
Neusticurus rudis Marques-Souza et al., 2018
|
publication ID |
https://doi.org/10.1080/00222933.2018.1439541 |
|
publication LSID |
lsid:zoobank.org:pub:33DCF862-11CF-4FD0-B4D6-706E2C6A339E |
|
persistent identifier |
https://treatment.plazi.org/id/37447D69-FF84-8269-FEB8-10E2175DFA48 |
|
treatment provided by |
Felipe |
|
scientific name |
Neusticurus rudis Marques-Souza et al. |
| status |
|
Neusticurus rudis Marques-Souza et al. (in press)
( Figures 5 View Figure 5 , 9 – 11 View Figure 9 View Figure 10 View Figure 11 ; Table 2)
Holotype
IRSNB2687 View Materials , an adult female collected on 11 May 2011 at 9:20 am by PJRK on the summit of Angasima-tepui ( Bolívar state, Venezuela, 5°02.5833N, 62°04.8667W, ca. 2200 m elevation). GoogleMaps
Paratypes (n = 8)
VENEZUELA. Bolívar state, summit of Angasima-tepui , ( 5°02.5833N, 62°04.8667W, ca. 2200 m elevation), one adult male ( IRSNB2690 View Materials ) collected on 14 May 2011 by Brad Wilson and two juveniles ( IRSNB2688 View Materials [formerly IRSNB18151 View Materials ], IRSNB2689 View Materials ) collected on 12 May and 13 May 2011, respectively, by PJRK GoogleMaps . BRAZIL. Roraima state, Pacaraima ( 4° 27.6333N, 61°08.7W, 900 m elevation), two adult males ( MZUSP106223 View Materials , MZUSP106226 View Materials ), two adult females ( MZUSP106224 View Materials , MZUSP106227 View Materials ), and one juvenile ( MZUSP106225 View Materials ) collected in May 2011 by MTR, SMS, AF, MTJ, Augustín Camacho and Renato Recoder GoogleMaps .
Referred material examined is listed in Table 2.
Etymology
The specific epithet is a noun in apposition honouring the Arekuna tribe (also known as the Pemon tribe) that lives in the region of Pantepui where the new species was collected.
Diagnosis
Neusticurus arekuna sp. nov. is characterised by the following combination of characters: (1) size moderate (maximum known SVL 74.5 mm in the holotype, IRSNB2687); (2) tail 1.7 – 2.1 times SVL; (3) tympanum moderately recessed, auditory meatus short; (4) lower eyelid with semi-transparent disc of 5 – 6 palpebrals; (5) frontonasal single; (6) occipital scales in contact with posterior edge of parietals and interparietal 4 – 7; (7) enlarged dorsal tubercles arranged in continuous, although sometimes poorly defined, longitudinal rows containing 40 – 44 scales; (8) flanks with large trihedral scales surrounded by smaller scales, heterogeneous in size; (9) ventral scales in 25 – 27 transverse rows; (10) subdigital lamellae under 4th finger 18 – 21; (11) subdigital lamellae under 4th toe 25 – 33; (12) total number of femoral pores 45 – 51 in males, 36 – 43 in females; (13) tail compressed with 4 – 5 transverse rows of lateral scales corresponding to two subcaudal scales and 2 – 3 dorsal tubercles (= verticils), the last dorsal tubercle of each verticil distinctly overlapping the next verticil; (14) hemipenis with lobes enlarged, distinctly capitate and detached from the hemipenial body; (15) hemipenis with all lateral flounces separated on the central asulcate face by an enlarged nude stripe that occupies ca. one third of the total asulcate face of the hemipenial body.
Comparison with congeneric species
Neusticurus arekuna sp. nov. is immediately distinguished from N. bicarinatus , N. medemi and N. racenisi in having the tympanum moderately recessed (deeply recessed in N. bicarinatus , N. medemi and N. racenisi ), and by a larger number of femoral pores in females ( 36 – 43 in N. arekuna vs 4 – 15 in N. bicarinatus , 9 – 10 in N. medemi and 10 – 15 in N. racenisi ). Neusticurus arekuna sp. nov. is readily distinguished from N. medemi , N. tatei and N. racenisi in having enlarged tubercles on dorsum (absent in N. medemi , N. tatei and N. racenisi ). Further compared with N. tatei , N. arekuna sp. nov. has fewer femoral pores in males ( 45 – 51 in N. arekuna vs 60 – 61 in N. tatei ), and a larger number of femoral pores in females ( 36 – 43 in N. arekuna vs eight in N. tatei ). Neusticurus arekuna sp. nov. is immediately distinguished from N. surinamensis by having a larger number of femoral pores in females ( 36 – 43 in N. arekuna vs 6 – 10 in N. surinamensis ) and by having a single frontonasal (divided in N. surinamensis ). Neusticurus arekuna sp. nov. can be further distinguished from N. medemi , N. racenisi and N. tatei by having hemipenial lobes enlarged, capitate and detached from the hemipenial body, and in having the flounces of the hemipenial body divided by a nude area at the centre of the asulcate face (hemipenial lobes not enlarged, non-capitate, continuous with the hemipenial body, and flounces not interrupted in N. medemi , N. racenisi and N. tatei ). Neusticurus arekuna sp. nov. is most similar to N. rudis from which it can be distinguished by the following combination of characters: enlarged dorsal tubercles arranged in well-defined, continuous longitudinal rows (usually arranged in poorly defined, discontinuous longitudinal rows in N. rudis ) and last dorsal tubercle of each verticil distinctly overlapping the next verticil (poorly overlapping in N. rudis ). Neusticurus arekuna sp. nov. is further distinguished from N. bicarinatus and N. rudis sensu stricto by having the hemipenial lobular tips ornamented with round knobs (hemipenial lobular tips lacking knobs or other visible ornamentation in N. bicarinatus and N. rudis sensu stricto).
Description of holotype
A female 74.5 mm SVL ( Figures 9 – 10 View Figure 9 View Figure 10 ). Tympanum moderately recessed, external auditory meatus short. Rostral well visible from above, wider than long, rounded anteriorly, in contact with anterior supralabials, nasal and frontonasal. Frontonasal single, laterally in contact with nasal and loreal, posteriorly in contact with prefrontals. Prefrontals irregularly pentagonal, in broad contact medially, anteriorly in contact with frontonasal, laterally in contact with loreal, first supraciliary and first supraocular, posteriorly in contact with frontal. Frontal longer than wide, bell-shaped, hexagonal, wider anteriorly, in contact with prefrontals, first three supraoculars and frontoparietals. A pair of pentagonal frontoparietals in broad contact medially, in contact with frontal, the last two supraoculars, parietals and interparietal; suture between frontoparietals indented by interparietal. Interparietal large, longer than wide, roughly hexagonal; interparietal projecting farther posteriorly than the parietals. Parietals irregularly pentagonal, wider than interparietal. Occipitals variable in shape and size, four in contact with posterior edge of parietals and interparietal. Supraoculars four, the anteriormost smallest with two small granules at the posterior angles; 1st supraocular contacting prefrontal, frontal and two first supraciliaries; 2nd supraocular in contact with 2nd and 3rd supraciliaries, and with frontal; 3rd supraocular in contact with frontal and frontoparietal, separated from supraciliaries by a row of small scales; 4th supraocular in contact with frontoparietal and parietal, in contact with 5th supraciliary on the left side and 6th supraciliary on the right side. Supraciliaries five on the left side, six on the right side. Canthus rostralis well- defined, nasal irregularly quadrangular, undivided, nostril near the centre, in contact with rostral, 1st supralabial, loreal (upper and lower segments), and frontonasal. Loreal irregularly pentagonal, divided in two by a horizontal suture, the lower segment smaller; lower segment of loreal in contact with 1st and 2nd supralabials and with nasal; upper segment of loreal in contact with 1st supraciliary, frontonasal, prefrontal, frenocular, and one preocular granules/scales. Frenocular pentagonal, longer than high, in contact with preocular granules/scales, both segments of loreal, 2nd and 3rd supralabials, followed by two suboculars, the 1st half the length of the second. Four postoculars on the right side, five on the left side, uppermost postocular the largest, in contact with temporals, parietal, and separated from supraciliaries and supraoculars by three granular scales on each side. Translucent palpebral disc with five palpebrals. Six supralabials on the right side, eight on the left side; supralabial behind eye the longest. Temporal region with juxtaposed scales, heterogeneous in size and shape, a few weakly keeled. Cephalic scales smooth, with small sensory pits along their edges, except on rostral, frontonasal, first supraocular and upper segment of the loreal on which pits are more homogeneously distributed over the scales. Ear opening large, anterior border ellipsoid, posterior border straight, auditory meatus short.
Mental wider than long, rounded anteriorly, straight posteriorly. Postmental pentagonal, pointed posteriorly, wider than long, posteriorly in contact with a pair of genials, laterally in contact with 1st and 2nd infralabials. Two pairs of genials (chinshields), 1st pair medially in broad contact, separated by a small medial scale posteriorly, laterally in contact with infralabials; 2nd pair narrower but longer, medially separated by four small scales; both pairs in broad contact with infralabials; second pair of genials followed by a row of three enlarged scales. Infralabials four. Pre-gulars smooth, round to ovoid, juxtaposed, slightly increasing in size laterally. No visible line of scales separates gulars from pre-gulars. Gulars smooth, slightly smaller anteriorly, increasing in size posteriorly, smaller scales juxtaposed, larger scales sometimes slightly imbricate. One row of paired enlarged gulars. Collar distinct, containing eight enlarged scales, collar fold covered with smooth juxtaposed granules.
Dorsum of neck covered with trihedral tubercles, in contact longitudinally, arranged in continuous rows separated by series of small, irregularly disposed and juxtaposed scales. Lateral neck scales strongly protruded, trihedral, arranged in longitudinal rows as on dorsal part of neck. Dorsum with two different types of scales: (1) small, juxtaposed smooth or slightly keeled scales, and (2) large, prominently keeled ovoid tubercles. Dorsal tubercles disposed rather regularly, arranged in longitudinal rows of 41 scales (from occipital scales to posterior margin of hind limb). Most tubercles aligned. Tubercles smaller and more rounded dorsolaterally, becoming conical, although still keeled, on flanks where they are arranged in poorly defined oblique rows anteriorly. Ventrals smooth, slightly imbricate, rounded posteriorly in 27 transverse and eight longitudinal rows (including one row of ‘ lateral scales ’ on each side). Scales on preanal plate equal in size to, or smaller than, ventral scales, mostly pentagonal or irregularly quadrangular. Median scale largest, slightly overlapping a smaller posterior scale. The latter is part of a transverse row of seven scales delineating the posterior margin of the preanal plate. Two slightly smaller (about two thirds of the median scale), irregularly hexagonal scales on each side of the median scale. Two much smaller (about one fourth of the median scale), irregularly quadrangular scales anterior to median scale. Four small rounded or ovoid scales border the lateral margins of the preanal plate, the two anteriormost in contact with femoral pores. Total number of femoral pores 36, in a continuous row separated medially from the preanal plate by two scales. Most femoral pores surrounded by 4 – 6 small scales, giving it a flower-like appearance.
Tail compressed, incomplete, tip regenerating, with a continuous double row of tubercles. Each tail segment (verticil) corresponds to two subcaudal scales and contains 4 – 5 transverse rows of keeled lateral scales and 2 – 3 keeled dorsal tubercles, the last dorsal tubercle of each verticil distinctly overlapping the next verticil. Subcaudals smooth, imbricate, quadrangular and with convex posterior margin.
Scales on dorsal surface of forelimbs smooth or slightly keeled, imbricate, with rhomboidal distal margin, arranged in longitudinal rows. Scales become smoother and smaller towards the ventral surface of the forelimbs. Scales on hand smooth, rhomboidal and strongly imbricate. Hind limbs with keeled rhomboidal and imbricate scales anteriorly; proximal and posterior region covered with granular scales. Subdigital lamellae on hand and foot smooth and single; subdigital lamellae on foot slightly tuberculate at the base. Number of lamellae on 4th finger 21, on 4th toe 29. Palms and soles covered with small, granular scales.
Colour of holotype in life ( Figure 10 View Figure 10 )
Head greenish brown dorsally and laterally, one vivid yellow spot on posterior of lower lip, under the eye; similar spots occur on the lower temporals and post-genial scales. Eye surrounded by a yellowish-brown ring; iris greenish. Dorsal ground colour of body and tail dark olive brown; a series of dorsolateral yellowish-ochre squarish spots extends from neck to ca. eighth verticil on tail. Light irregular yellowish-ochre spots on flanks corresponding to one or more tubercles, which in combination with the dorsolateral spots make an inconspicuous checked pattern. Limbs dark olive brown with scattered yellowish-ochre spots, posterior surface of thighs uniform dark brown. Ventral surface of head and throat creamy white with numerous dark brown markings, more extensive on genials and laterally. Venter whitish anteriorly and medially, with an orange tint posteriorly; irregular dark brown flecks on lateral ventral scales. Ventral surface of forelimbs greyish white, of hind limbs orangish-grey. Ventral surface of tail cream with numerous dark brown flecks increasing in number posteriorly where the tail becomes completely dark brown. Palms and soles dark brown.
Colour of holotype in preservative ( Figure 9 View Figure 9 )
Similar to colour in life, but less vividly marked. Vivid yellow spots on upper and lower lip, on lower temporals and on post-genial scales became white. Dorsal surfaces mostly dark greyish-brown, yellowish-ochre markings became light brown. Ground colour of ventral surfaces more uniform, creamy white with dark brown markings as described above.
Hemipenis ( Figure 5 View Figure 5 )
The hemipenis of the paratype MZUSP106223 (Pacaraima, Brazil) is bilobed, with the hemipenial body ornamented with 10 transversal flounces with no vestige of spines or any other calcified structure. Hemipenial body roughly cylindrical. Lobes distinctly detached from hemipenial body, capitate and with a pair of rounded knobs ornamenting the tip of each lobe. The lobular surface is nude, with no flounce or other ornamentation. The sulcus spermaticus is deep and central in position, originating at the base of the organ, and proceeding in a straight line towards the lobular basis, bifurcating just before the lobular crotch. A small V-shaped fleshy fold with conspicuous projections at the tip of the lobular crotch divides the sulcus, and the resulting branches extend across the lobes, ending between the knobs at lobular tips. Two narrow areas with no ornamentation are parallel to the sulcus spermaticus. A wide nude area separates the 10 hemipenial body transversal flounces into two lateral sets on the asulcate face. Each of the body flounces runs downwards from the sulcate face, changing its direction upwards and returning to the downward direction on the sides towards the asulcate face, making a Wshape. The preparation of a hemipenis of the paratype IRSNB2690 (Angasima-tepui, Venezuela) is suboptimal (inflation slightly incomplete), but hemipenial morphology is similar to MZUSP106223.
Variation in the type series ( Figure 10 View Figure 10 )
Most variation in the type series is covered in the diagnosis; see also Table 2. All paratypes except two paratopotypes ( IRSNB2688 View Materials and IRSNB2690 View Materials ) have an undivided loreal as in the holotype, making the undivided loreal the most common pattern in the species (found in 78% of the specimens examined) . Compared to the holotype, other minor differences are found in the scalation: the interparietal may rarely be transversally divided; the two occipitals posterior to the interparietal are often dorsally elongated, crossing the whole occipital area. Occipital scales in contact with the posterior edge of parietals 4 – 7 (four in holotype). There is significant variation in colour pattern: ground dorsal colour varies from dark greenish/olive brown to chestnut brown and dorsal pattern varies from conspicuous to almost absent ( Figure 10 View Figure 10 ). Some of this variation could apparently be attributed to ontogeny or sexual dimorphism, although our sample is too small to support these conclusions. Juveniles are rather variable, one juvenile paratopotype ( IRSNB2689 View Materials ) has a broad chestnut brown dorsal band and an inconspicuous series of dorsolateral ochre squarish spots as in the holotype . Additional discreet squarish spots on flanks make an inconspicuous checked pattern as in the holotype. The other juvenile paratopotype ( IRSNB2688 View Materials ) lacks a dorsal band but also has an inconspicuous series of dorsolateral ochre squarish spots; no checked pattern occurs in that specimen . Among paratypes from Pacaraima, the checked pattern on flanks is visible in the juvenile ( MZUSP106225 View Materials ) and in females ( MZUSP106224 View Materials , MZUSP106227 View Materials ), although less conspicuous in the latter . The checked pattern is absent in males from Pacaraima and inconspicuous in the male from Angasima-tepui.
Distribution and natural history
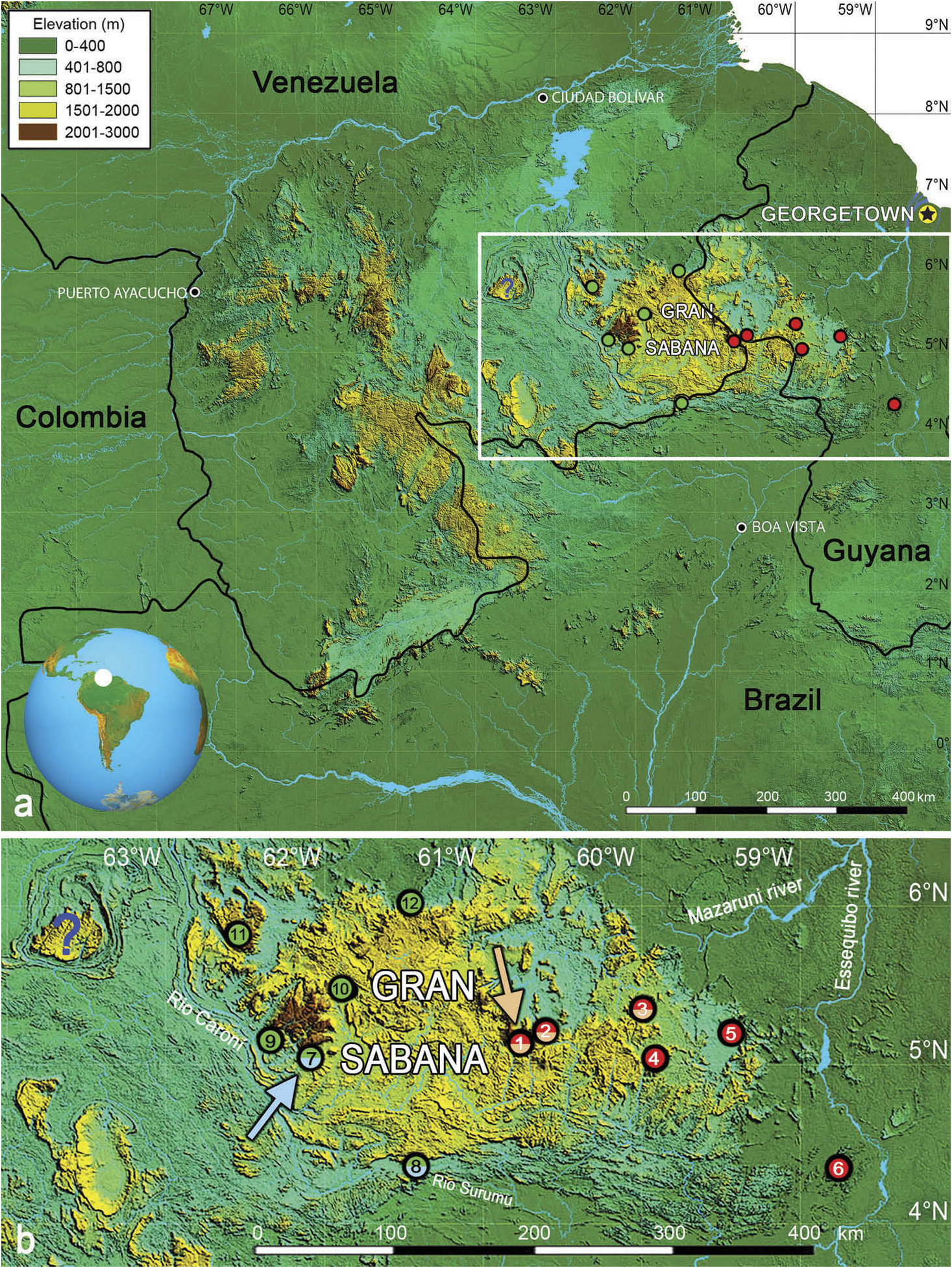
Neusticurus arekuna sp. nov. sensu stricto is currently only known from two localities west of the Venezuelan Gran Sabana: the summit of Angasima-tepui in Bolívar state, Venezuela ( 2162 m elevation), and a stream on the southern slope of the Serra Pacaraima in Roraima state, Brazil ( 900 m elevation, Figure 1 View Figure 1 ). The unnamed stream flows into the Rio Surumu in the Rio Branco Basin. The new species is expected to occur between these two localities. Other populations provisionally referred to that species (see above and Discussion) are currently found exclusively west of the Venezuelan Gran Sabana, from the La Escalera region to Chivatón, the summit of Abakapá-tepui, and the slopes and summit of Auyán-tepui, Venezuela, between 1100 and 2203 m elevation.
Neusticurus arekuna sp. nov. seems ecologically highly plastic as it was found in very different habitats and along a broad elevational range ( Figure 11 View Figure 11 ). Macrohabitat at the type locality (summit of Angasima-tepui) is characterised by tepui-summit scrub and meadows on peat and rock ( McDiarmid and Donnelly 2005). All specimens at the type locality were collected by day; the holotype was swimming across a small puddle on rocks, IRSNB2688 View Materials was found under a rock, IRSNB2689 View Materials was found 170 cm above the ground on a rock wall and IRSNB2690 View Materials was collected walking among pitcher plants ( Heliamphora sp ) . Other paratypes (from Pacaraima , Brazil) were found in a more typical habitat for Neusticurus , a cascading stream in primary forest, with large granitic boulders .
Geographic variation within the N. arekuna clade ( Figure 12 View Figure 12 ; Table 2)
Dorsal tubercles in adult specimens from Chivatón ( Neusticurus arekuna UCS 1; two females) and the summit of Abakapá-tepui ( Neusticurus arekuna UCS 1; one male) are generally larger (especially on the tail) and more strongly keeled than in specimens from other populations in the clade. Conversely, four adult female specimens from the La Escalera region ( Neusticurus arekuna UCS 2) have distinctly smaller and less keeled dorsal tubercles than other populations in the clade; these tubercles are distributed in a similar way to those in the rudis clade (arranged in poorly defined, discontinuous longitudinal rows, see Figure 12 View Figure 12 ) making confusion with N. rudis easy. Two adult female specimens from Auyán-tepui ( Neusticurus arekuna UCS 3) are morphologically very similar to Neusticurus arekuna sp. nov. sensu stricto, and the two populations apparently have a similar hemipenial organ (as described in Myers and Donnelly 2008), while the hemipenis from the population of Abakapá-tepui shows significant differences compared to the hemipenis of Neusticurus arekuna sp. nov. sensu stricto, regarding the hemipenial body and lobular shapes. The hemipenis of the specimen from Abakapá-tepui (IRSNB18150) has distinctly reduced lobes, not detached from the hemipenial body or capitate; the lobular tips lack any visible ornamentation, but have a pair of knobs directed basally at the basis of their sulcate face. On the other hand, the hemipenis of the specimen of Neusticurus arekuna sp. nov. sensu stricto from Pacaraima (MZUSP106223) has enlarged lobes that are distinctly capitate and detached from the hemipenial body, with round knobs ornamenting the lobular tips, but lack the knobs at the basis of the sulcate face. Moreover, the specimen from Pacaraima has an elongate and roughly conical hemipenial body, whereas the specimen from Abakapátepui has a short and globose hemipenis. Hemipenes from the populations of La Escalera and Chivatón are currently unavailable.





| SMS |
Missouri State University |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |