
Proa valdearinnoensis, Mcdonald, Andrew T., Espílez, Eduardo, Mampel, Luis, Kirkland, James I. & Alcalá, Luis, 2012
|
publication ID |
https://doi.org/10.11646/zootaxa.3595.1.3 |
|
publication LSID |
lsid:zoobank.org:pub:B5D4F5C3-BD6C-4ECE-BA28-88D1AD587207 |
|
persistent identifier |
https://treatment.plazi.org/id/366B87BB-FFB7-FFB2-CDC4-FACCFAAEF841 |
|
treatment provided by |
Felipe |
|
scientific name |
Proa valdearinnoensis |
| status |
gen. et sp. nov. |
Proa valdearinnoensis gen. et sp. nov.
Etymology. The generic name is the Spanish word for prow (‘ proa ’), in reference to the pointed shape of the predentary. The specific name is in reference to Val de Ariño, the traditional name of the coal mining area around the municipality of Ariño, near which the fossils were discovered, with the Latin ending - ensis (‘from’).
Holotype. AR-1/19 (deposited at MAP), a partial skeleton consisting of a disarticulated but associated skull including the premaxillae, partial maxillae, quadrates, supraorbitals, articulated braincase and skull roof, predentary, left dentary, and partial right surangular (AR-1-2012); right dentary (AR-1-2013); an isolated tooth (AR-1-2014); and several unprepared postcranial bones.
Paratype. AR-1/48 (deposited at MAP), a disarticulated but associated skull (AR-1-1364, AR-1-1367, AR-1- 1373–AR-1-1376, AR-1-1380, AR-1-1382, AR-1-1384–AR-1-1386, AR-1-1388, AR-1-1395), right dentary (AR- 1-1365, AR-1-1366), a left dentary (AR-1-1383), and six isolated teeth (AR-1-1369–AR-1-1372, AR-1-1378, AR- 1-1379) .
Referred Material. AR-1/57, a disarticulated but associated skull and dentary; AR-1/58 and AR-1/70, two partial skeletons. All referred material is deposited at MAP. Much of this material is still being prepared.
Locality and Horizon. Site AR-1 (bone concentrations AR-1/19, AR-1/48, AR-1/57, AR-1/58, AR-1/70), Mina Santa María, Ariño, Teruel Province, Spain. Middle Interval with Coal, Lower Sedimentary Succession, Escucha Formation ( Rodríguez-López et al. 2009), lower Albian ( Alcalá et al. 2012; Tibert et al. in prep.).
Diagnosis of genus and species by monotypy. Diagnosed by a single autapomorphy: predentary comes to a point at its rostral margin, with divergent lateral processes. Also diagnosed by the following unique combination of characters: dentary tooth row is convex dorsally in lateral view (otherwise known in only Owenodon hoggii [ Galton 2009], and this might be due to crushing [ Norman 2012]; if so, this feature would be another autapomorphy of Proa ); dentary tooth row extends caudal to the base of the coronoid process; platform between the dentary tooth row and the base of the coronoid process; coronoid process expanded along rostral and caudal margins; maxilla lacks a rostrodorsal process; quadrate straight in lateral view; ilium with dorsal margin convex dorsally, non-pendant supraacetabular process, and postacetabular process that tapers without a break in slope along its dorsal margin; cranial pubic process concave along its dorsal margin but lacks expansion of distal end.
Description. Each premaxilla bears two rostrocaudally elongated denticles, as in Dakotadon (SDSM 8656), Iguanodon ( Norman 1980) , and Mantellisaurus (NHMUK R5764; Norman 1986); between the two denticles is a deep circular fossa ( Fig. 1A View FIGURE 1 ). The premaxillae are slightly expanded transversely, giving the rostral margin of the oral cavity a rounded shape in dorsal view ( Fig. 1B View FIGURE 1 ); this is similar to other styracosternans such as Dakotadon ( Fig. 1C View FIGURE 1 ) (SDSM 8656; Weishampel & Bjork 1989) and Iguanodon ( Norman 1980) , although the premaxillae of Proa taper more towards their rostral ends ( Fig. 1B View FIGURE 1 ). The caudoventral corner of the rostral margin is a broad, gentle curve comparable to those of Dakotadon ( Weishampel & Bjork 1989) , Iguanodon ( Norman 1980) , and Mantellisaurus ( Norman 1986) . The ventrolateral process of the premaxilla is extremely robust and gradually expands dorsoventrally towards its caudal end ( Fig. 1D View FIGURE 1 ).
The maxilla lacks a rostrodorsal process, as in Fukuisaurus ( Kobayashi & Azuma 2003) . The rostroventral process of the maxilla projects rostrally ( Fig. 1E View FIGURE 1 ). The ventral margin of the tooth row is gently concave. The ascending process of the maxilla is rostrocaudally broad and subtriangular with a rounded apex in lateral view ( Fig. 1E View FIGURE 1 ). The caudal margin of the ascending process bears a small semicircular depression, a remnant of the antorbital fossa, as in Iguanodon ( Norman 1980) , Mantellisaurus (NHMUK R5764; Norman 1986), and Ouranosaurus ( Taquet 1976) . Caudoventral to the base of the ascending process on the lateral surface of the maxilla is a shelf that would have been overlapped by the maxillary process of the jugal, forming a ‘scarf’ contact as in Hippodraco ( McDonald et al. 2010b) , Iguanacolossus ( McDonald et al. 2010b) , Dakotadon (SDSM 8656; Weishampel & Bjork 1989), and Fukuisaurus ( Kobayashi & Azuma 2003) . A row of ‘special foramina’ forms an arch dorsal to the tooth row on the medial surface of the maxilla ( Fig. 1F View FIGURE 1 ).
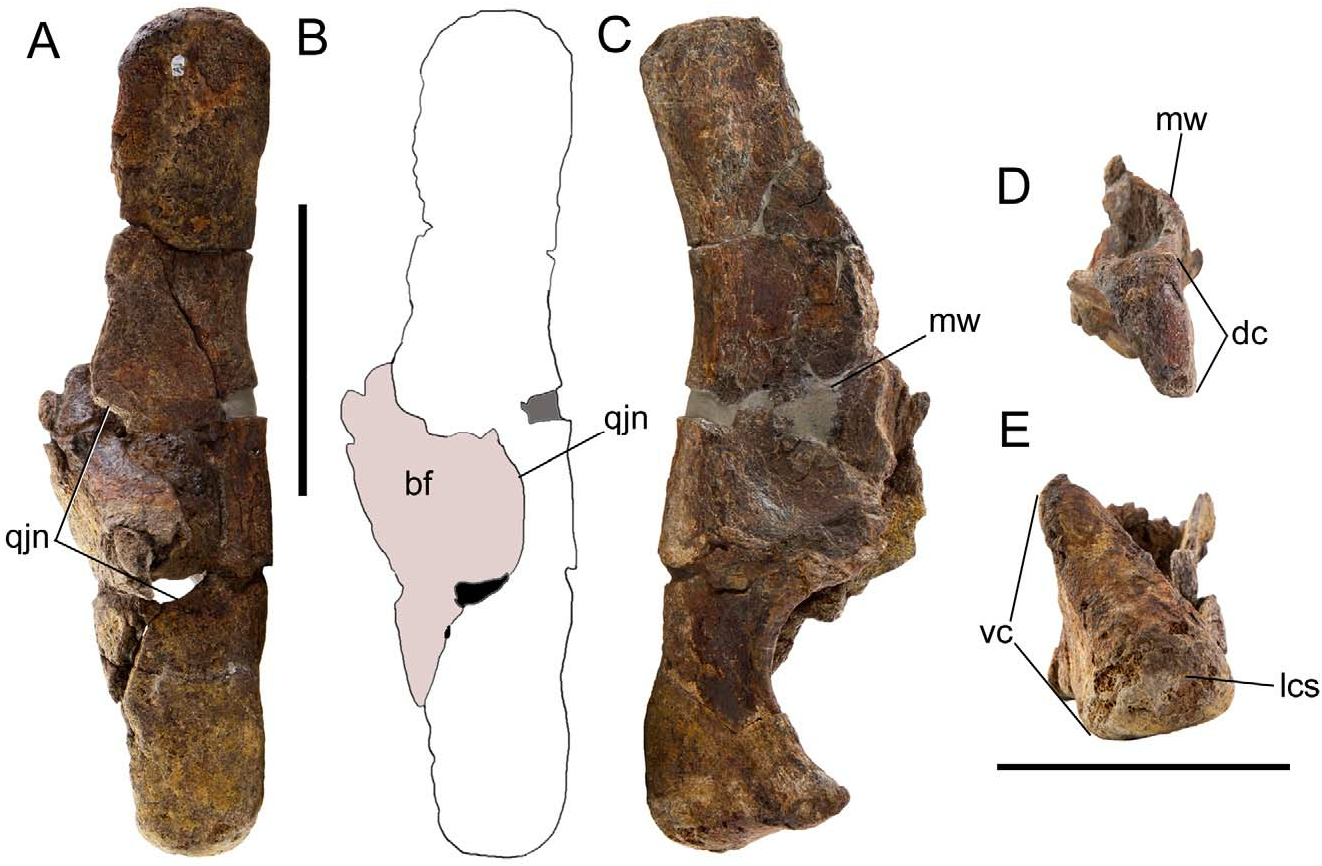
The lateral wing of the quadrate bears a semicircular quadratojugal notch ( Fig. 2A, B View FIGURE 2 ), as in many other basal iguanodonts, such as Iguanacolossus ( McDonald et al. 2010b) , Lurdusaurus ( Taquet & Russell 1999) , Fukuisaurus ( Kobayashi & Azuma 2003) , Iguanodon ( Norman 1980) , Mantellisaurus ( Norman 1986) , Ouranosaurus ( Taquet 1976) , and Jinzhousaurus ( Barrett et al. 2009) . The medial wing of the quadrate is a broad triangular flange that projects rostrally to contact the pterygoid ( Fig. 2C View FIGURE 2 ). The quadrate is straight in lateral view ( Fig. 2A, B View FIGURE 2 ), as in Iguanacolossus ( McDonald et al. 2010b) , Lurdusaurus ( Taquet & Russell 1999) , Fukuisaurus ( Kobayashi & Azuma 2003) , Iguanodon ( Norman 1980) , and Protohadros ( Head 1998) . The dorsal condyle is subtriangular, with a broader rostral margin and tapering towards the caudal margin ( Fig. 2D View FIGURE 2 ). The ventral condyle of the quadrate is rostrocaudally compressed and mediolaterally wide with an enlarged lateral condylar surface ( Fig. 2E View FIGURE 2 ).
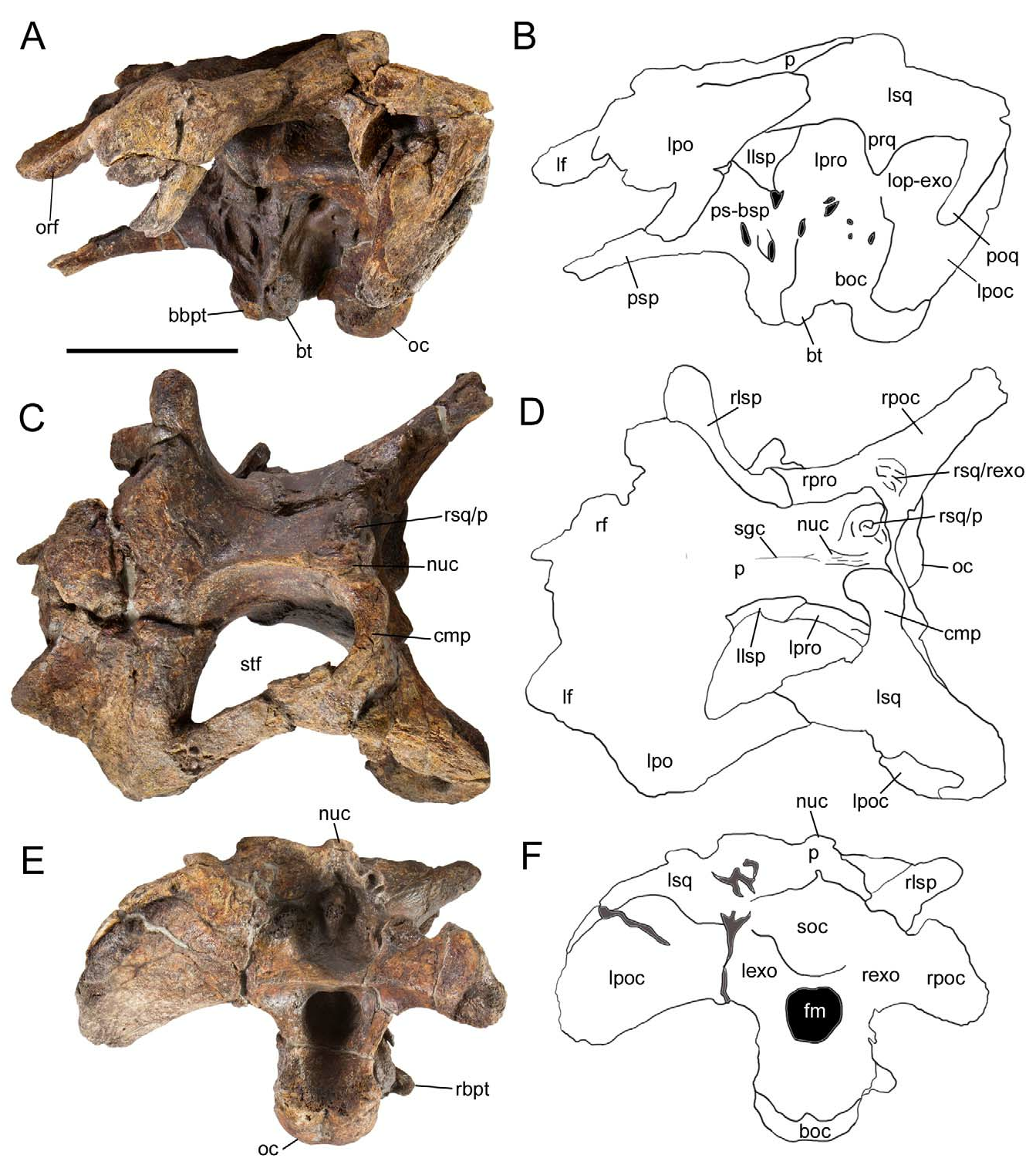
Two exceptionally well preserved braincases are known for Proa ; that of the holotype, AR-1-2012, is briefly described herein. This specimen includes not only the complete uncrushed braincase and skull roof, but also the left postorbital and squamosal. The postorbital forms the caudodorsal margin of the orbit and consists of a central portion and three processes: a medially-projecting platform that contacts the frontal, a rostroventrally-directed process that meets the jugal, and a caudally-directed process that contacts the squamosal to form the dorsal margin of the infratemporal fenestra and the lateral margin of the supratemporal fenestra ( Fig. 3A–D View FIGURE 3 ). The caudal end of the squamosal process is rounded, as in Hippodraco ( McDonald et al. 2010b) , Mantellisaurus ( Norman 1986) , Ouranosaurus ( Taquet 1976) , Altirhinus ( Norman 1998) , Jinzhousaurus ( Barrett et al. 2009) , Equijubus ( You et al. 2003b) , and Xuwulong ( You et al. 2011) . The squamosal exhibits a deep glenoid to receive the dorsal condyle of the quadrate, bounded rostrally by the prequadrate process and caudally by the longer postquadrate process ( Fig. 3A, B View FIGURE 3 ). Rostral to the glenoid is the rostrally-directed postorbital process. Medial to the glenoid, the caudomedial process of the squamosal curves rostromedially; the articulated left squamosal and contact surface for the right squamosal on the parietal indicate that the squamosals were separated by only a narrow sliver of the parietal ( Fig. 3C, D View FIGURE 3 ), as in Jinzhousaurus ( Barrett et al. 2009) , Jintasaurus ( You & Li 2009) , Probactrosaurus (Norman 2002) , Eolambia ( McDonald et al. 2012) , and Bactrosaurus ( Godefroit et al. 1998) .
The frontal forms part of the dorsal margin of the orbit ( Fig. 3A–D View FIGURE 3 ). The laterosphenoid curves laterally to meet the medial surface of the postorbital (3C, D). The supraoccipital is excluded from the foramen magnum by the left and right exoccipitals ( Fig. 3E, F View FIGURE 3 ). The paroccipital process is pendant and ventrally directed ( Fig. 3A, B, E, F View FIGURE 3 ), as in Hippodraco ( McDonald et al. 2010b) , Iguanodon ( Norman 1980) , Bolong ( Wu & Godefroit 2012) , Jintasaurus ( You & Li 2009) , and Probactrosaurus (Norman 2002) , but in contrast to the rostrally curved process of Ouranosaurus ( Taquet 1976) . The basipterygoid processes project ventrolaterally and curve caudally along their lengths. Caudal to the bases of the basipterygoid processes are the rugose basal tubera ( Fig. 3A, B View FIGURE 3 ). The occipital condyle is directed caudoventrally ( Fig. 3A View FIGURE 3 ). The foramen magnum is formed entirely by the exoccipitals, without participation of the basioccipital ( Fig. 3E, F View FIGURE 3 ).
The predentary of Proa is unique among iguanodontians in its overall shape; the lateral processes diverge from each other and the predentary comes to a point at its rostral margin ( Fig. 4A View FIGURE 4 ). This morphology is reminiscent of the predentaries of non-iguanodontian ornithischians such as Lesothosaurus ( Sereno 1991) , Haya ( Makovicky et al. 2011), and Hypsilophodon ( Galton 1974) . In other iguanodontians, the predentary is arcuate, with a broad, rounded rostral margin and rounded rostrolateral corners ( Fig. 4B–F View FIGURE 4 ). The unusual morphology of the predentary of Proa , in conjunction with the tapered premaxillae described above, suggests a feeding ecology different from those of other basal iguanodonts. In other features, the predentary of Proa is similar to those of other basal iguanodonts. There are two grooves on the rostral margin of the predentary, one on either side of the median marginal denticle ( Fig. 5A View FIGURE 5 ). The ventromedial process is bifurcated ( Fig. 5B, C View FIGURE 5 ). The dorsomedial process is a short, caudally-projecting prong that arises caudodorsal to the base of the ventromedial process ( Fig. 5D View FIGURE 5 ). The predentary bears a large conical median denticle with a large denticle on either side of it and smaller denticles along the lateral processes ( Fig. 5A View FIGURE 5 ), as in Dakotadon (SDSM 8656; Weishampel & Bjork 1989), Iguanodon ( Norman 1980) , Mantellisaurus ( Norman 1986) , and Ouranosaurus ( Taquet 1976) .
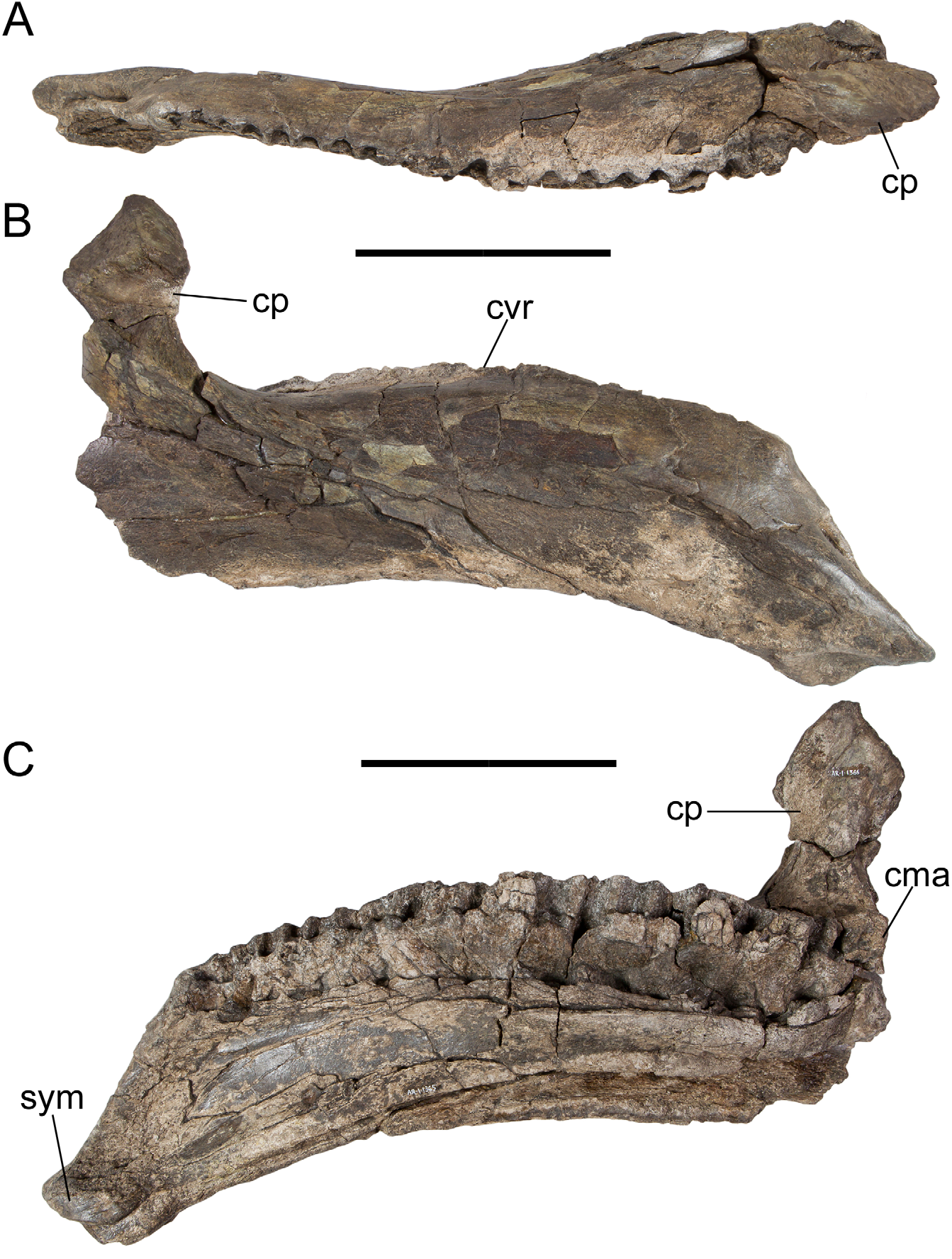
In dorsal view, the dentary tooth row is straight from the first alveolus to approximately the twelfth, at which point it curves caudolaterally towards the base of the coronoid process ( Fig. 6A View FIGURE 6 ). The dentary tooth row is convex dorsally in lateral and medial views ( Fig. 6B, C View FIGURE 6 ), a feature otherwise observed in only Owenodon (NHMUK R2998; Galton 2009), though this morphology might be due to crushing in the latter case ( Norman 2012). The dentary tooth row extends caudally past the base of the coronoid process ( Fig. 6C View FIGURE 6 ), as in hadrosaurids ( Horner et al. 2004) but in contrast to other non-hadrosaurid iguanodontians, such as Mantellisaurus ( Norman 1986) , Ouranosaurus ( Taquet 1976) , Altirhinus ( Norman 1998) , Probactrosaurus (Norman 2002) , and Jeyawati ( McDonald et al. 2010c) ; this feature appears to have evolved independently in Proa . The alveoli mirror the shape of the dentary teeth as in Iguanodon ( Norman 1980) , Mantellisaurus ( Norman 1986) , Ouranosaurus ( Taquet 1976) , and Altirhinus ( Norman 1998) , and in contrast to the alveoli formed by parallel vertical walls in Probactrosaurus (Norman 2002) , Eolambia ( McDonald et al. 2012) , Jeyawati ( McDonald et al. 2010c) , Protohadros ( Head 1998) , Shuangmiaosaurus ( You et al. 2003a) , Bactrosaurus ( Godefroit et al. 1998) , Levnesovia ( Sues & Averianov 2009) , Telmatosaurus ( Weishampel et al. 1993) , and hadrosaurids ( Horner et al. 2004). The dorsal and ventral margins of the dentary are parallel, and the ventral margin is inflected ventrally towards the symphysis ( Fig. 6B, C View FIGURE 6 ). The coronoid process is laterally offset from the tooth row by a narrow shelf, as in Altirhinus ( Norman 1998) , Probactrosaurus (Norman 2002) , Jeyawati ( McDonald et al. 2010c) , Protohadros ( Head 1998) , Shuangmiaosaurus ( You et al. 2003a) , Bactrosaurus ( Godefroit et al. 1998) , and hadrosaurids ( Horner et al. 2004). The vertical coronoid process is rostrocaudally expanded along its rostral and caudal margins as in Altirhinus ( Norman 1998) , Probactrosaurus (Norman 2002) , and Bactrosaurus ( Godefroit et al. 1998) ( Fig. 6B, C View FIGURE 6 ).
The maxillary teeth bear a distally offset primary ridge with multiple fainter accessory ridges mesial and distal to it ( Fig. 7A View FIGURE 7 ). Unworn dentary teeth bear parallel secondary and distally offset primary ridges of similar prominence with multiple faint accessory ridges arising from the marginal denticles ( Fig. 7B View FIGURE 7 ). There appears to be only one replacement tooth per dentary alveolus, and only one active tooth participating in the occlusal plane.
The preacetabular process of the ilium terminates in a horizontal boot ( Fig. 8A View FIGURE 8 ), as in many other styracosternans, including Iguanacolossus ( McDonald et al. 2010b) , Cedrorestes ( Gilpin et al. 2006) , Barilium ( Norman 2011) , Iguanodon ( Norman 1980) , Mantellisaurus ( Hooley 1925; Norman 1986; McDonald 2012a), and Eolambia ( McDonald et al. 2012) . The dorsal margin of the ilium is strongly convex dorsally ( Fig. 8A View FIGURE 8 ), as in Barilium ( Norman 2011) , Iguanodon ( Norman 1980) , Bolong ( Wu & Godefroit 2012) , and Equijubus ( You et al. 2003b) . Dorsal to the ischial peduncle, the dorsal margin of the ilium expands laterally to form a bulbous, non-pendant supraacetabular shelf ( Fig. 8A View FIGURE 8 ), as in Bactrosaurus ( Godefroit et al. 1998) and Gilmoreosaurus ( Prieto-Márquez & Norell 2010) . This structure appears to have evolved convergently in Proa , as other, more derived styracosternans, such as Mantellisaurus ( Norman 1986) , Ouranosaurus ( Taquet 1976) , Xuwulong ( You et al. 2011) , and Probactrosaurus (Norman 2002) , possess only a thickened laterally-everted rim at this location on the ilium. The postacetabular process tapers caudally with no break in slope along its dorsal margin, as in Iguanodon ( Norman 1980) .
The cranial pubic process of the pubis is remarkably similar to those of Camptosaurus (YPM 7334; McDonald 2011: fig. 5), Uteodon ( Carpenter & Wilson 2008; McDonald 2011), and Iguanacolossus ( McDonald et al. 2010b) ; the process is concave along its dorsal margin but its distal end is not expanded ( Fig. 8B View FIGURE 8 ). The morphology of the cranial pubic process of Proa is in strong contrast to the broadly expanded processes of Lanzhousaurus ( You et al. 2005) , Iguanodon ( Norman 1980) , Mantellisaurus ( Hooley 1925; Norman 1986), Delapparentia ( Ruiz-Omeñaca 2011) , Ouranosaurus ( Taquet 1976) , Xuwulong ( You et al. 2011) , Probactrosaurus (Norman 2002) ; and Eolambia ( McDonald et al. 2012) .
The intercondylar extensor groove of the femur is a canal that is fully enclosed by expansion and fusion of the lateral and medial distal femoral condyles ( Fig. 8C, D View FIGURE 8 ), as in Bactrosaurus ( Godefroit et al. 1998) and hadrosaurids ( Horner et al. 2004). The intercondylar flexor groove is partially enclosed by the lateral expansion of the medial condyle ( Fig. 8C, E View FIGURE 8 ). The distal half of the femoral shaft is straight in lateral and medial views ( Fig. 8F View FIGURE 8 ). The fourth trochanter is proximodistally broad and triangular, and arises at approximately the midpoint of the femoral shaft ( Fig. 8E, F View FIGURE 8 ).





No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.