
Proutia nigra Saigusa et Sugimoto, 2014
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3869.2.3 |
|
publication LSID |
lsid:zoobank.org:pub:8AE80644-6D1B-487B-BE9F-FF5AC1A3C37B |
|
DOI |
https://doi.org/10.5281/zenodo.4947201 |
|
persistent identifier |
https://treatment.plazi.org/id/376A87F5-274F-FF81-5BA1-FD044555FD03 |
|
treatment provided by |
Felipe |
|
scientific name |
Proutia nigra Saigusa et Sugimoto |
| status |
sp. nov. |
Proutia nigra Saigusa et Sugimoto View in CoL , sp. nov.
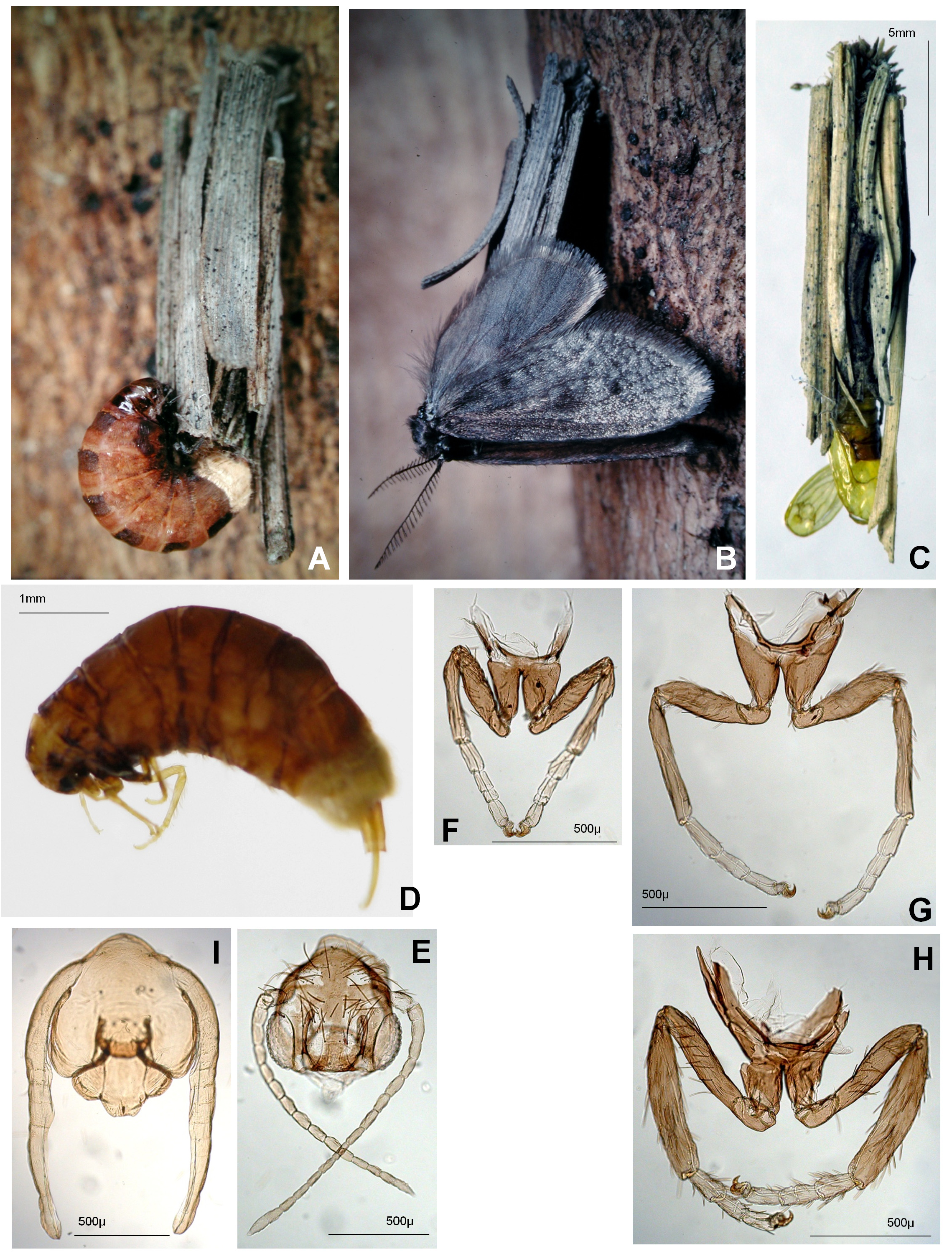
( Figs. 3B View FIGURE 3 , 4B View FIGURE 4 , 5 View FIGURE 5 , 6 View FIGURE 6 )
Diagnosis. Male: Median-sized species of Proutia , with pointed narrow wings, forewing upperside blackish brown to dark brown, without markings. Female: Antenna very short, nearly as long as width of head; abdominal membrane light beige in colour.
Description. Male ( Fig. 2 View FIGURE 2 ): Wing expanse 10.4– 1.30 mm. Coloration of vestiture. Head clothed with blackish to dark brown hair-like scales, mixed with a few greyish ones in front. Dorsal surface of antennal shaft covered with dark brown scales mixing a few greyish yellow scales on posterior margin of basal 1/2 and occasionally on apex of each flagellomere. Thoracic nota and abdominal terga clothed with dark brown more or less broad hair-like scales. Legs covered with dark brown scales, mixing somewhat paler ones at tips of tarsomeres and spurs of mid and hind legs. Forewing ( Fig. 5B View FIGURE 5 ) upperside uniformly brownish black in fresh specimens, dark to blackish brown in aged specimens. Hindwing ( Fig. 5B View FIGURE 5 ) upperside slightly lighter than forewing. Fringe of both wings uniformly dark brown.
Antenna ( Fig. 5I View FIGURE 5 ) slightly shorter than forewing including fringe (0.43–0.46); flagellum bipectinated and consisting of 17–19 (M. 18 n=10) flagellomeres; pectination on 7th flagellomere longest, 350–400µ (M. 374 µ) long, and 2.0–2.75 times as long as flagellomere; pectinations abruptly becoming longer to 7th flagellomere, then strongly decreasing in length towards distal flagellomere. Fore tibia with epiphysis 0.57–0.69 (M. 0.65) times as long as fore tibia, arising from 0.29–0.45 (M. 0.36) length of tibia from base.
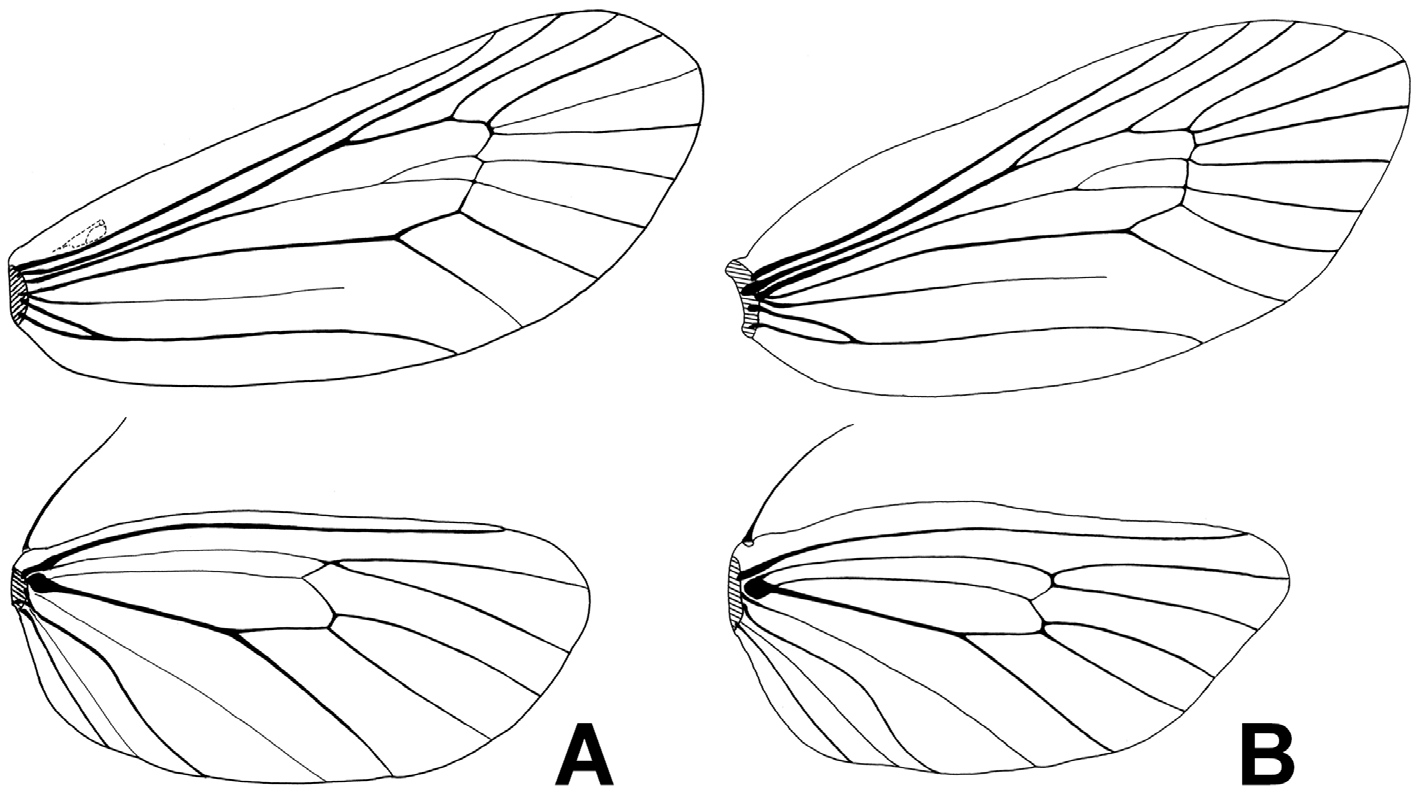
Forewing 4.2–5.2 mm (M. 4.9 mm) in length excluding fringe, 2.15–2.41 (M. 2.31) times as long as wide, with termen distinctly produced; forewing length including fringe 5.2–6.2 (M. 5.64) mm; discoidal cell 0.66–0.72 (M. 0.69) times as long as forewing. Hindwing 3.2–3.9 (M. 3.7) mm long excluding fringe, 1.94–2.22 (M. 2.08) times as long as wide, with termen distinctly produced and roundly pointed; discoidal cell 0/66–0.72 (M. 0.69) times as long as hindwing. Forewing venation ( Fig. 3B View FIGURE 3 ): Accessory cell absent; intercalary cell present and long; forking point of vein M in discoidal cell usually at level between origins of veins R1 and R2; bases of all veins from cell separated from each other. Hindwing venation ( Fig. 3B View FIGURE 3 ): Veins Rs and M1 usually separated, but rarely connate; vein M in discoidal cell simple.
Wing vestiture. Forewing upperside covered with broad upper scales 130–160 µ (M. 150 µ) long, 30–40 µ (M. 34 µ) wide, distal margin of most scales usually with 4 serrations, occasionally 3 or 5, length of dentation usually slightly longer than 10 µ; lower scales of upperside of forewing 55–85 µ (M. 68 µ) long, 14–23 µ (M. 20 µ) wide, distal margin of most of scales usually with 3 serrations 3–10 µ in length. Hindwing upperside covered with broad upper scales 110–180 µ (M. 152 µ) long, 15–25 µ (M. 20 µ) wide, distal margin of most scales usually with 2 serrations, occasionally simple, length of dentation 5–10 µ; lower scales of upperside of forewing 50–75 µ (M. 62 µ) long, 10–17 µ (M. 14 µ) wide, distal margin of most of scales with 2 serrations 3–8 µ in length.
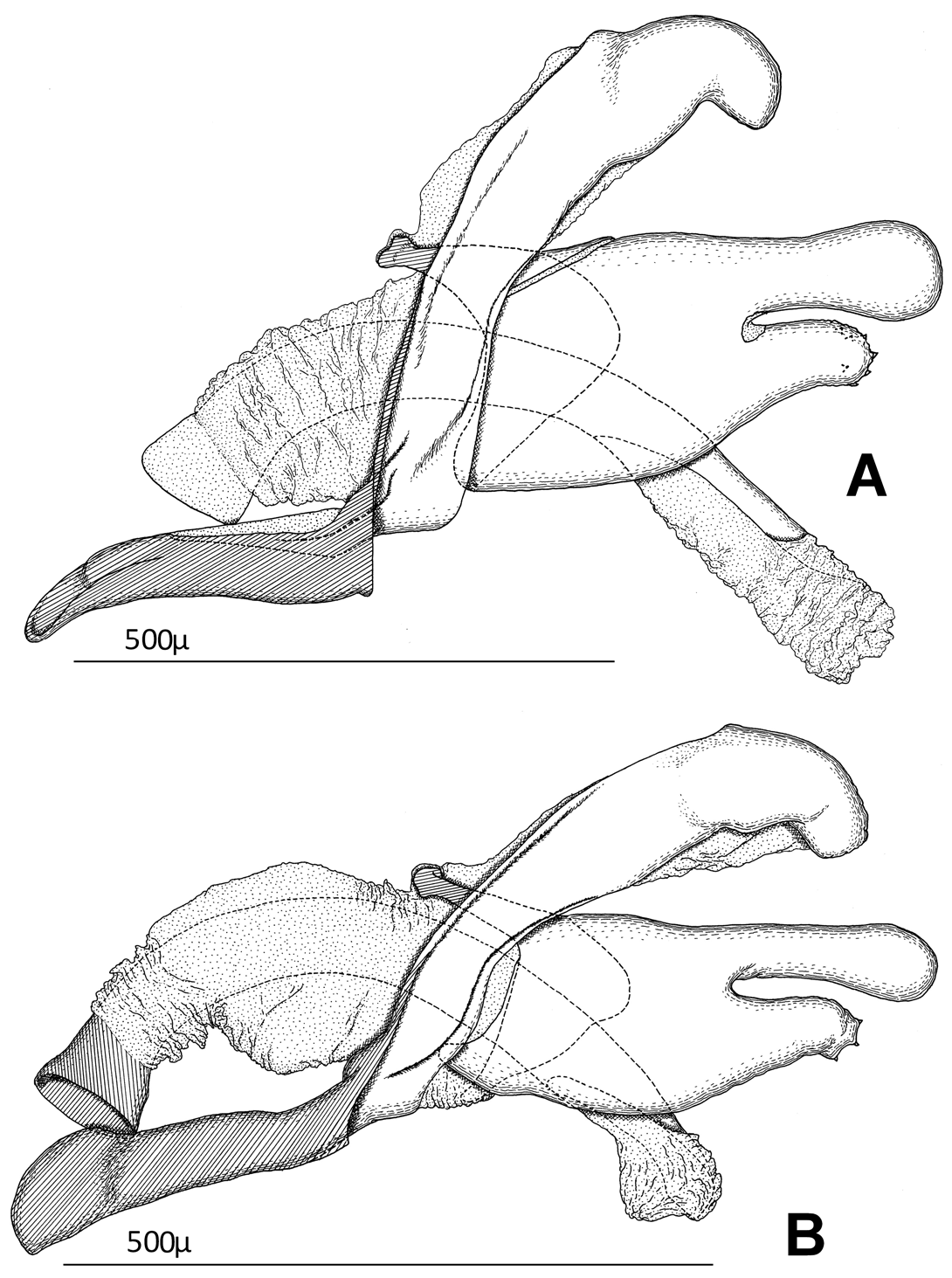
Male genitalia ( Figs 5 C–F View FIGURE 5 , 4B View FIGURE 4 ): Similar to preceding species, ampulla of valva ( Fig. 5E View FIGURE 5 ) longer and more slender. Relative length of ampulla to dorsal margin of valva (including ampulla and excluding transtilla) 0.56–0.75; ampulla 4.2–4.3 times as long as narrowest width; distal margin of dorsum ( Fig. 5C View FIGURE 5 ) weakly produced with shallow median notch; phallus ( Fig. 5 F View FIGURE 5 ) rather strongly curved ventrally without denticles of vesica; anellus ( Fig. 5 D View FIGURE 5 ) bearing only fine setulae.
Female ( Figs 6A–E View FIGURE 6 ). Coloration. Sclerites of head, thorax and legs light yellowish brown, abdominal terga and sterna brown, slightly lighter than P. maculatella sp. nov.. Meso- and metanotum with irregular darker dorsal and sublateral markings succeeding to markings of mature larvae. Membranous areas of abdomen light beige in colour, much paler than P. maculatella sp. nov.. Abdomen sparsely clothed with light greyish brown fine hair-like scales. Corethrogyne yellowish white.
Structure. Antenna ( Fig. 6B View FIGURE 6 ) short, 350–580 µ long, 0.8–1.3 times as long as head width. Flagellum indistinctly divided into 4–7 flagellomeres. Legs. ( Figs. 6C–E View FIGURE 6 ) with tarsi divided into 3 tarsomeres, occasionally mid and hind tarsi into 4 and 5 respectively; fore leg not much short as in P. chinensis .
Distance from head to apical part of abdominal segment 7 in natural posture curving ventrally: 3.1–4.3 mm.
Type material. HOLOTYPE ♂ (with pupal case and exuviae), Innakiyama , Miyawaka-shi, Fukuoka Pref., Kyushu, Japan, May 12, 1969, T. Saigusa. Donated to the Kyushu University Museum . PARATYPES (most paratypes with pupal case and exuviae): 1♂, same locality as holotype, May 10, 1958, T. Saigusa ; 3♂, same locality, May 3–8, 1961, T. Saigusa ; 7♂ (1♂ macerated and right wing venation on a slide), same locality, May 9–14, 1967, T. Saigusa ; 1♂, same data as holotype ; 1♂ (only wings on a slide), same locality, May 19, 1970, T. Saigusa ; 1♂ (macerated and right wings and left wings venation on slides), same locality, May 25, 1970, T. Saigusa ; 1♂ (only wings on slide), May 14, 1971, T. Saigusa ; 9♂ (1♂ macerated and right wing venation on slide, 1♂ (only wings on a slide), same locality, May 13–19, 1972, T. Saigusa ; 3♂ (each macerated and right wings on a slide), same locality, May 22, 1999, T. Saigusa ; 9♀ (in ethanol), same locality, May 3–22, 1999, T. Saigusa ; 8♀ (in ethanol), same locality, May 5–8, 2000, T. Saigusa .
Type locality: Innakiyama (Inunakiyama), Miyawaka-shi, Fukuoka Pref., Kyushu, Japan .
Distribution. Kyushu.
Remarks. The male of P. nigra sp. nov. most resembles that of P. betulina , but differs from the latter in more strongly pointed wings, especially hindwings, and the wide tegumen and longer ampulla and harpe of the genitalia. The female of the new species differs from betulina in strongly shortened antennae and the yellowish white corethrogyne, that is snow-white (silver, silverweiss) in betulina ( Meyrick 1927; Kozhantshikov 1956; Hättenschwiler 1985). The covering of larval case of betulina is different from that of the new species, and consists of short broad pieces of plant material, such as bark, pine-needles, grass and lichen ( Hättenschwiler 1985; Hättenschwiler et Chao 1990). Proutia nigra sp. nov. also differs from P. breviserrata , P. norvegica and P. rotunda in its pointed wings. The valva of P. breviserrata is more slender than that of nigra . Proutia norvegica has light brown wings. The larval case of P. rotunda is somewhat similar to P. betulina , and consequently quite different from that of P. nigra sp. nov. ( Palmqvist 2008). The Chinese P. chinensis is similar to P. nigra sp. nov. in uniformly dark forewings, but the former has flagellomere pectinations much longer than P. nigra sp. nov. in proportion to flagellomeres, and has no distinctive intercalary cell in the forewing discoidal cell ( Hättenschwiler et Chao 1990). The female of P. chinensis resembles P. nigra sp. nov. female in the abbreviated antennae, but it differs from P. nigra sp. nov. in extremely shorter forelegs in proportion to the mid and hind legs ( Hättenschwiler et Chao 1990).
The habitat of this species was found in evergreen forests and Japanese cedar plantations ( Cryptomeria japonica ), but not in open grasslands, the main habitat of P. maculatella sp. nov. The type locality is in the low mountain zone about 400 m in altitude. The species was not found in lowlands in northern Kyushu. Adults appear in late April to mid May.
The mature larvae fix their pupation cases on fences, walls of deserted sheds, tree trunks, etc in forests in mid spring. This species seems to be univoltine, but this has not been confirmed by breeding. The larval case ( Fig. 6G View FIGURE 6 ) is similar to that of P. maculatella sp. nov., however its covering consists of needles of Japanese cedar and other material found on the forest floor, not of narrow pieces of herbaceous plants attached to cases of P. maculatella sp. nov. Head ( Fig. 6F View FIGURE 6 ) and forelegs of pupal exuviae of females are not shed, so that females wear the pupal head cuticle as a mask and the forelegs can not grasp the posterior end of the pupation case.


| T |
Tavera, Department of Geology and Geophysics |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |