |