
Kimminsula, Peters & Edmunds, 1970
|
publication ID |
https://doi.org/10.11646/zootaxa.5212.1.1 |
|
publication LSID |
lsid:zoobank.org:pub:485EA061-41EA-4E0C-894D-593B84DB41D8 |
|
DOI |
https://doi.org/10.5281/zenodo.7400863 |
|
persistent identifier |
https://treatment.plazi.org/id/380E87A6-F168-FF9A-3181-FE23FDD45CE4 |
|
treatment provided by |
Plazi |
|
scientific name |
Kimminsula |
| status |
|
Kimminsula View in CoL -complex
( Figs 3 View FIGURES 1–6 , 7–710 View FIGURES 7–15 View FIGURES 16–26 View FIGURES 27–29 View FIGURES 30–35 View FIGURES 36–42 View FIGURES 43–50 View FIGURES 51–58 View FIGURES 59–62 View FIGURES 63–67 View FIGURES 68–71 View FIGURES 72–77 View FIGURES 78–80 View FIGURES 81–84 View FIGURES 85–93 View FIGURES 94–100 View FIGURES 101–107 View FIGURES 108–113 View FIGURES 114–118 View FIGURES 119–123 View FIGURES 124–129 View FIGURES 130–144 View FIGURES 145–153 View FIGURES 154–165 View FIGURES 166–171 View FIGURES 172–181 View FIGURES 182–187 View FIGURES 188–195 View FIGURES 196–204 View FIGURES 205–208 View FIGURES 209–218 View FIGURES 219–220 View FIGURES 221–228 View FIGURES 229–235 View FIGURES 236–242 View FIGURES 243–250 View FIGURES 251–260 View FIGURES 261–271 View FIGURES 272–277 View FIGURES 278–289 View FIGURES 290–293 View FIGURES 294–299 View FIGURES 300–305 View FIGURES 306–310 View FIGURES 311–316 View FIGURES 317–320 View FIGURES 321–331 View FIGURES 332–335 View FIGURES 336–338 View FIGURES 339–347 View FIGURES 348–368 View FIGURES 369–373 View FIGURES 374–376 View FIGURES 377–382 View FIGURES 383–386 View FIGURES 387–394 View FIGURES 395–405 View FIGURES 406–412 View FIGURES 413–416 View FIGURES 417–420 View FIGURES 421–427 View FIGURES 428–431 View FIGURES 432–435 View FIGURES 436–439 View FIGURES 440–443 View FIGURES 444–451 View FIGURES 452–457 View FIGURES 458–463 View FIGURES 464–467 View FIGURES 468–473 View FIGURES 474–475 View FIGURES 476–486 View FIGURES 487–495 View FIGURES 496–498 View FIGURES 499–506 View FIGURES 507–527 View FIGURES 528–538 View FIGURES 539–544 View FIGURES 545–549 View FIGURES 550–551 View FIGURES 552–561 View FIGURES 562–568 View FIGURES 569–573 View FIGURES 574–580 View FIGURES 581–586 View FIGURES 587–600 View FIGURES 601–606 View FIGURES 607–615 View FIGURES 616–619 View FIGURES 620–625 View FIGURES 626–631 View FIGURES 632–640 View FIGURES 641–663 View FIGURES 664–672 View FIGURES 673–682 View FIGURES 683–687 View FIGURES 688–693 View FIGURES 694–699 View FIGURES 700–706 View FIGURES 707–710 )
The Kimminsula -complex belongs to subsequently subordinated taxa: Leptophlebiidae >Atalophleboadentata Kluge 2009 > Atalophlebopectinata Kluge 2009 > Atalophleboculata Kluge 2009 > Atalophlebomaxillata Kluge 2009 > Atalophlebolinguata Kluge 2009. General system of the taxon Atalophlebolinguata remains poorly elaborated.
The Kimminsula -complex is distributed only in the Indian Peninsula and the Island of Sri Lanka and includes Kimminsula Peters & Edmunds 1970 (with 3–5 species), Petersula Sivaramakrishnan 1984 (with 2–3 species), Ghatula gen. n. (with 2 species), Ceylonula gen. n. (with 1 species) and Hubbardula gen. n. (with 1 species). Larvae of all these species are similar in general appearance [see (1) below]; characters separating larvae of the Kimminsula -complex from other taxa with a similar larval appearance are the presence of fronto-clypeal incisions [see (2) below] and a characteristic shape and setation of femora [see (10) below]. Imagines of the Kimminsula -complex differ from other Oriental Leptoplebiidae by well developed hind wings of plesiomorphic shape and venation [see (16) below] and by the claw structure [see (17) below].
Common characters of the Kimminsula -complex
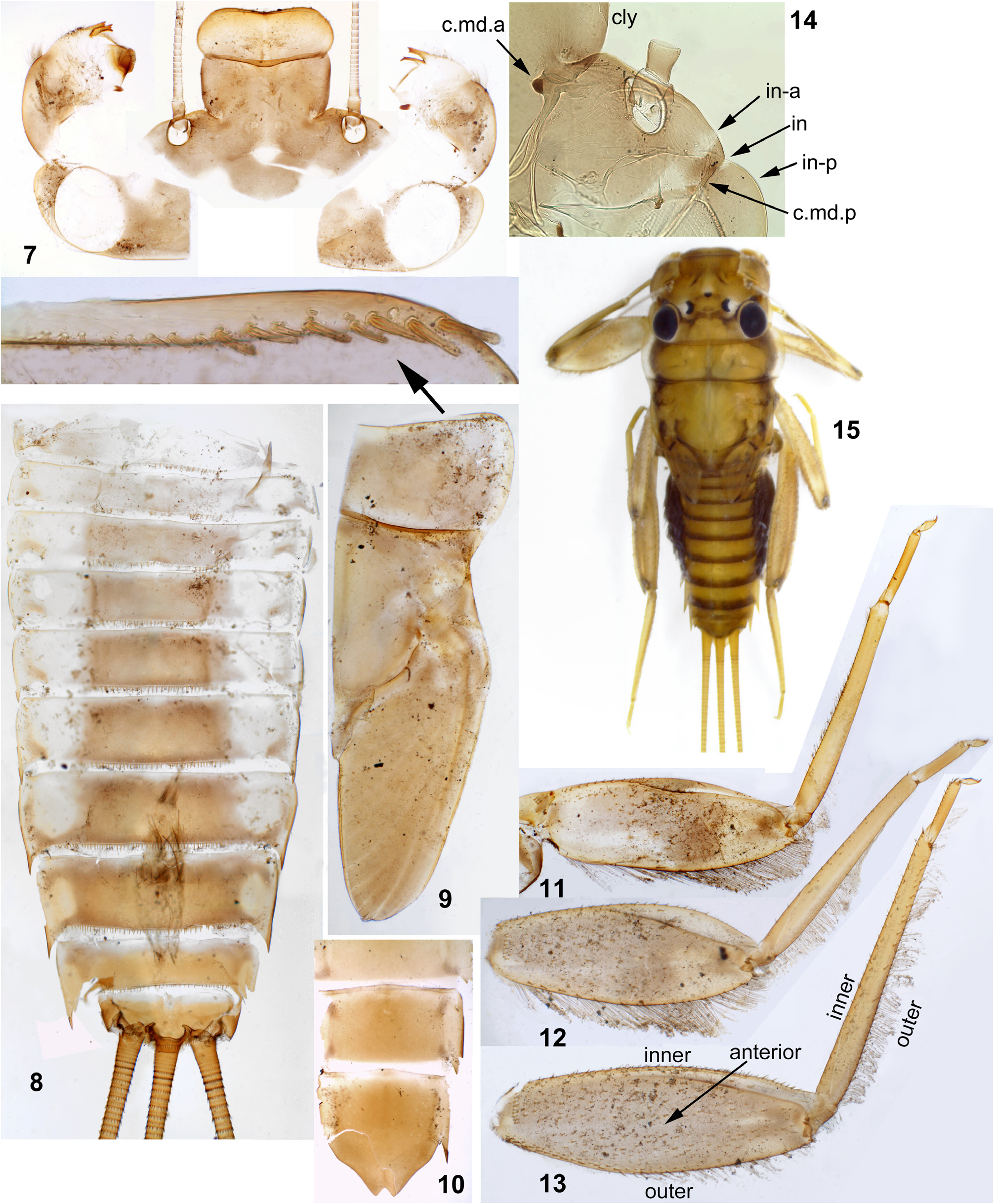
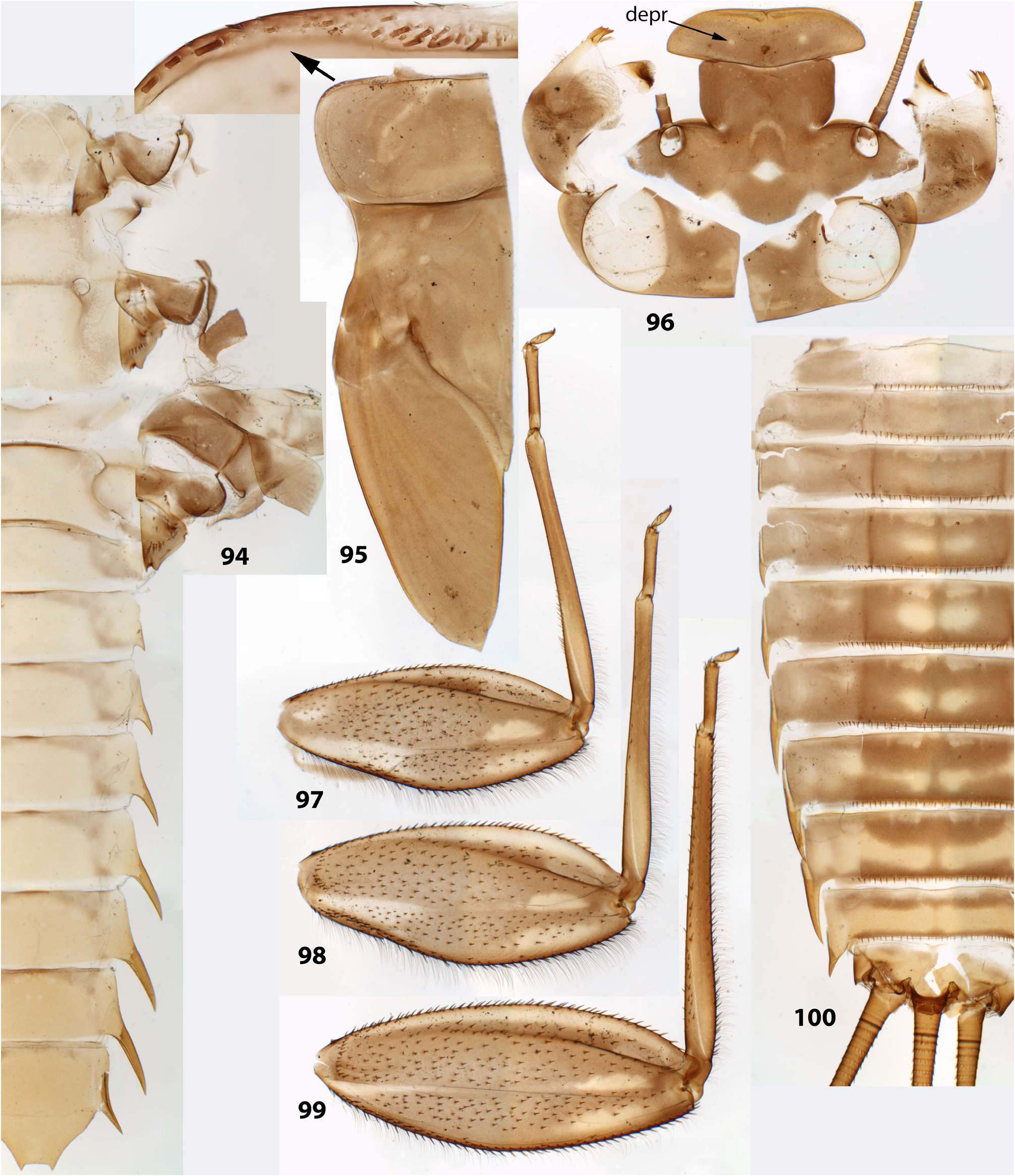
(1) General appearance. Larva has flattened body, widened head and enlarged legs with broad femora and claws bent perpendicular to the leg flatness ( Figs 202 View FIGURES 196–204 , 440–443 View FIGURES 440–443 ). Superficially, they resemble larvae of Heptageniidae , but in contrast to them have no frontal shield; instead, the wide dorsal head surface contains surfaces of closely pressed together clypeus, labrum and mandibles ( Fig. 235 View FIGURES 229–235 ) (the same in other Atalophlebomaxillata). Initially such larval structure is an adaptation for inhabiting stone surfaces in rapidly running water ( Fig. 219 View FIGURES 219–220 ), but larvae of Ceylonula femoralis inhabit soft substrates in stagnant water ( Fig. 220 View FIGURES 219–220 ).
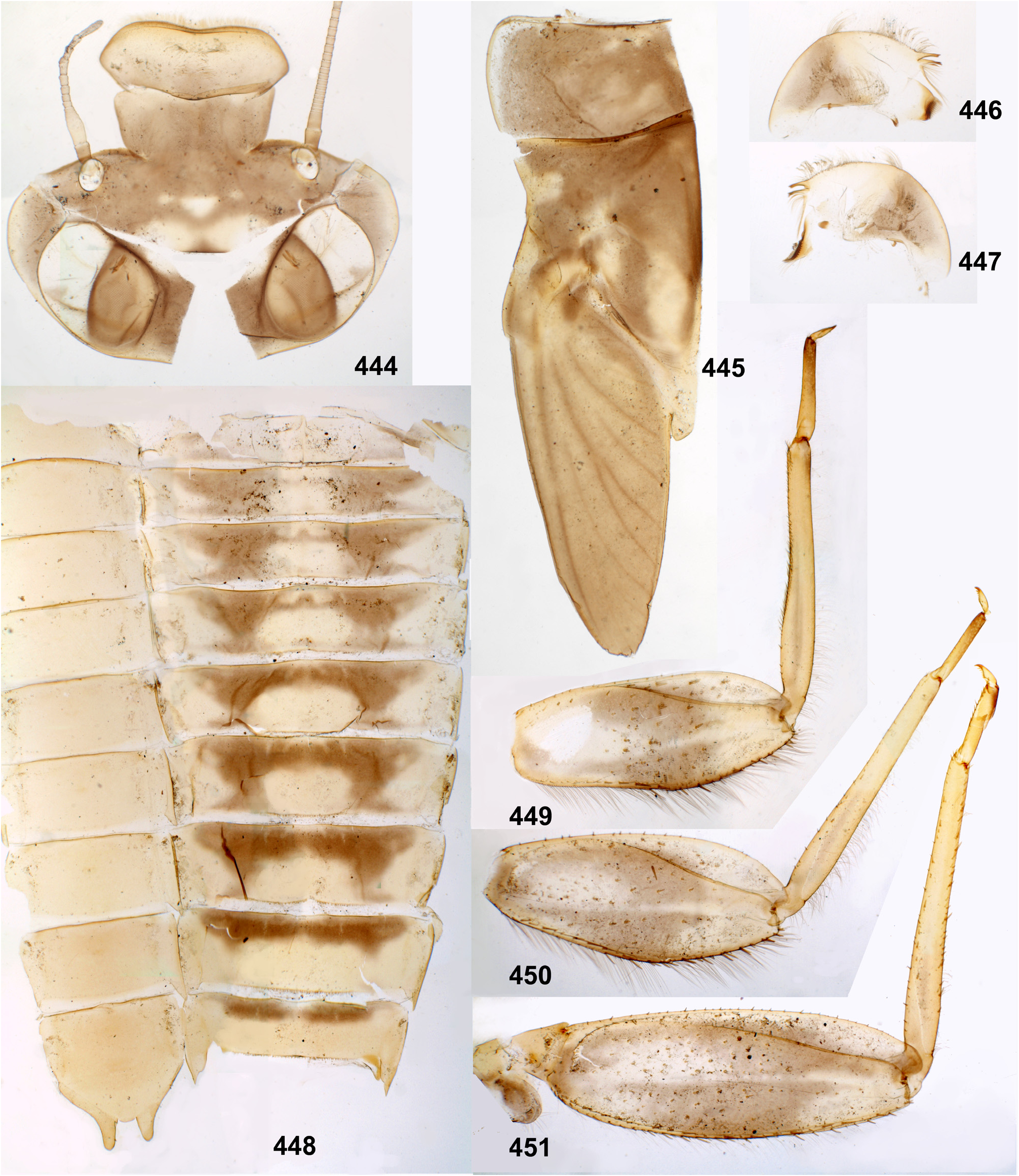
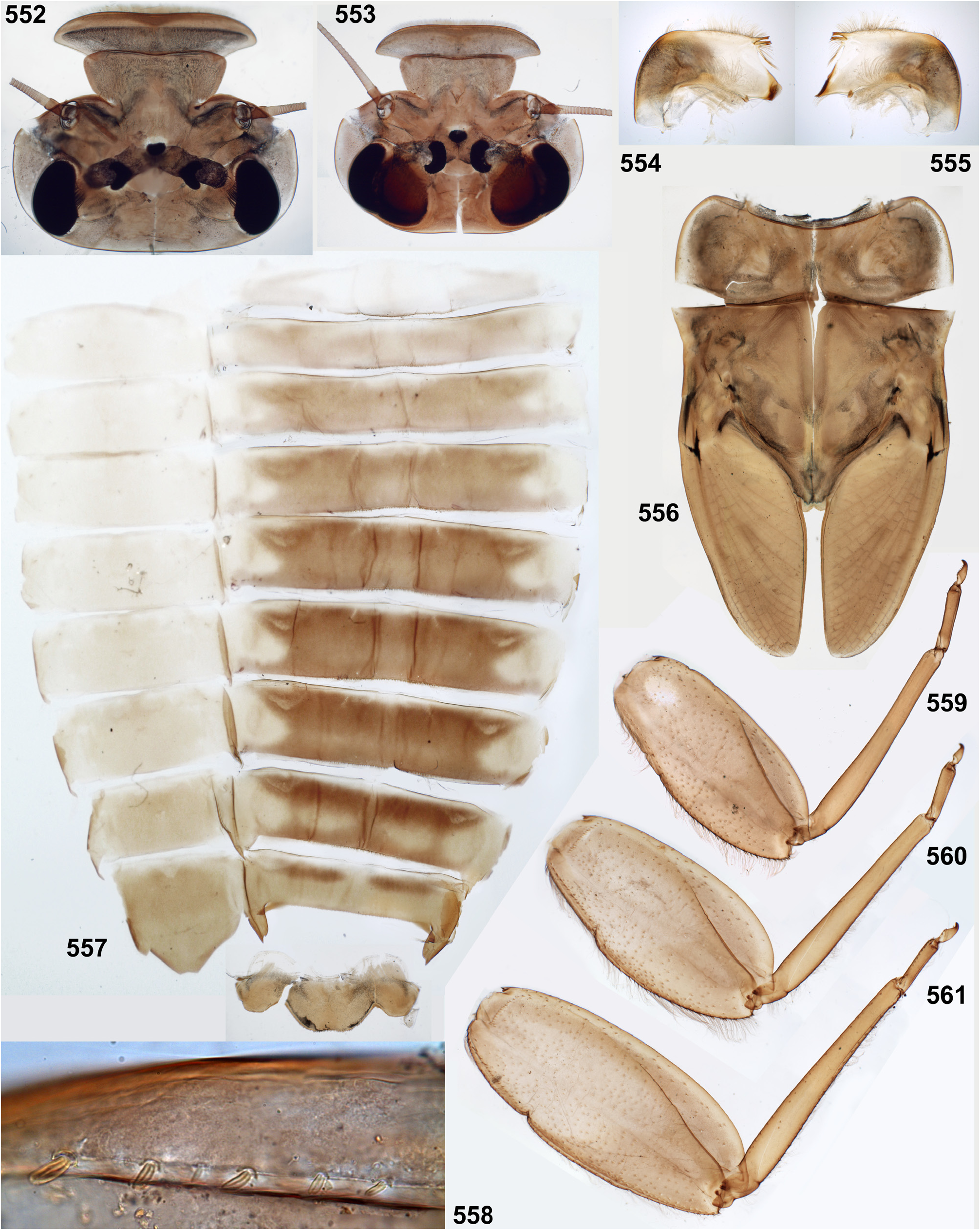
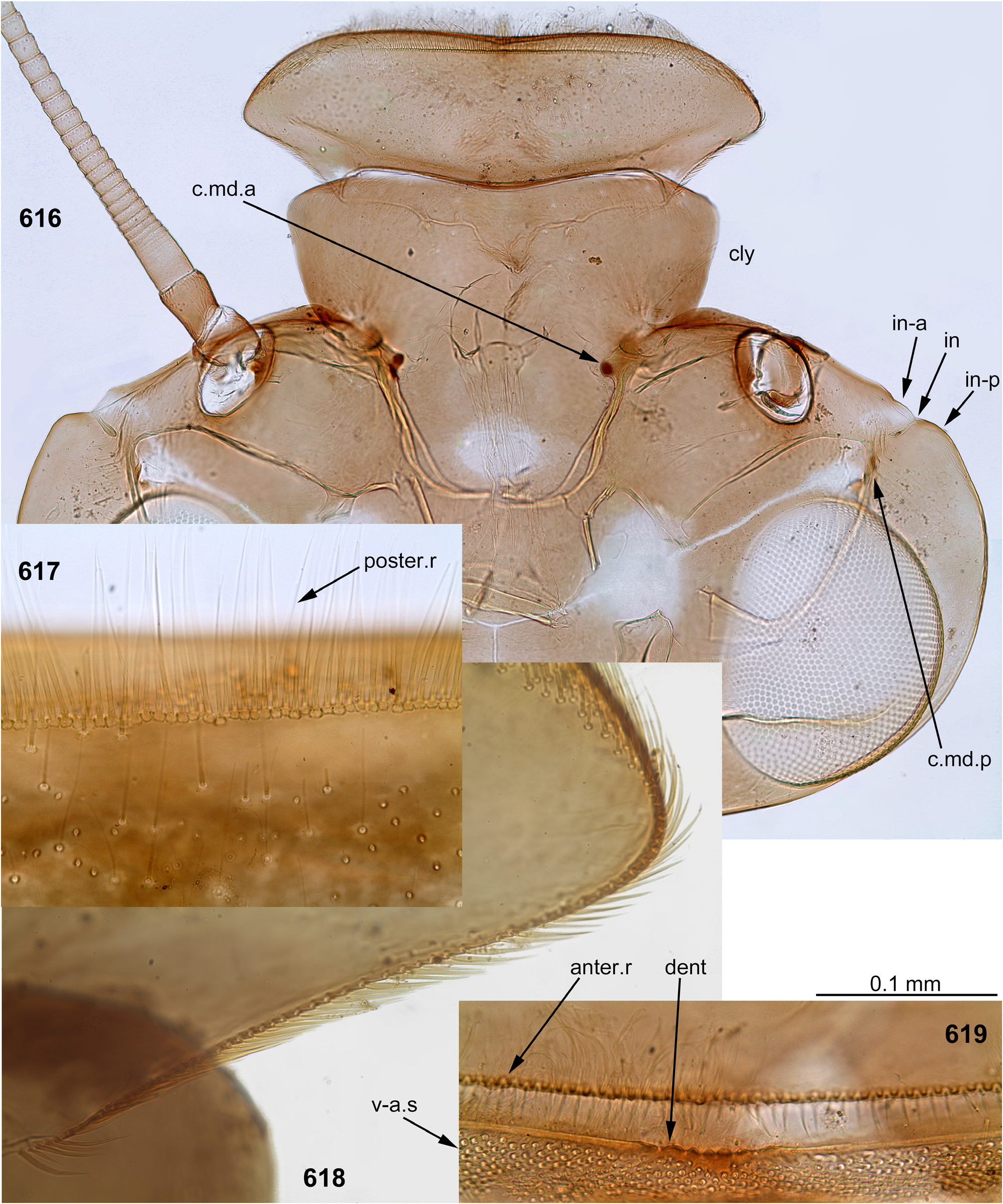
(2) Fronto-clypeal incisions. Lateral margins of the clypeus are separated from lateral margins of the frons by a pair of sharp incisions leading into the anterior tentorial pits; within these incisions, the margins of the frons overlap the margins of the clypeus dorsally ( Figs 3 View FIGURES 1–6 , 14 View FIGURES 7–15 , 68 View FIGURES 68–71 , 105 View FIGURES 101–107 , 235 View FIGURES 229–235 , 335 View FIGURES 332–335 , 389 View FIGURES 387–394 , 444 View FIGURES 444–451 , 485 View FIGURES 476–486 , 552–553 View FIGURES 552–561 , 616 View FIGURES 616–619 ). This differs from many other Atalophlebolinguata, where the frons and the clypeus have an integral dorsal surface, being separated only by a pair of smooth concavities of the lateral margins ( Figs 5–6 View FIGURES 1–6 ) (see below, discussion of systematic position of the Kimminsula -complex).
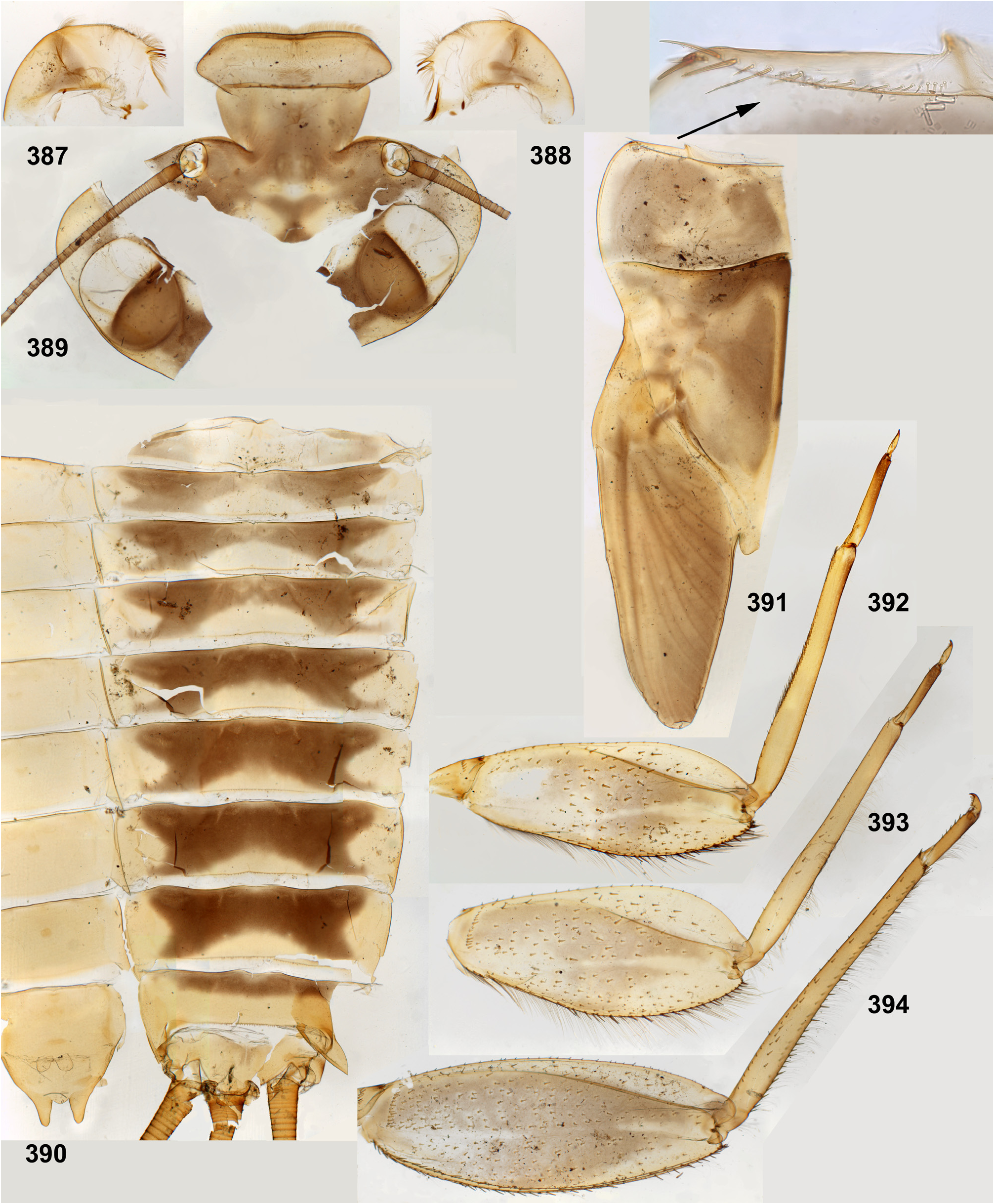
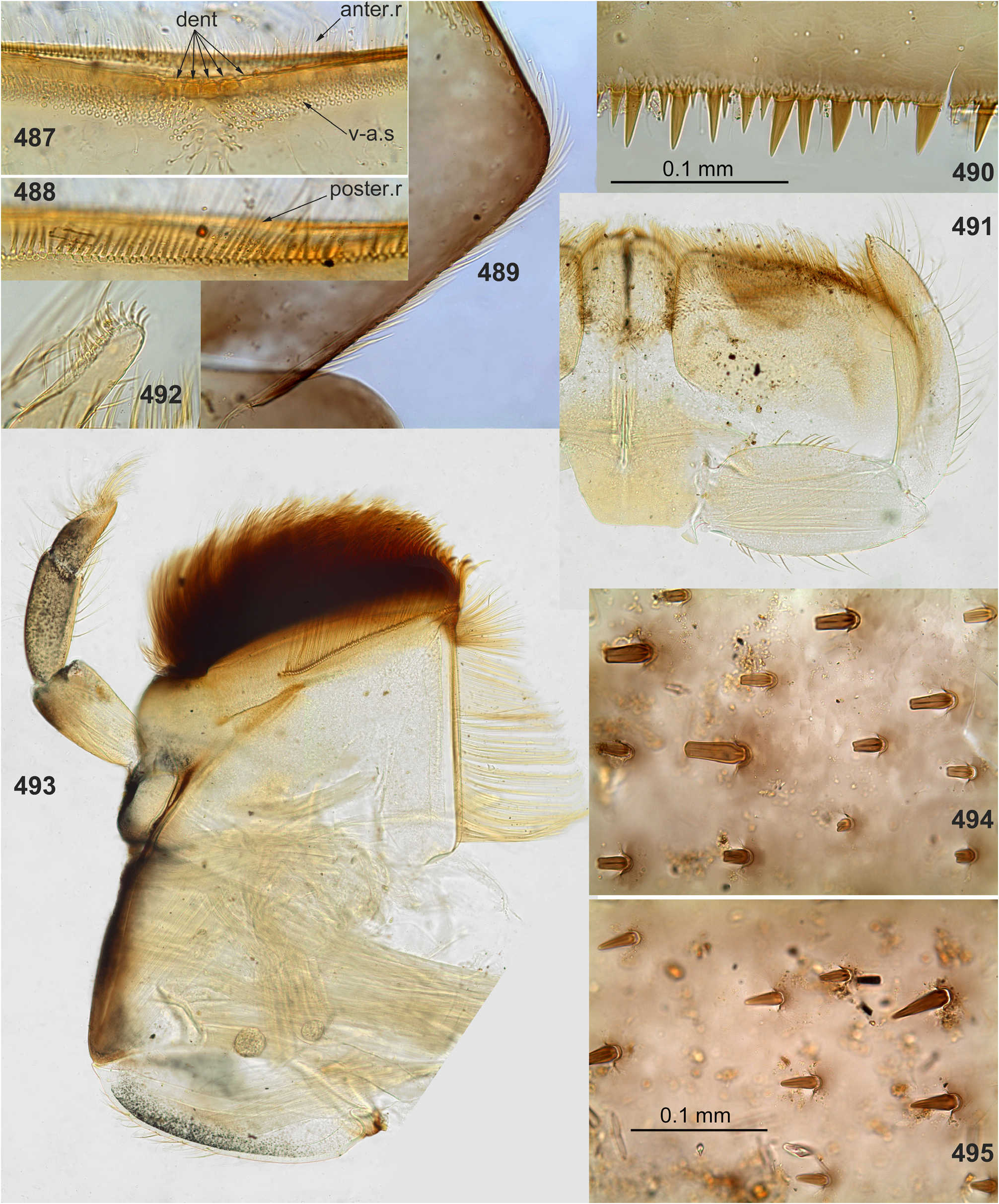
(3) Labrum. As in most other Atalophlebopectinata, the dorsal surface of the labrum bears the posterior transverse setal row and the anterior transverse setal row; in the Kimminsula -complex both rows stretch nearly across the labrum width; each row is dense, either regular, i.e. with one seta per section ( Figs 16 View FIGURES 16–26 , 73 View FIGURES 72–77 , 232 View FIGURES 229–235 : anter.r, 488, 619), or has the form of a strip with a few setae per section ( Figs 102 View FIGURES 101–107 , 232 View FIGURES 229–235 : poster.r, 617). In all species of Kimminsula -complex, the anterior margin of labrum is hooded, i.e. the initial anterior margin is bent ventrally, so that in dorsal view the posterior transverse setal row appears to be located on the visible anterior margin or close to it, while the anterior transverse setal row is often hidden ventrally ( Fig. 101 View FIGURES 101–107 ). Setae of the true ventral side of labrum are situated irregularly (i.e. not forming rows), and form a distinctly outlined transverse ventro-anterior strip close to the anterior margin ( Figs 101, 103 View FIGURES 101–107 ). In all species except Ghatula rufa gen. sp. n., labrum is wider than clypeus. Shape of the median incision is variable: either shallow with more or less developed denticles on the initial anterior margin (in the Ceylonese taxa Ceylonula gen. n., Kimminsula and Hubbardula gen. n., Figs 233 View FIGURES 229–235 , 487 View FIGURES 487–495 , 619 View FIGURES 616–619 ), or cleft-like and lacking denticles (in the Indian taxa Ghatula gen. n. and Petersula , Figs 17 View FIGURES 16–26 , 103 View FIGURES 101–107 ). Petersula differs from other taxa by peculiar labral depressors [see below, Petersula (2)]
(4) Mandibles. As in majority of Atalophlebolinguata, mandibles are articulated at ventral side of the head capsule some distance from its lateral margins ( Fig. 235 View FIGURES 229–235 ); mandibles are flattened, with outer margins convex and projected laterally. The outer margin of mandible bears the distal tuft of setae, which occupies an area adjacent to the base of the incisor, and the middle tuft of setae which is more compact and located more laterally ( Figs 22–23 View FIGURES 16–26 , 75–76 View FIGURES 72–77 , 229 View FIGURES 229–235 , 563 View FIGURES 562–568 , 620 View FIGURES 620–625 ). Among members of the Kimminsula -complex, the middle tuft is absent only in Petersula ( Figs 145–146 View FIGURES 145–153 ). The same in some other leptophlebiid taxa ( Kluge 2020).
(5) Maxillae. Maxilla is wide. The subapical ventral row of comb-like setae (peculiar for Leptophlebiidae ) has more or less expressed curvature, which divides it into two portions: a lateral portion consisting of setae with smaller sockets, and a median portions consisting of setae with larger sockets (length of all setae is equal). In the Ceylonese taxa, i.e. Ceylonula gen. n., Kimminsula and Hubbardula gen. n., the lateral portion is very long and the median portion is short, consisting of 5–10 setae ( Figs 332–334 View FIGURES 332–335 ). In the Indian taxa, i.e. Ghatula gen. n. and Petersula , the whole subapical ventral setal row is shorter, and its both portions are subequal in length ( Figs 112–113 View FIGURES 108–113 ) (the same in many other leptophlebiid taxa).
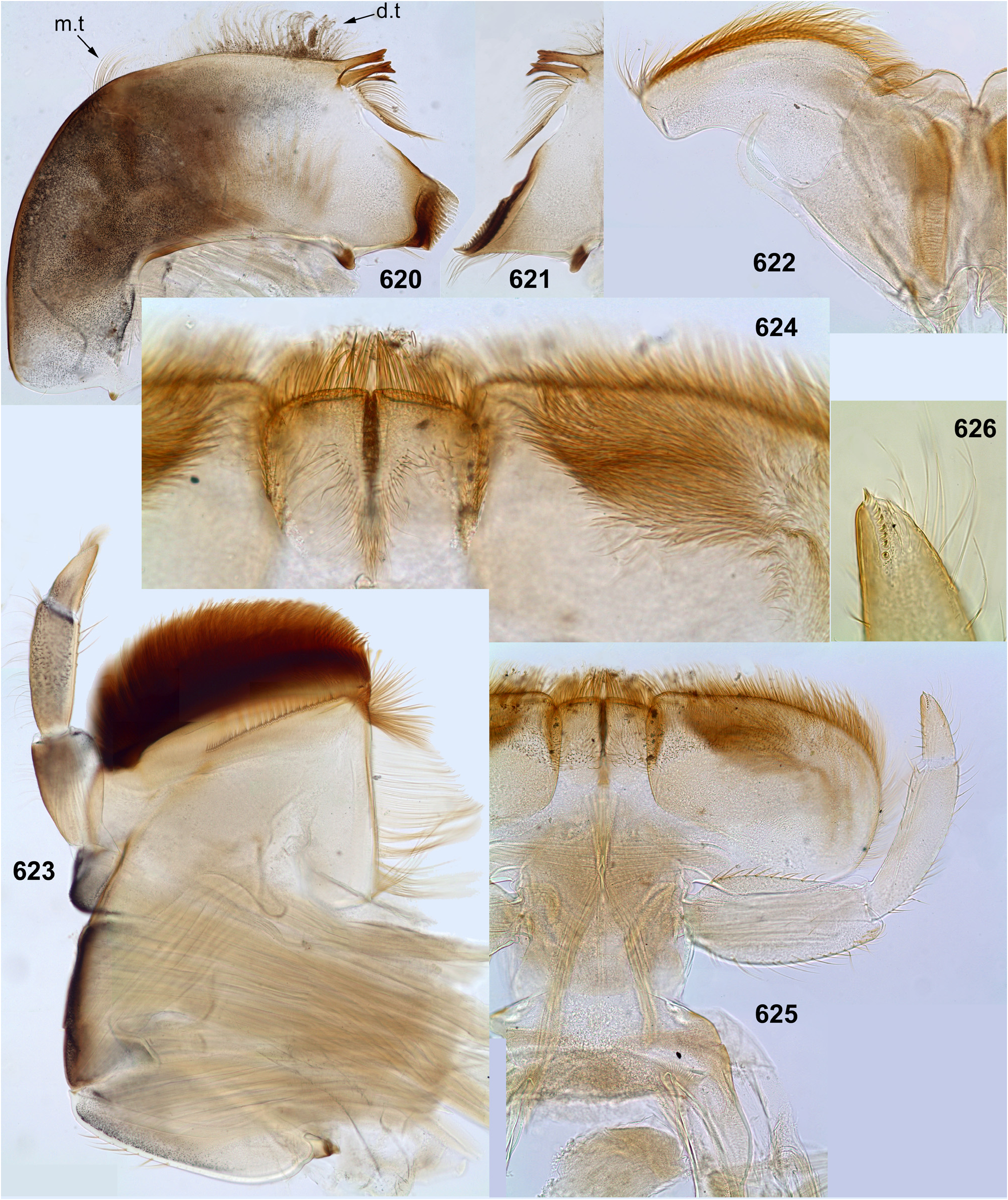
(6) Maxillary palps. Maxillary palp bears long setae on outer sides of the 2nd and 3rd segments, dense setae on ventral side and on apex of the 3rd segment. The inner side of the 3rd segment bears a regular row of setae, whose size and number vary individually and do not provide taxonomic characters within the Kimminsulacomplex. The inner side of the 2nd segment bears longer setae which are either concentrated at its apex (in both species of Petersula , Fig. 111 View FIGURES 108–113 ) or form a more or less long longitudinal row (in other species, Figs 19 View FIGURES 16–26 , 236 View FIGURES 236–242 , 334 View FIGURES 332–335 , 493 View FIGURES 487–495 , 566 View FIGURES 562–568 , 623 View FIGURES 620–625 ).
(7) Labium. Shape and setation of labium is uniform in all representatives ( Figs 26 View FIGURES 16–26 , 77 View FIGURES 72–77 , 116–118 View FIGURES 114–118 , 241 View FIGURES 236–242 , 492 View FIGURES 487–495 , 567 View FIGURES 562–568 , 624–625 View FIGURES 620–625 ). Submentum is bare without prominent setae: either without visible setae at all ( Figs 116 View FIGURES 114–118 , 567 View FIGURES 562–568 , 625 View FIGURES 620–625 ), or with very small occasional setae. The glossae are not large, not projected from paraglossae neither ventrally, nor dorsally; long setae of their ventral side are situated densely and irregularly ( Fig. 625 View FIGURES 620–625 ); long setae on the dorsal side of glossa form irregular row or strip of a horseshoe shape ( Fig. 242 View FIGURES 236–242 , 624 View FIGURES 620–625 ) [as in Thaulodes (Kuge 2020: fig. 20)].
(8) Labial palps. The last (3rd) segment of labial palp is shorter and narrower than others and apically blunt. A small, regular row of stout claw-like setae crosses apex of this segment. In Petersula , these claw-like setae are relatively long ( Fig. 117 View FIGURES 114–118 ); in other taxa they are very short, with length about twice exceeding width at base ( Figs 25 View FIGURES 16–26 , 240 View FIGURES 236–242 , 492 View FIGURES 487–495 , 567 View FIGURES 562–568 , 626 View FIGURES 626–631 ). The same condition occurs in most other Atalophleboculata (see discussion about systematic position below).
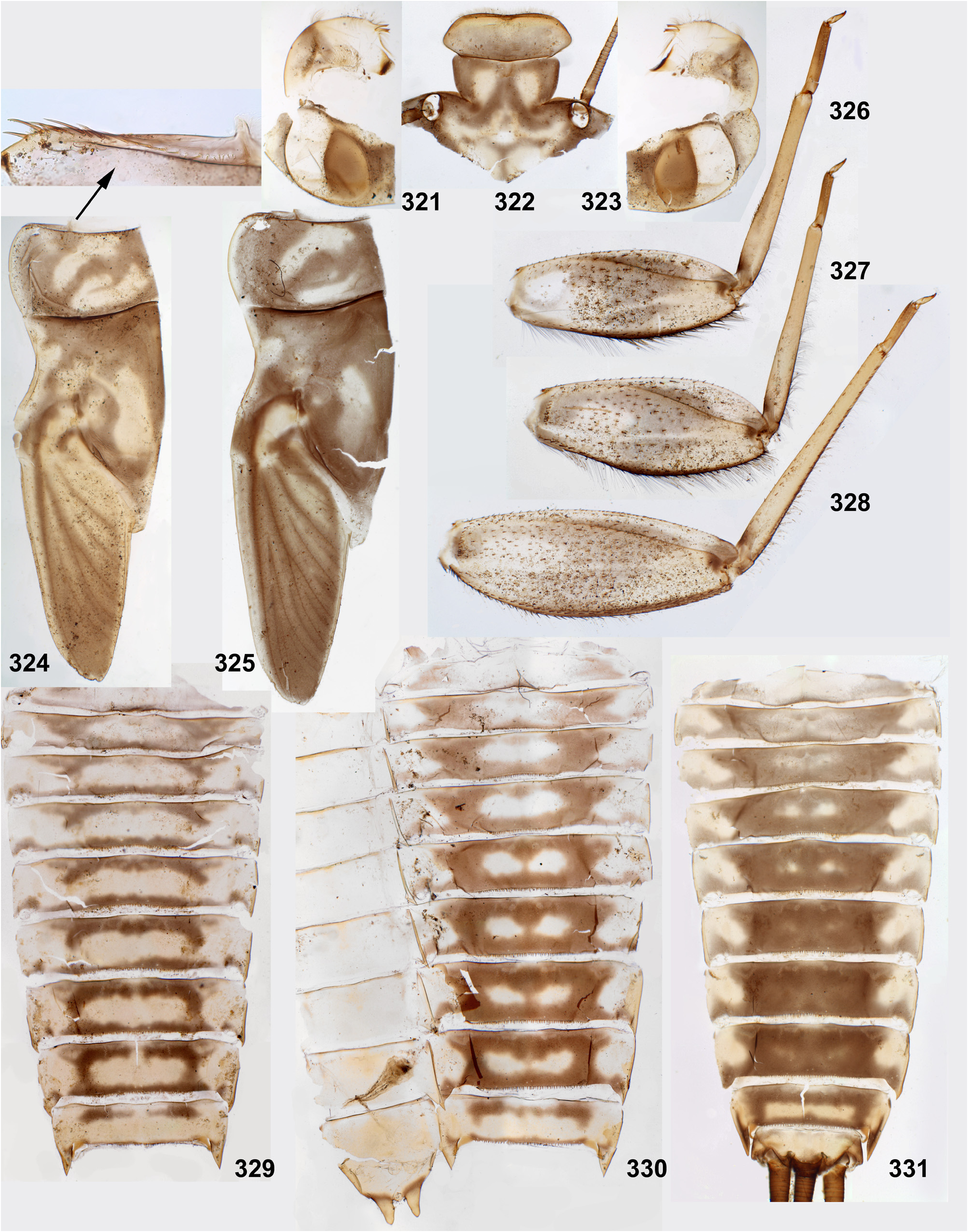
(9) Larval thorax. The pronotum is nearly rectangular, with rounded antero-lateral angles and a free anterior margin (i.e. transverse margin between antero-lateral angles and the neck membrane); a transverse ridge runs parallel to the free anterior margin; humeral setae are located between the free anterior margin and the ridge, being absent in other places ( Figs 9 View FIGURES 7–15 , 91 View FIGURES 85–93 , 95 View FIGURES 94–100 , 228 View FIGURES 221–228 , 324 View FIGURES 321–331 , 391 View FIGURES 387–394 , 481 View FIGURES 476–486 , 556 View FIGURES 552–561 , 610 View FIGURES 607–615 ) (the same in many other Atalophlebolinguata).
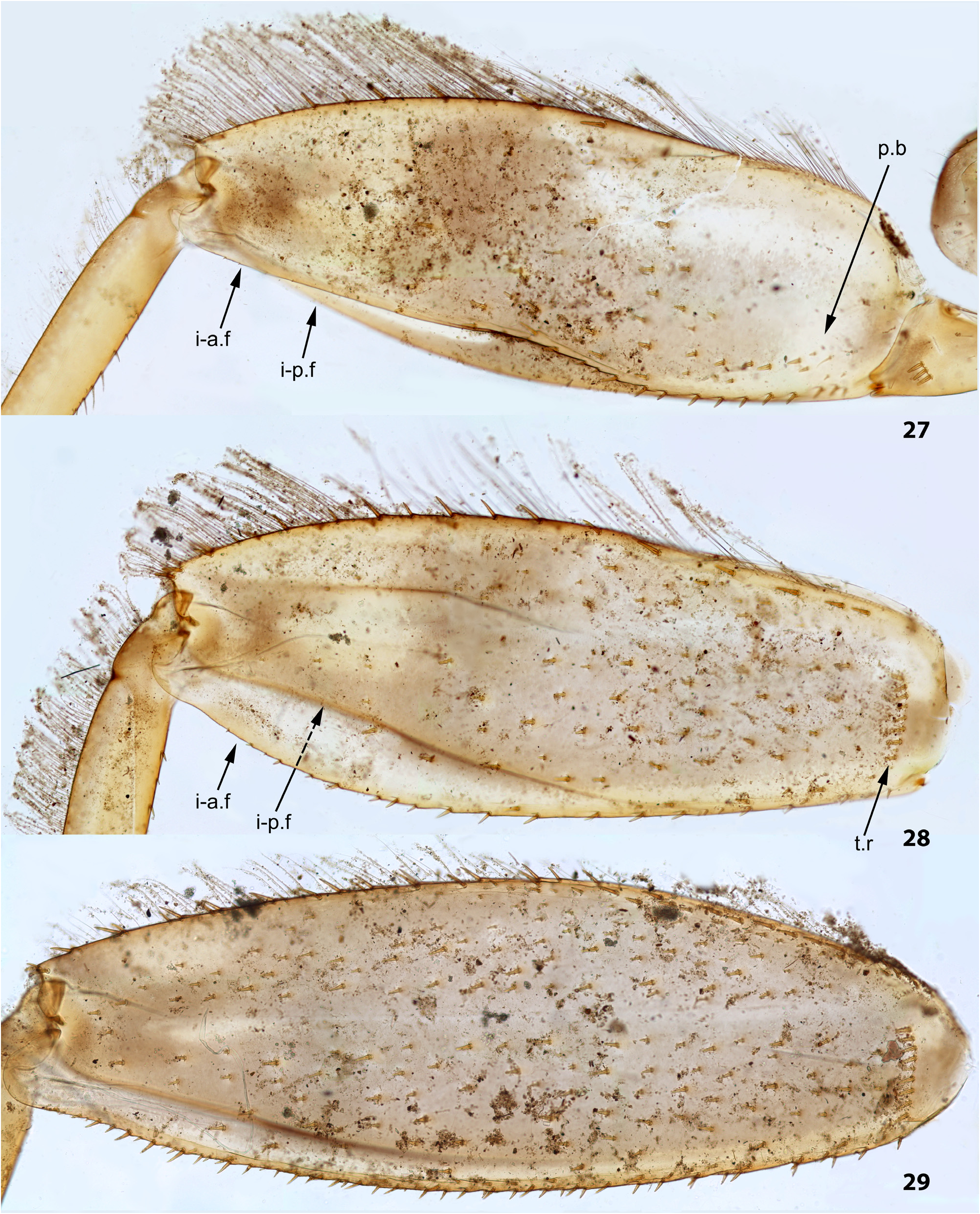
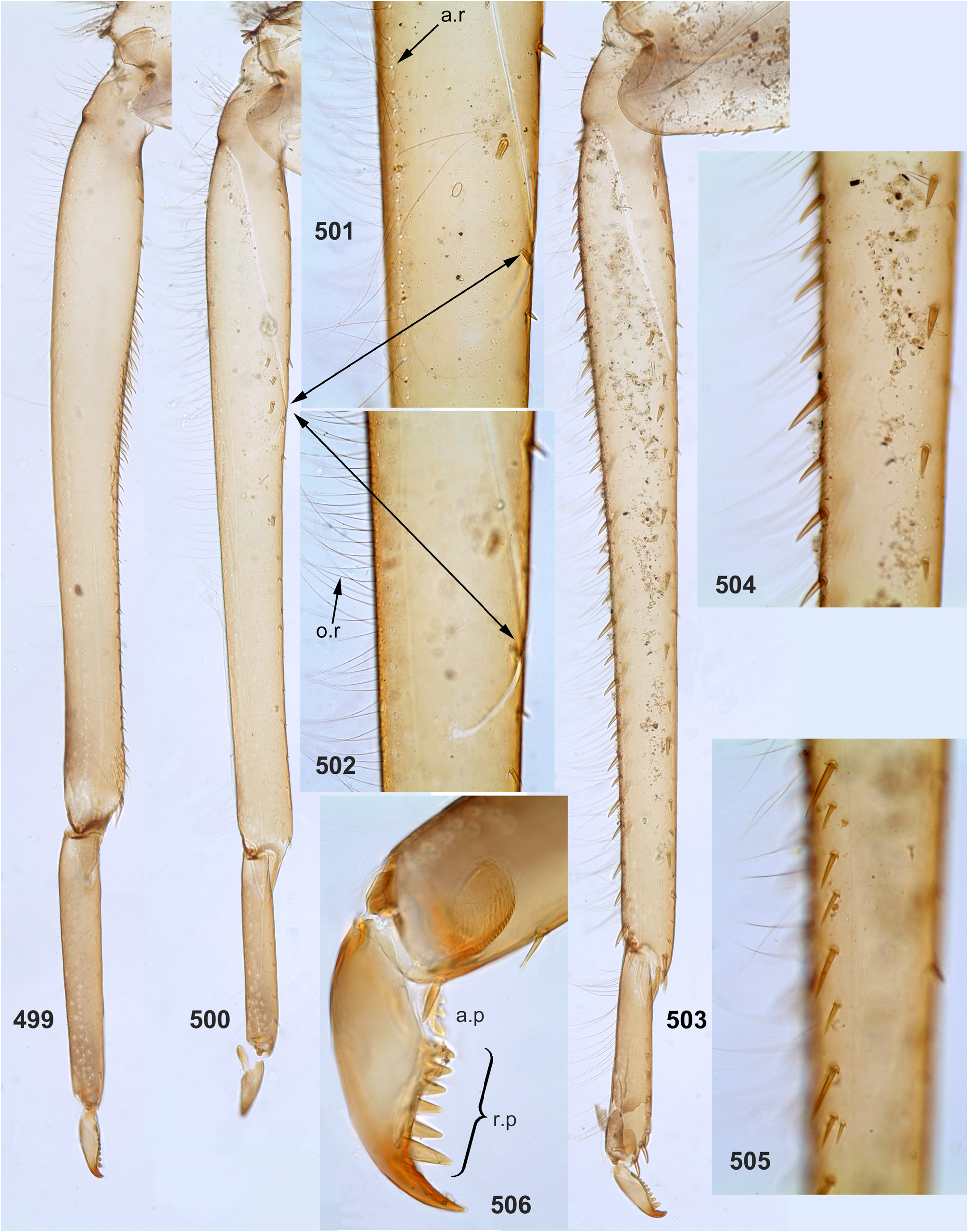
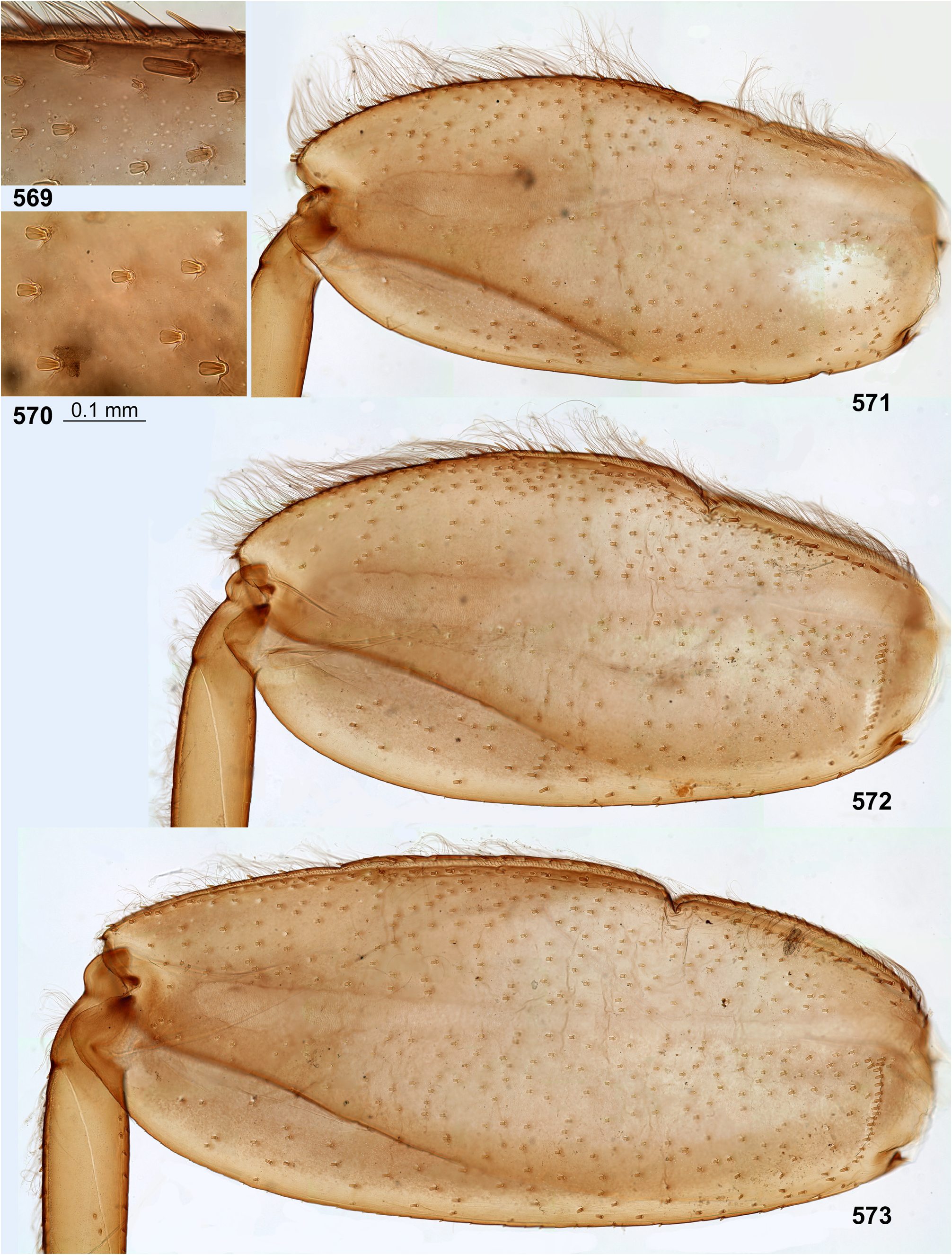
(10) Larval femora. Femora are widened, so the groove on inner side (into which the tibia can be partly inserted) is bordered with prominent flanges (called here the inner-anterior flange and the inner-posterior flange); at least on the middle femur, the inner-anterior flange is expanded in its distal part, so the femur appears widest in distal part ( Fig. 28 View FIGURES 27–29 ); the forefemur has similar shape in all species except Ghatula rufa gen sp. n., in which the inner-anterior flange of forefemur is reduced, being narrower than the inner-posterior flange ( Fig. 27 View FIGURES 27–29 ); the hind femur retains the usual elongate-ellipsoid shape, being widest in the middle ( Fig. 29 View FIGURES 27–29 ).
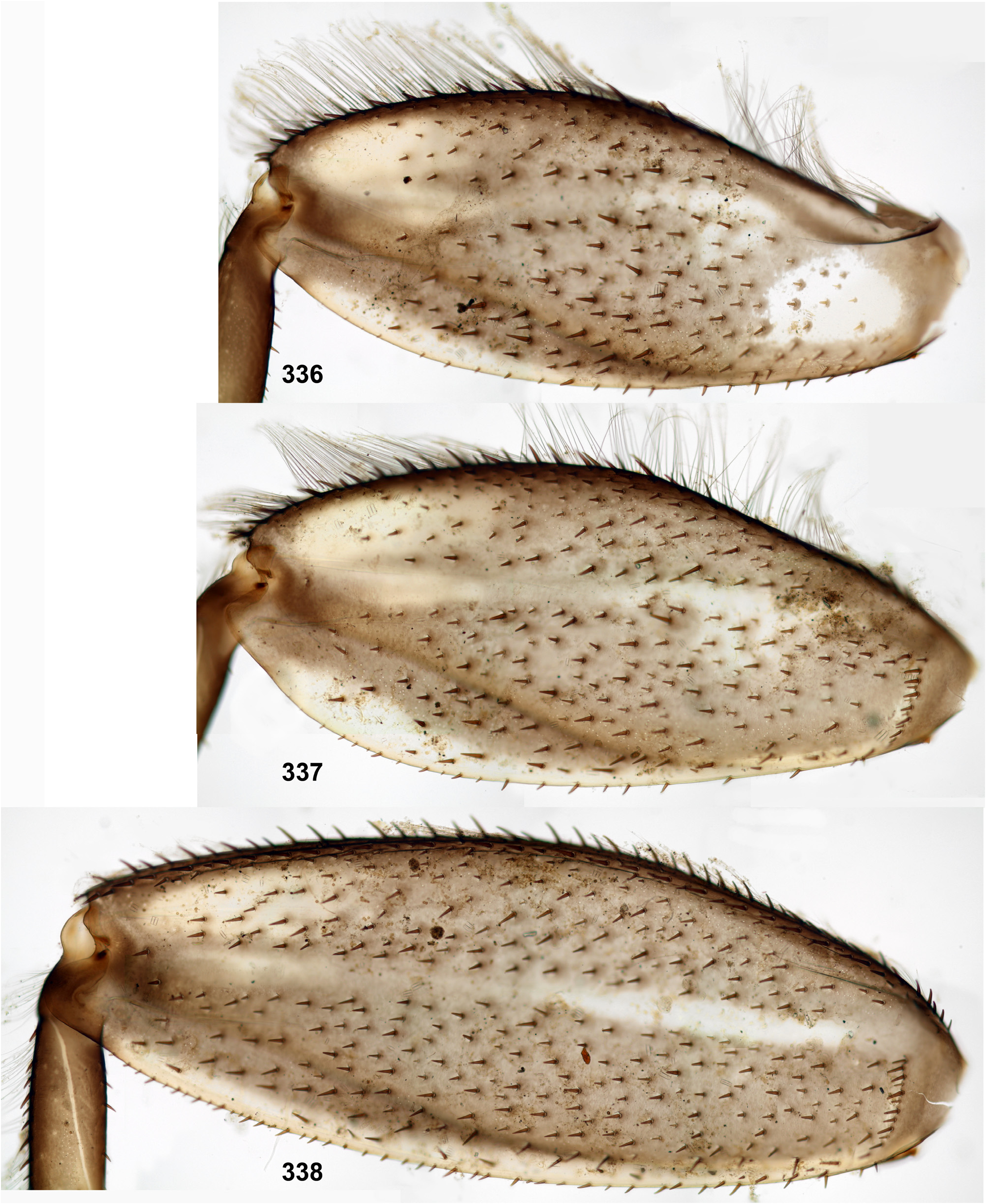
The forefemur is not widened proximally (in contrast to many other Leptophlebiidae ), but the cuticle of its anterior surface bears the proximal blank peculiar for mayfly larvae whose forefemur is proximally wider ( Kluge 2020: p. 15). The proximal blank of foreleg differs from blanks occurring on the middle and hind legs ( Fig 27 View FIGURES 27–29 , 119 View FIGURES 119–123 , 243 View FIGURES 243–250 , 336 View FIGURES 336–338 , 392 View FIGURES 387–394 , 449 View FIGURES 444–451 , 496 View FIGURES 496–498 , 571 View FIGURES 569–573 , 626 View FIGURES 626–631 ).
The anterior surface of each femur bears more or less numerous, irregularly situated, stout setae, which can be either blunt ( Fig. 494 View FIGURES 487–495 ), or pointed ( Fig. 495 View FIGURES 487–495 ), or variable. On the inner side of the femur the stout setae form a more or less regular row running by margin of the inner-anterior flange. On middle and hind femora (but never on forefemur) the stout setae form a regular, arched, transverse row situated near the femur base ( Figs 28–29 View FIGURES 27–29 , 79–80 View FIGURES 78–80 , 120–121 View FIGURES 119–123 , 245 View FIGURES 243–250 , 337–338 View FIGURES 336–338 , 497–498 View FIGURES 496–498 , 572–573 View FIGURES 569–573 , 627 View FIGURES 626–631 ). On outer side of each femur the stout setae are enlarged and form two more or less regular rows (which can be called the outer-anterior row and the outer-posterior row), between which the outer strip of thin hairs is located; hairs of this strip are situated densely and irregularly ( Fig. 629 View FIGURES 626–631 ). Hairs of the outer strip are generally longer and/or denser on the fore- and the middle femora, than on the hind femur ( Figs 27–29 View FIGURES 27–29 , 78–80 View FIGURES 78–80 , 119–121 View FIGURES 119–123 , 336–338 View FIGURES 336–338 , 392–394 View FIGURES 387–394 , 449–451 View FIGURES 444–451 , 496– 498 View FIGURES 496–498 , 571–573 View FIGURES 569–573 , 612–614 View FIGURES 607–615 ); in Ceylonula gen. n. they are completely absent on the hind femur ( Fig. 245 View FIGURES 243–250 ). The posterior side of the hind femur bears curved setae —these are small, pointed, usually pectinate setae, curved toward the inner margin of the femur and forming an irregular strip along inner margin of the femur ( Figs 36 View FIGURES 36–42 , 123 View FIGURES 119–123 , 247 View FIGURES 243–250 ).
The curved setae are probably initial for Leptophlebiidae ; other features of femur setation, especially the transverse proximal rows on middle and hind femora, distinguish the Kimminsula -complex from many other Leptophlebiidae (see below, discussion about systematic position of the Kimminsula -complex).
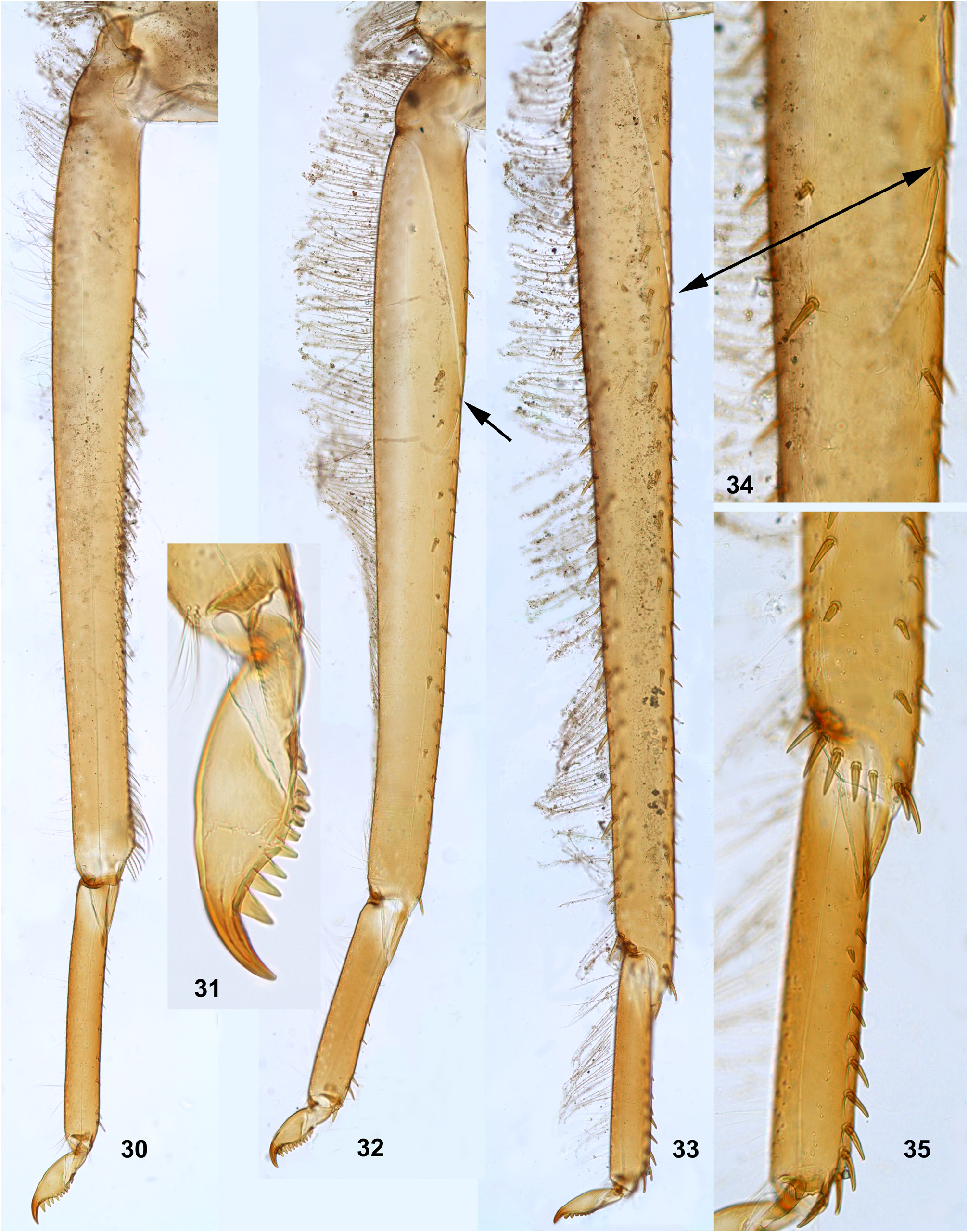
(11) Larval tibiae. Patella-tibial suture, which initially for mayflies is absent on the forelegs and present on the middle and the hind legs ( Kluge 2004), retains this condition in Ghatula gen. n., Ceylonula gen. n. and Hubbardula gen.n.; it is differently modified in Petersula and in Kimminsula [see below, Petersula (7) and Kimminsula (7)].
As in most other Leptophlebiidae , tibiae of three leg pairs are differentiated: the pointed pectinate setae of inner side are preferably developed on the foreleg, and the stout setae of outer side are preferably developed on the hind leg.
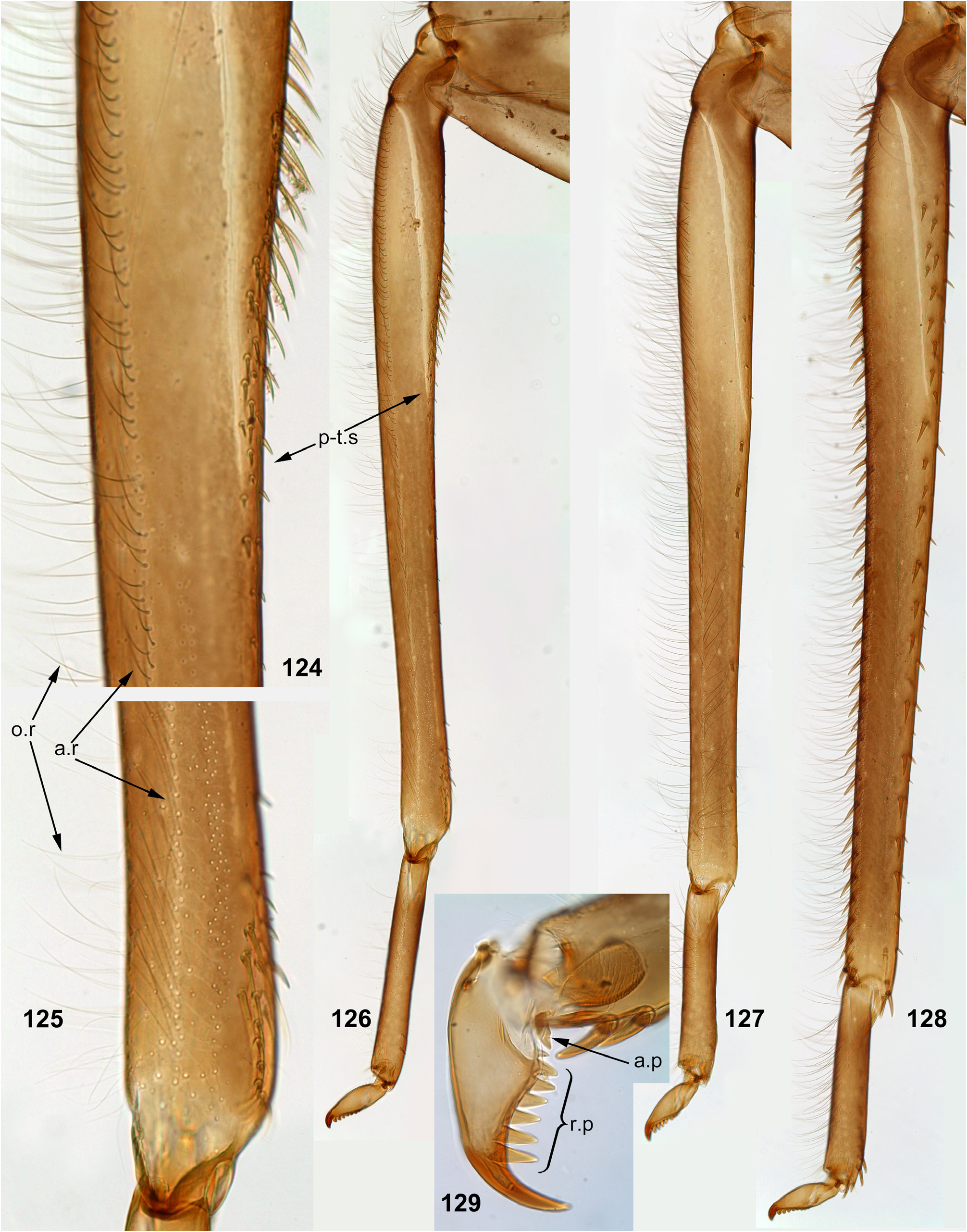
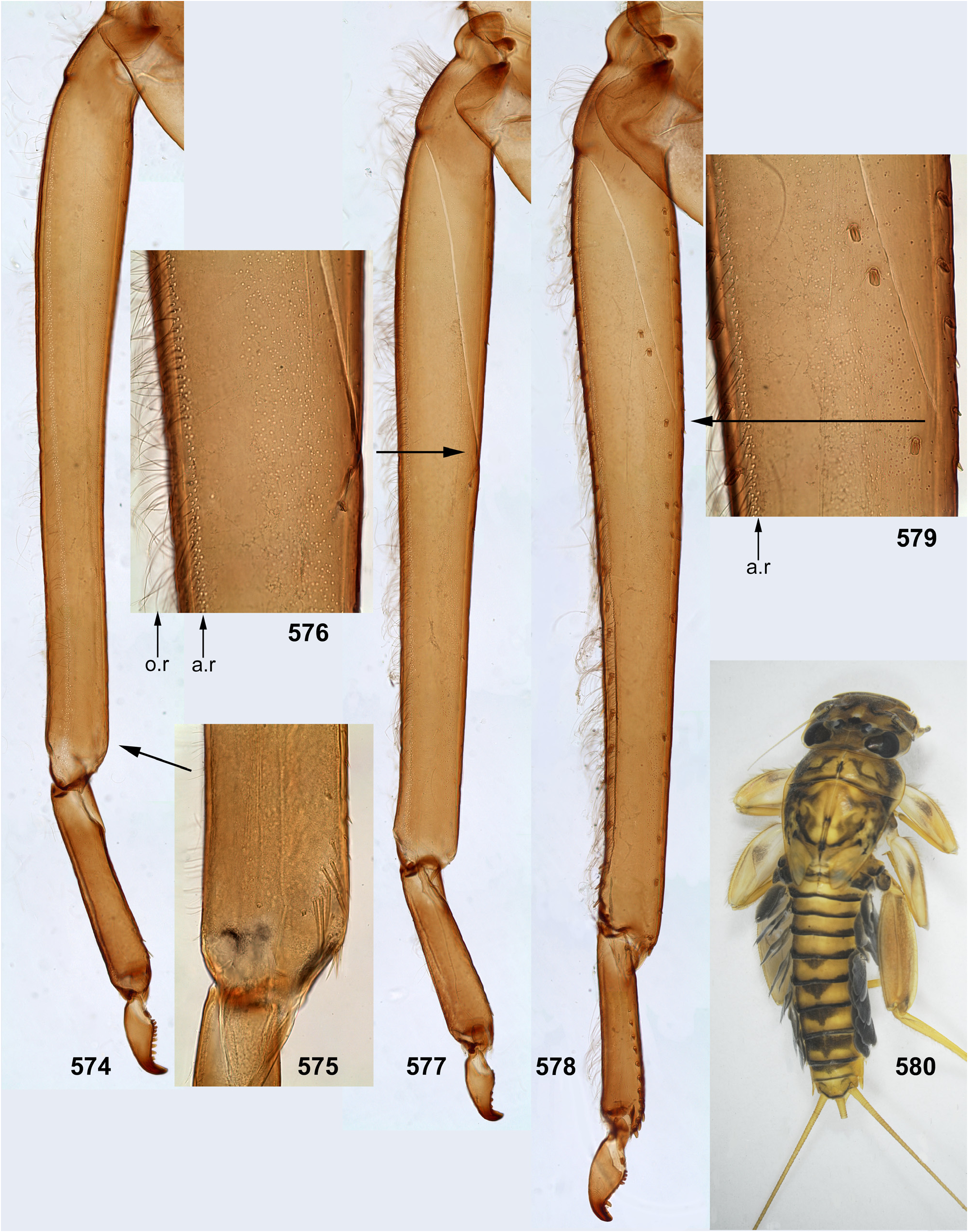
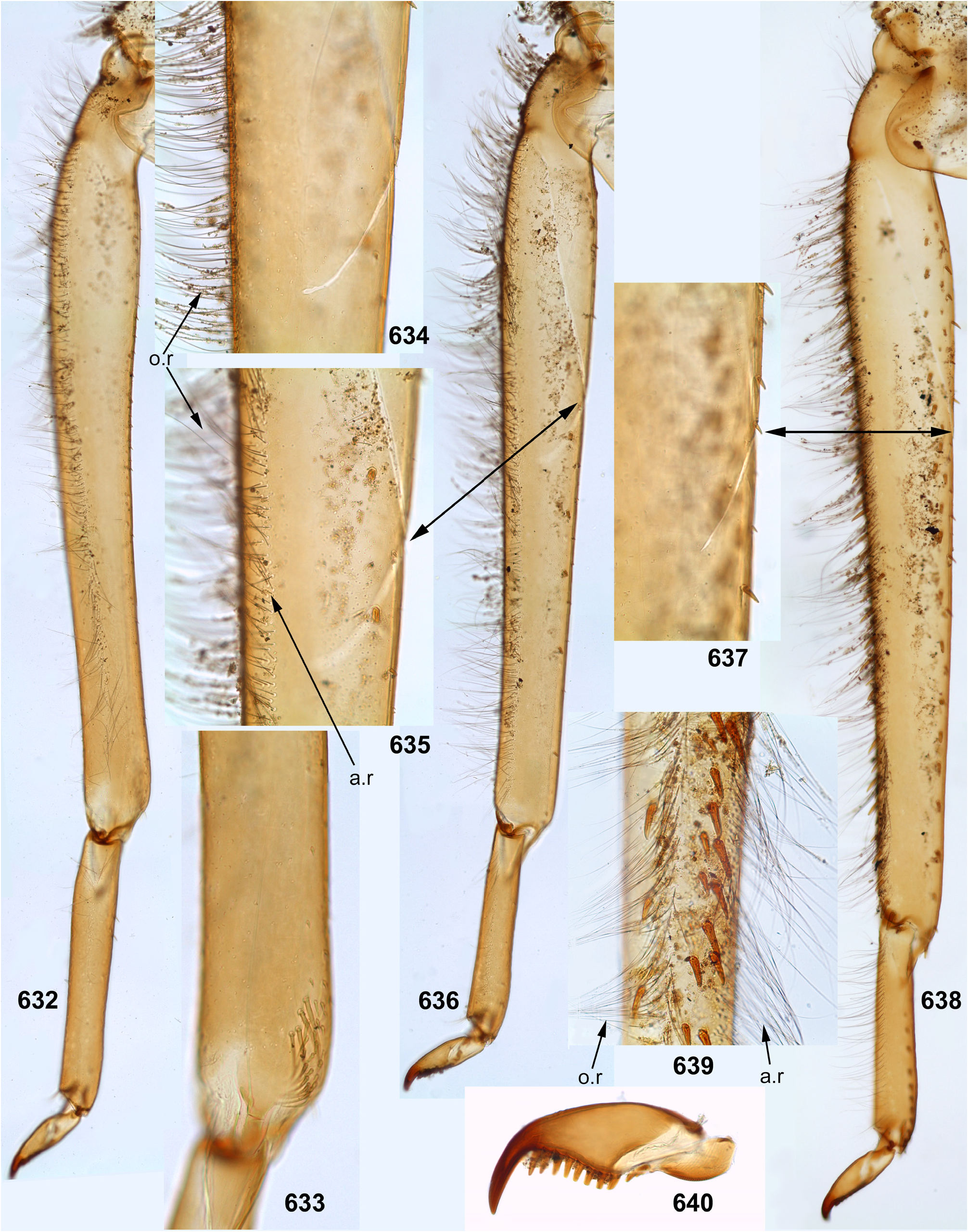
The foretibia bears stout pointed setae, which are often bipectinate; these setae occupy a longer or shorter part of the inner side of the foretibia, being always present at least near the apex of the tibia ( Figs 124–126 View FIGURES 124–129 , 253–254 View FIGURES 251–260 , 339–340 View FIGURES 339–347 , 574–575 View FIGURES 574–580 , 632–633 View FIGURES 632–640 ); on the middle tibia such setae are present only in Ceylonula gen. n. ( Figs 255–256 View FIGURES 251–260 ); on the hind tibia such setae are absent in all representatives of the Kimminsula -complex.
The hind tibia differs from other tibiae by the presence of stout setae located on all sides, forming two longitudinal rows on outer side of the tibia—the outer-posterior and the outer-anterior ones ( Figs 258 View FIGURES 251–260 , 504–505 View FIGURES 499–506 , 639 View FIGURES 632–640 ) and forming a transverse row on posterior side at the tibia apex ( Figs 35 View FIGURES 30–35 , 259 View FIGURES 251–260 ).
As in many other Leptophlebiidae , the outer side of each tibia bears a longitudinal row of hairs (long and slender setae). In Ghatula gen. n. and Ceylonula gen. n. this row of hairs is single, but Petersula , Kimminsula and Hubbardula gen. n. have a second row located more anteriorly. Each of these rows is either regular, i.e. with one seta per section ( Figs 124–125 View FIGURES 124–129 , 342 View FIGURES 339–347 , 501 View FIGURES 499–506 ), or has a form of strip with a few setae per section ( Figs 576 View FIGURES 574–580 , 639 View FIGURES 632–640 ). On the hind tibia, which has two rows of stout setae on the outer side, the initial outer row of hairs is located between them, and the additional anterior row of hairs is located anteriad of the outer-anterior row of stout setae ( Fig. 639 View FIGURES 632–640 ).
(12) Larval tarsus. The tarsus of each leg pair bears hairs on the outer side similar to hairs on the tibia; these hairs are longer and denser on the hind leg. Stout setae on inner sides of tarsi are differentiated as follows: on the fore- and middle legs they are small and few, can be absent; on the hind leg stout setae always form a longitudinal row at least on the distal part of the tarsus; in proximal part of the row setae are sparse and small, but they become denser, larger and hooked toward apex of the tarsus ( Fig. 35 View FIGURES 30–35 ). Sometimes inner side of the foretarsus bears several stout, pointed, pectinate setae of the same structure as the setae on inner side of the tibia of the same leg; such setae occur in Ceylonula femoralis ( Figs 251, 253 View FIGURES 251–260 ) and some others.
(13) Larval claw. Claw is bent perpendicular to the leg flatness and distinctly divided into the small articulatory portion and the large rigid portion ( Figs 129 View FIGURES 124–129 , 506 View FIGURES 499–506 ) ( Kluge 2020).
(14) Larval abdomen. The surface of terga and sterna is always smooth, without prominent relief or significant setae. Posterior margin of each abdominal tergum bears a row of sharply pointed denticles greatly varying in size; medium size of denticles is either subequal on all terga I–X ( Figs 38–41 View FIGURES 36–42 ), or significantly differs on different terga. Posterior margin of the 10th tergum always has shallow median concavity ( Figs 41 View FIGURES 36–42 , 398 View FIGURES 395–405 , 586 View FIGURES 581–586 , 631 View FIGURES 626–631 ). Posterior margins of all sterna I–IX are always smooth, without denticles ( Fig. 94 View FIGURES 94–100 ).
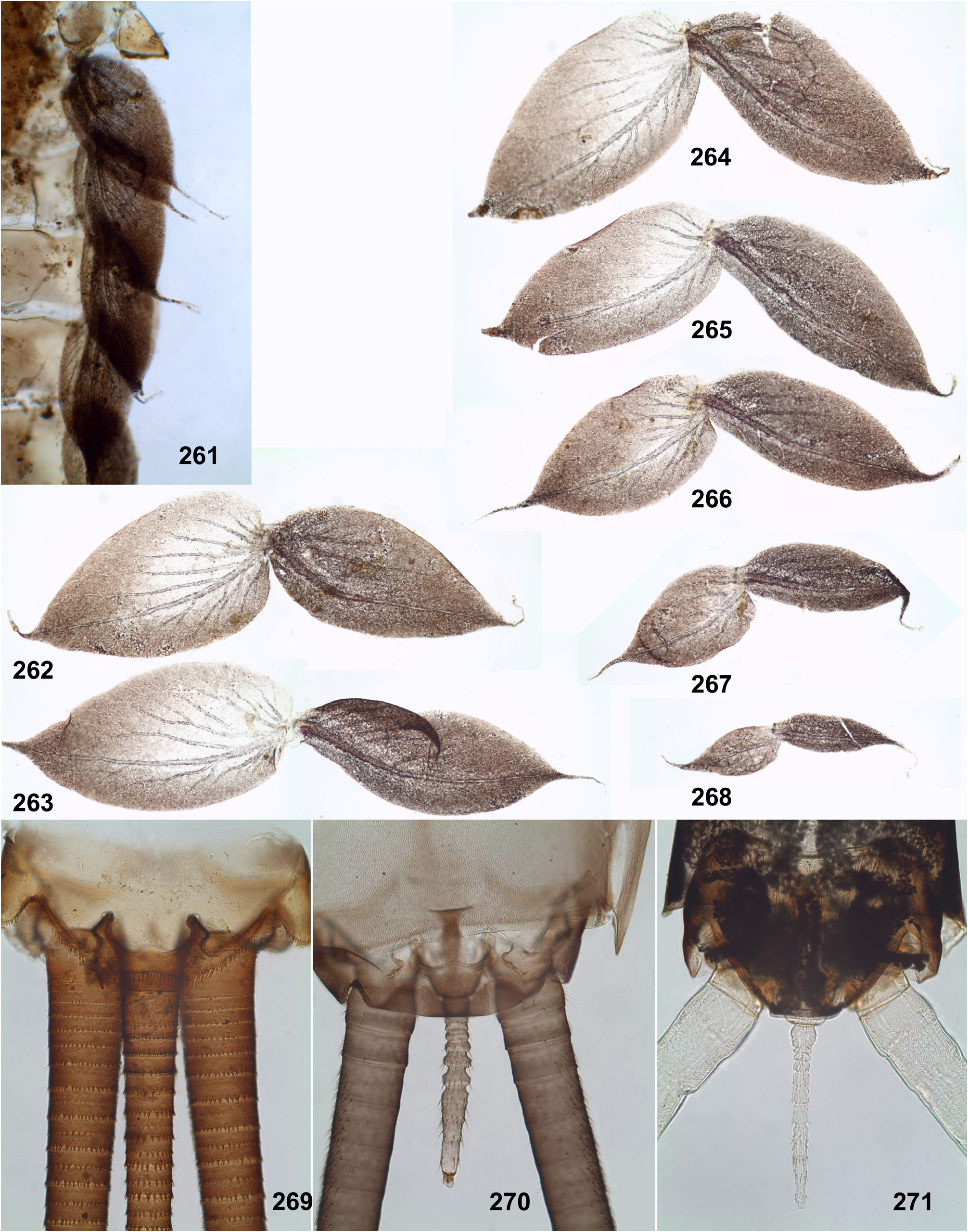
Caudalii are about 2–3 times longer than the body; all 3 caudalii (the cerci and the paracercus) are equally developed both in Ceylonula femoralis (whose paracercus is vestigial in the winged stages, Figs 269–271 View FIGURES 261–271 ) and in other species (where the paracercus is equal to the cerci in all stages); posterior margin of each segment of cercus and paracercus bears whorls of pointed denticles and setae in all species ( Figs 41–42 View FIGURES 36–42 ).
(15) Tergalii. Tergalii always retain 2 lamellae (initial for Leptophlebiidae ); tergalii of all pairs have similar structure, with both lamellae similar. Tergalii make rhythmic undulate movements with the wave going from front to back; either all pairs of tergalii participate equally in respiration ( Figs 440–442 View FIGURES 440–443 ) or tergalii I do not participate in respiratory movement ( Fig. 202 View FIGURES 196–204 ). In other respects, structure of tergalii significantly differs in different taxa [see below, Ghatula (9), Petersula (9), Ceylonula (9), Kimminsula (9) and Hubbardula (9)]. All 7 pairs of tergalii are retained in Petersula , Ceylonula gen. n., Kimminsula and Hubbardula gen. n., while Ghatula gen. n. has no tergalii of 7th pair, thus retains only 6 pairs of tergalii.
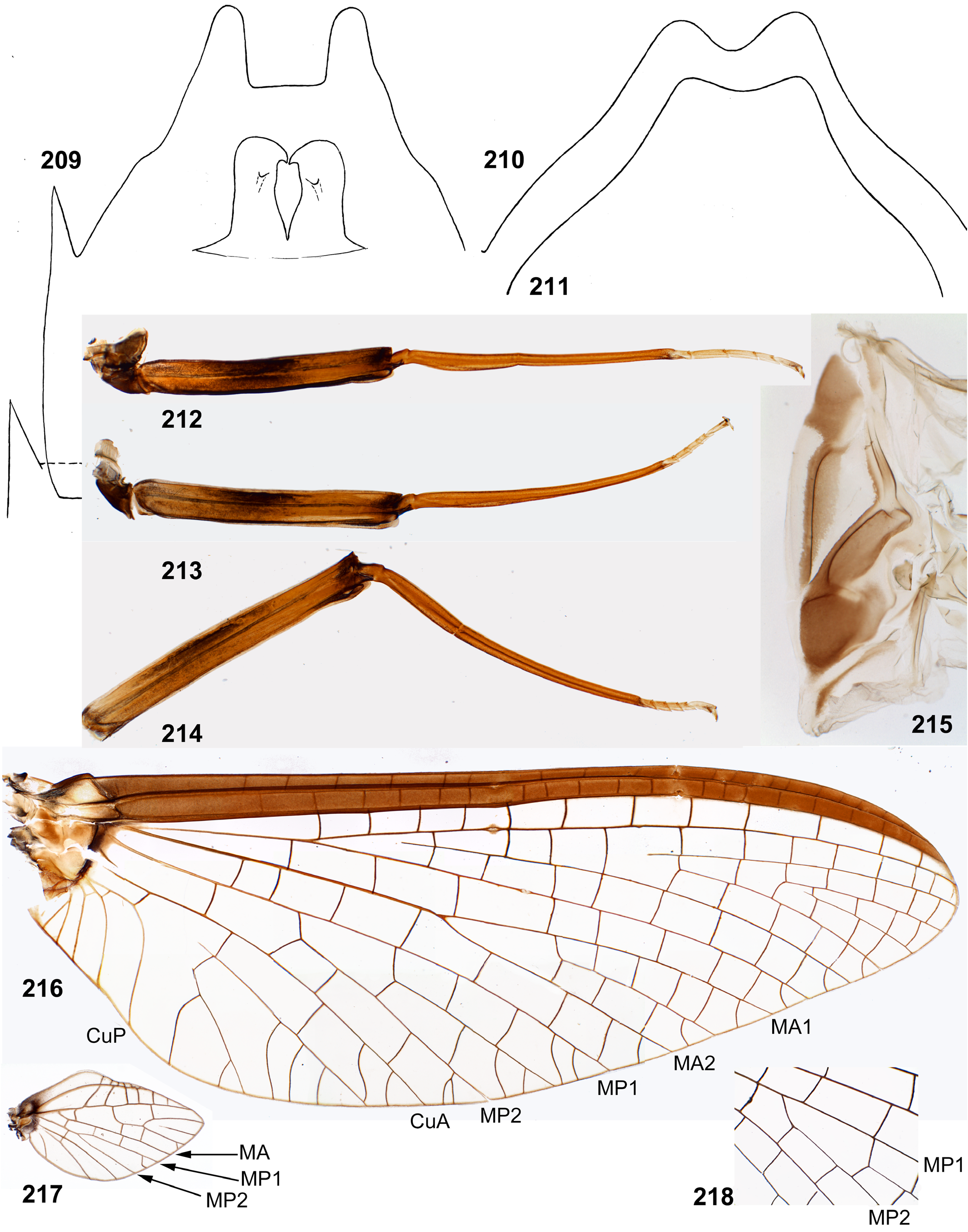
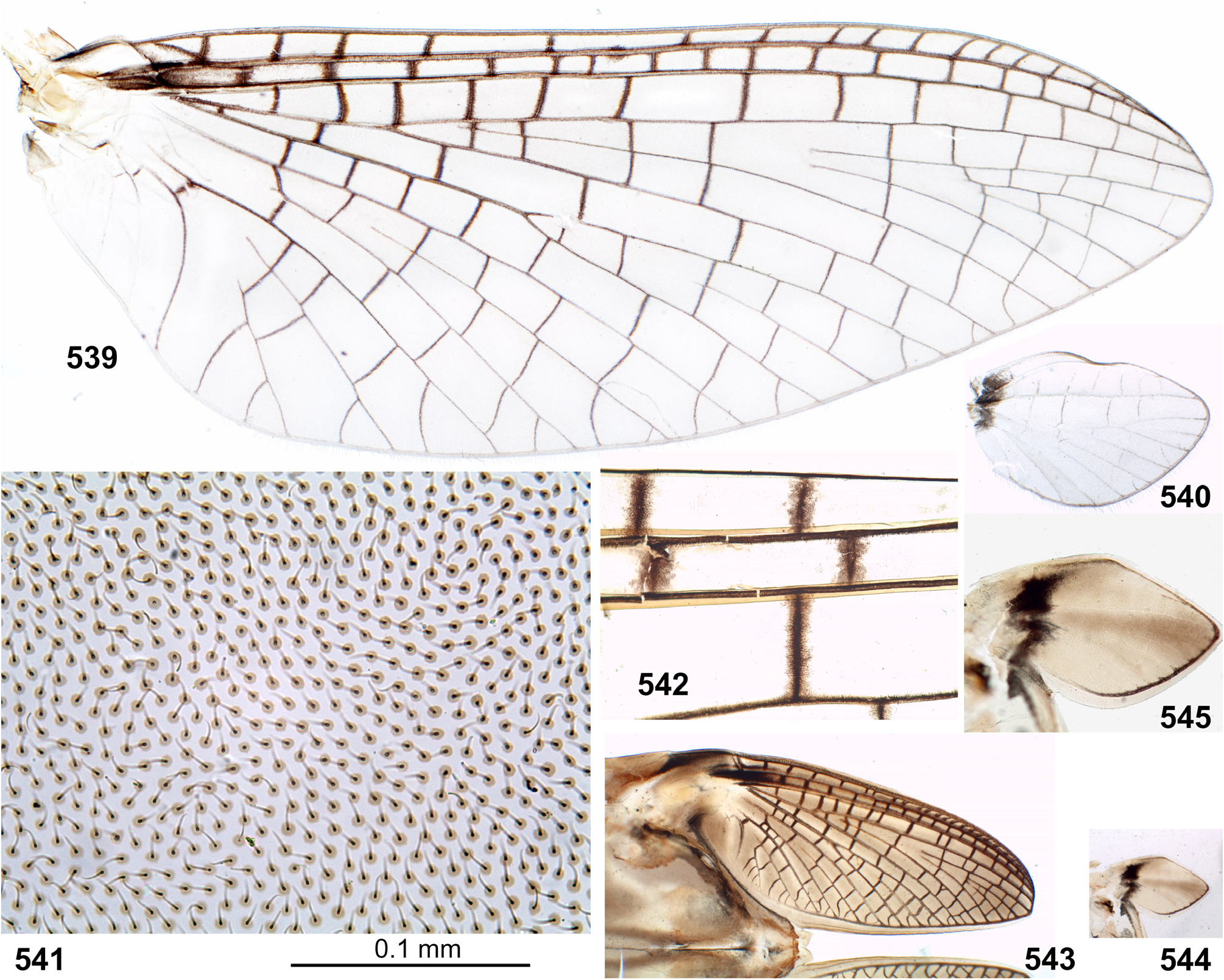
(16) Forewings. Furcation of MA is symmetrical or slightly asymmetrical. Furcation of MP is more proximal than furcation of RS and is sharply asymmetrical: vein MP 2 is attached at base to veins MP 1 and CuA either by equally oblique crossveins ( Fig. 411 View FIGURES 406–412 ), or the crossvein connecting MP 2 with MP 1 is more oblique and may be interpreted as asymmetric arising of MP 2 from MP 1 ( Figs 51 View FIGURES 51–58 , 68 View FIGURES 68–71 , 151 View FIGURES 145–153 , 156 View FIGURES 154–165 , 279 View FIGURES 278–289 , 453 View FIGURES 452–457 , 539 View FIGURES 539–544 , 670 View FIGURES 664–672 ); this difference may be individual ( Figs 216, 218 View FIGURES 209–218 ). Cubital field has 2 intercalaries between Cu 1 and Cu 2; in Kimminsula latifolia sp. n. a third, additional intercalary is present between them ( Fig. 581 View FIGURES 581–586 ). Crossveins are numerous; some crossveins reach posterior margin of the wing between ends of the longitudinal and the intercalary veins. Crossveins in the area of pterostigma are numerous, complete and non-anastomosed; in Ceylonula femoralis the costal field is wider than the subcostal field, and the pterostigmatic crossveins are nearly perpendicular to Sc ( Fig. 279 View FIGURES 278–289 ); in other taxa the costal field is not wider than the subcostal field, and the pterostigmatic crossveins vary from sharply oblique ( Fig. 414 View FIGURES 413–416 ) to perpendicular to Sc ( Figs 670 View FIGURES 664–672 ).
(17) Hind wings. Forewing has well expressed tornus, and the hind wing is relatively large, about as long as the basi-tornal margin of the forewing. Costal projection of the hind wing is blunt and slightly projected; Sc is continued far distad of the costal projection ( Figs 52 View FIGURES 51–58 , 153 View FIGURES 145–153 , 157–158 View FIGURES 154–165 , 217 View FIGURES 209–218 , 278 View FIGURES 278–289 , 365 View FIGURES 348–368 , 412 View FIGURES 406–412 , 454 View FIGURES 452–457 , 540 View FIGURES 539–544 , 671 View FIGURES 664–672 ). The vein MA is non-bifurcate as in all other Furcatergaliae (see Kluge 2004); other veins are either fully developed ( Figs 157–158 View FIGURES 154–165 ), or MP is non-bifurcate (in Ghatula gen. n., Fig. 52 View FIGURES 51–58 ).
(18) Claws of winged stages. On each leg of imago and subimago, both claws are pointed and hooked ( Figs 289 View FIGURES 278–289 , 676 View FIGURES 673–682 ) (see below, discussion about systematic position of the Kimminsula -complex).
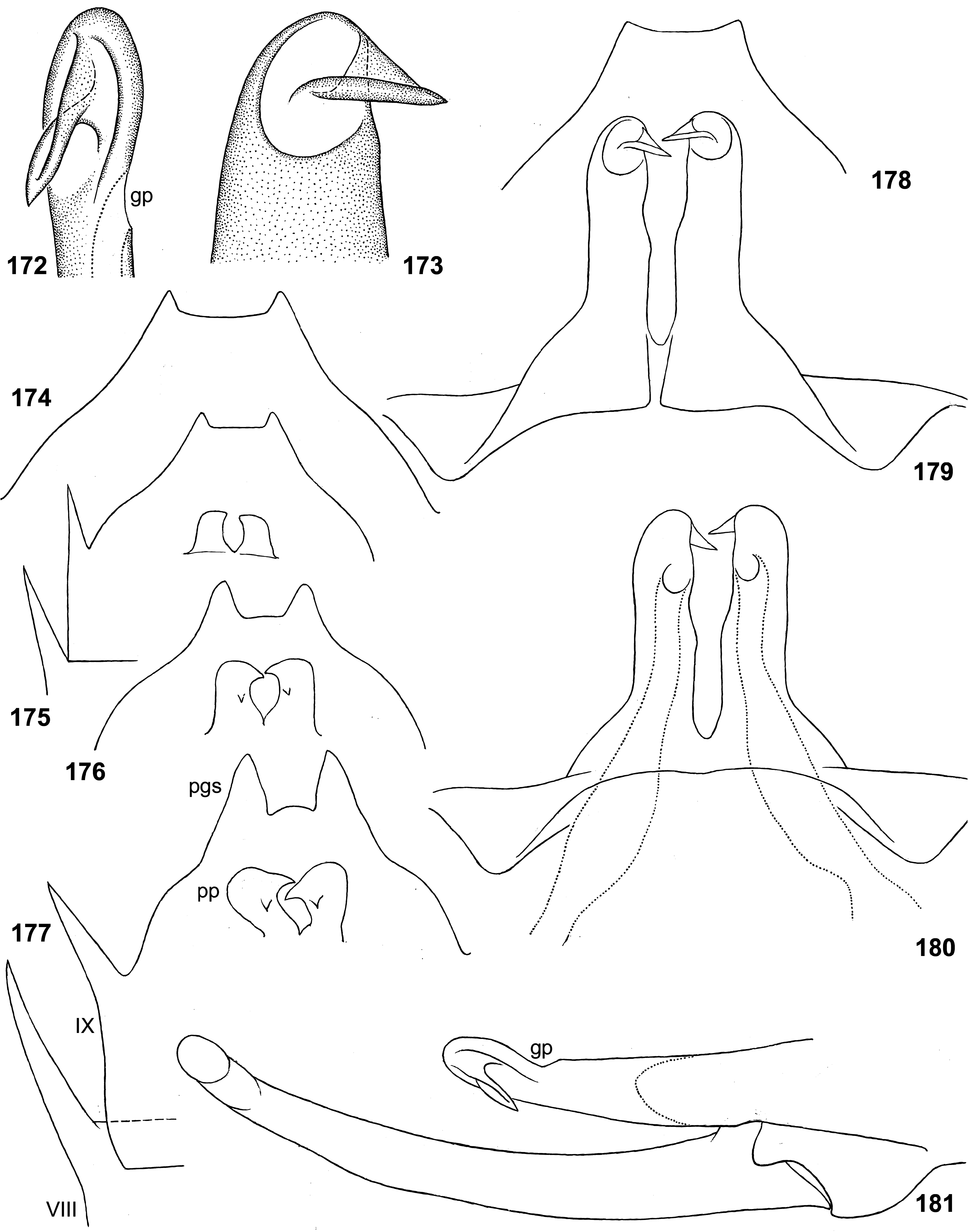
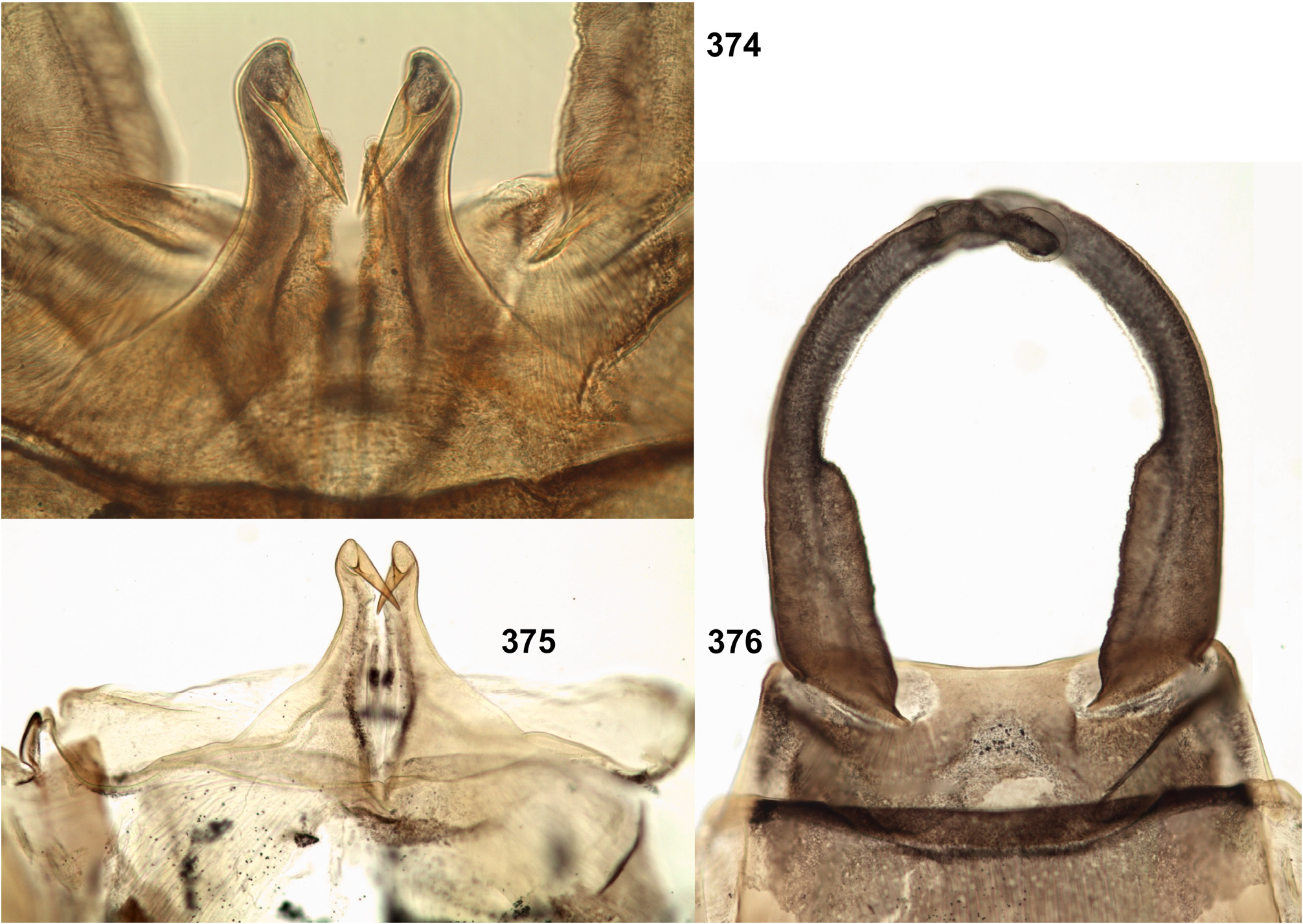
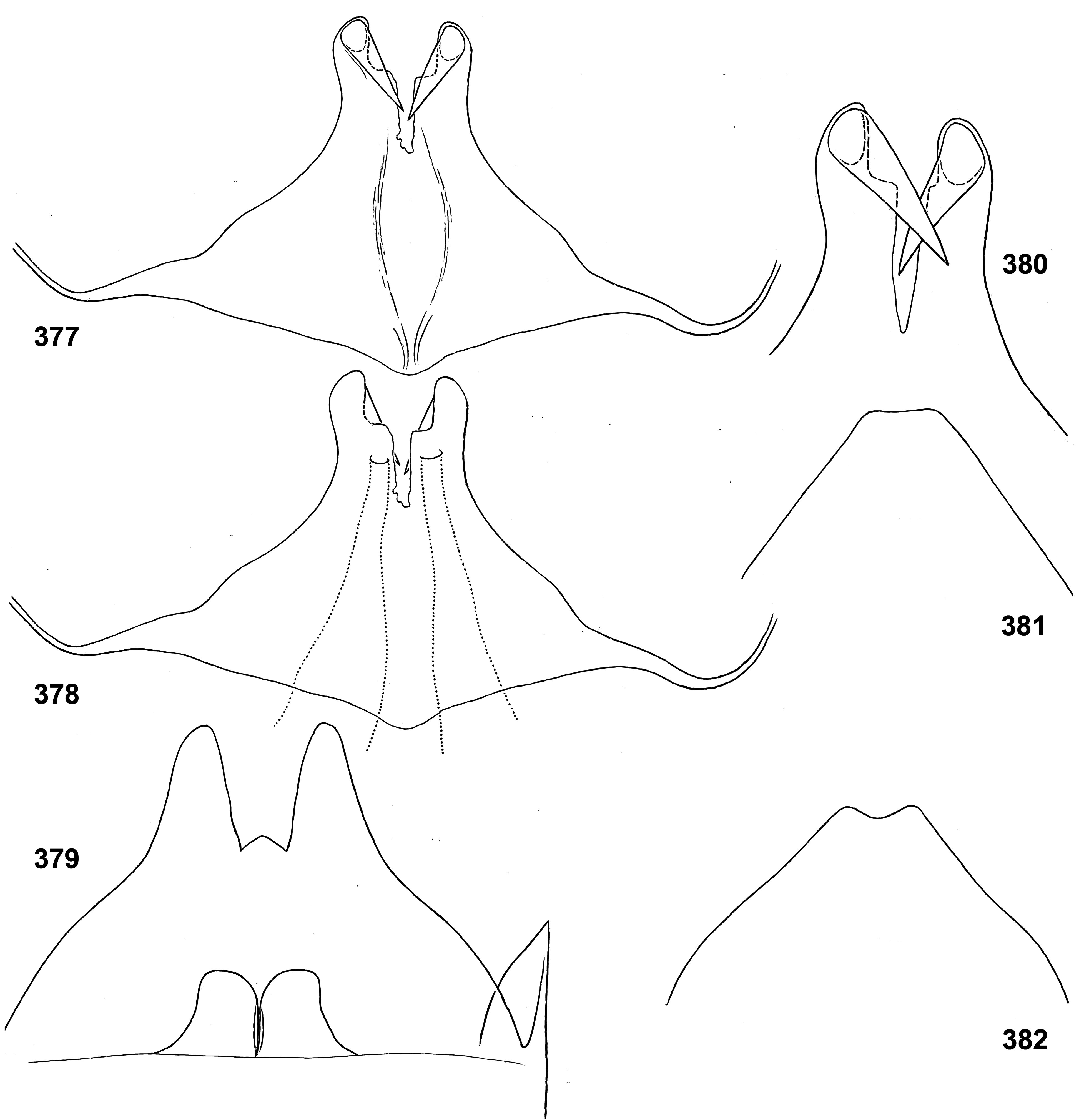
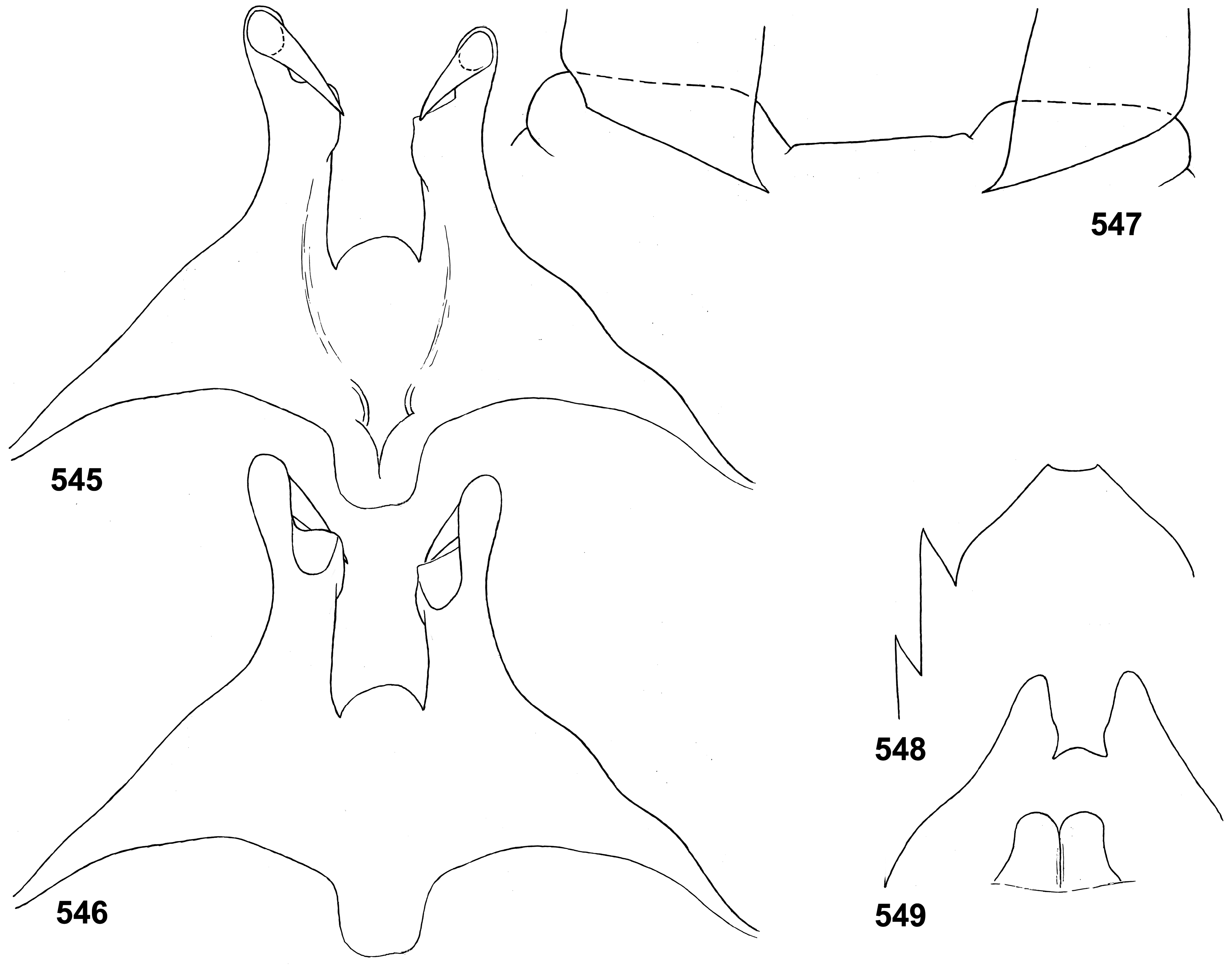
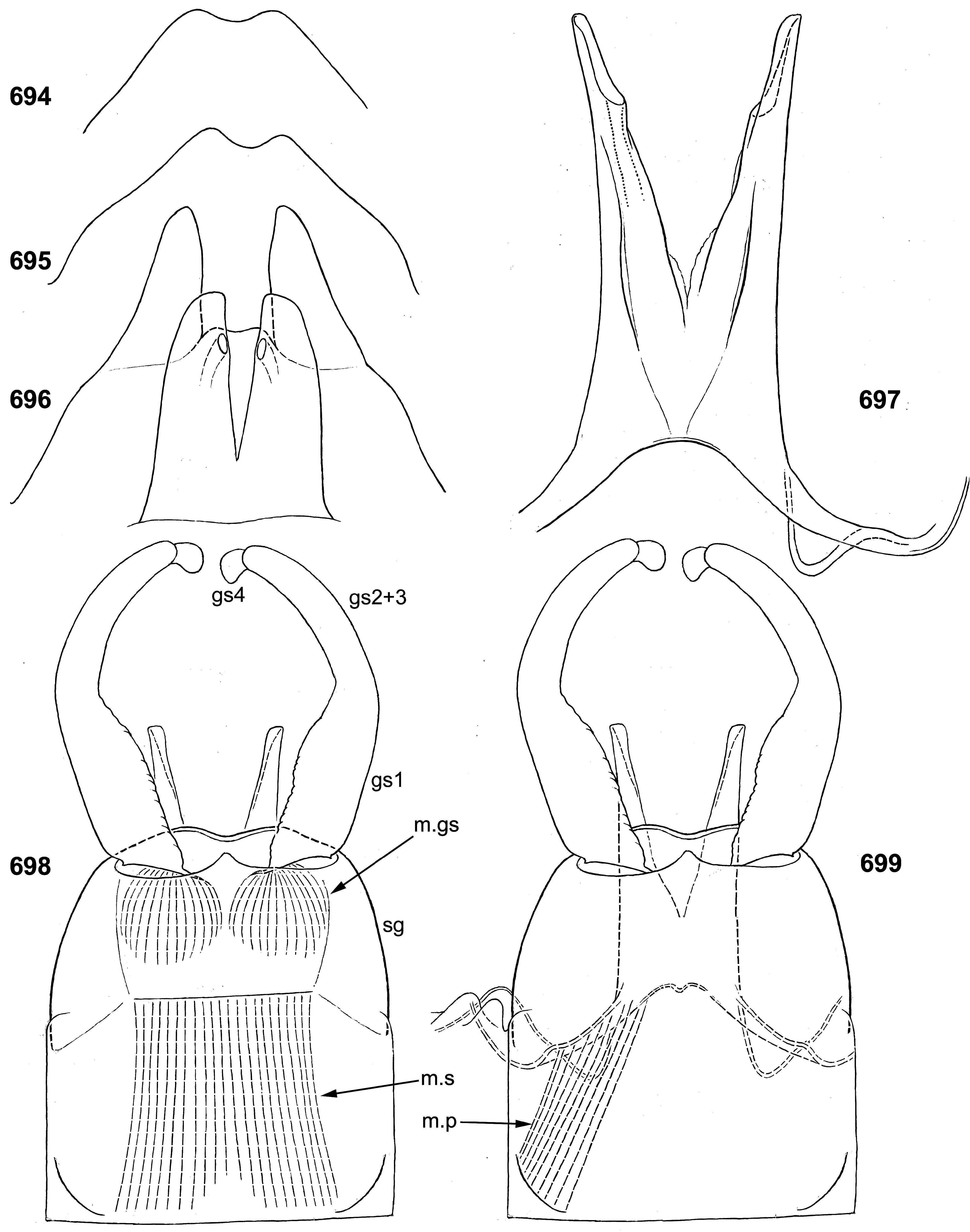
(19) Penis. The pair of penis lobes are movably connected at base, so that are able to move apart and be brought together under action of the sterno-penial muscles. In Petersula the penial lobes are separated nearly to the base ( Figs 179 View FIGURES 172–181 , 183 View FIGURES 182–187 ), while in Ceylonula gen. n., Kimminsula and Hubbardula gen. n., the median sides of penis lobes are connected one with another at significant distance ( Figs 297 View FIGURES 294–299 , 301 View FIGURES 300–305 , 375 View FIGURES 374–376 , 377 View FIGURES 377–382 , 418 View FIGURES 417–420 , 421 View FIGURES 421–427 , 469–470 View FIGURES 468–473 , 534 View FIGURES 528–538 , 545 View FIGURES 545–549 , 690 View FIGURES 688–693 , 697 View FIGURES 694–699 ); in the last case their mobility is ensured by the fact that the cuticle of these median areas is delicate, soft and elastic.
In Petersula , Kimminsula and Ceylonula gen. n., apex of each paired penis lobe bears a pointed process directed medio-ventrally; such processes are not present in in Hubbardula gen. n. ( Figs 690 View FIGURES 688–693 , 697 View FIGURES 694–699 ); genitalia of Ghatula gen. n. are unknown. In taxa with the medio-ventral processes, these processes arise from the penis apices and look similarly, but their structure is significantly different: in Kimminsula they are conic, i.e. round in cross section ( Figs 374–375 View FIGURES 374–376 , 377, 380 View FIGURES 377–382 , 417–418 View FIGURES 417–420 , 421 View FIGURES 421–427 , 469–470 View FIGURES 468–473 , 545 View FIGURES 545–549 ), in Ceylonula gen. n. they are grooved with concavities facing medio-dorsally ( Figs 300–301 View FIGURES 300–305 ), and in Petersula they are grooved with concavities faced latero-ventrally ( Figs 172–173, 179 View FIGURES 172–181 ).
In Petersula , the pointed projections are already present on larval protopenis, where they have the same position as in imago ( Figs 175–177 View FIGURES 172–181 , 209 View FIGURES 209–218 ). In Ceylonula gen. n. and Kimminsula , larval protopenis has no any precursors of these projections ( Figs 304–305 View FIGURES 300–305 , 379 View FIGURES 377–382 , 425–426 View FIGURES 421–427 , 549 View FIGURES 545–549 , 602 View FIGURES 601–606 ).
In all cases (in Petersula , Kimminsula , and Ceylonula gen. n.) the medio-ventral pointed processes arise from the ventral sides of penis lobes, significantly distad to the gonopores, while the gonopores are opened on the dorsal sides of the penis lobes. The medio-ventral pointed processes have no connection with the gonoducts: in Petersula and Ceylonula gen. n. their grooves do not touch the gonopores ( Figs 172 View FIGURES 172–181 , 300 View FIGURES 300–305 ), and in Kimminsula they have no any grooves or canals at all. The pair of gonoducts passing inside the penis lobes, have no musculated sperm pumps, in contrast to Thraulodes ( Kluge 2020: figs 101, 111) and some others.
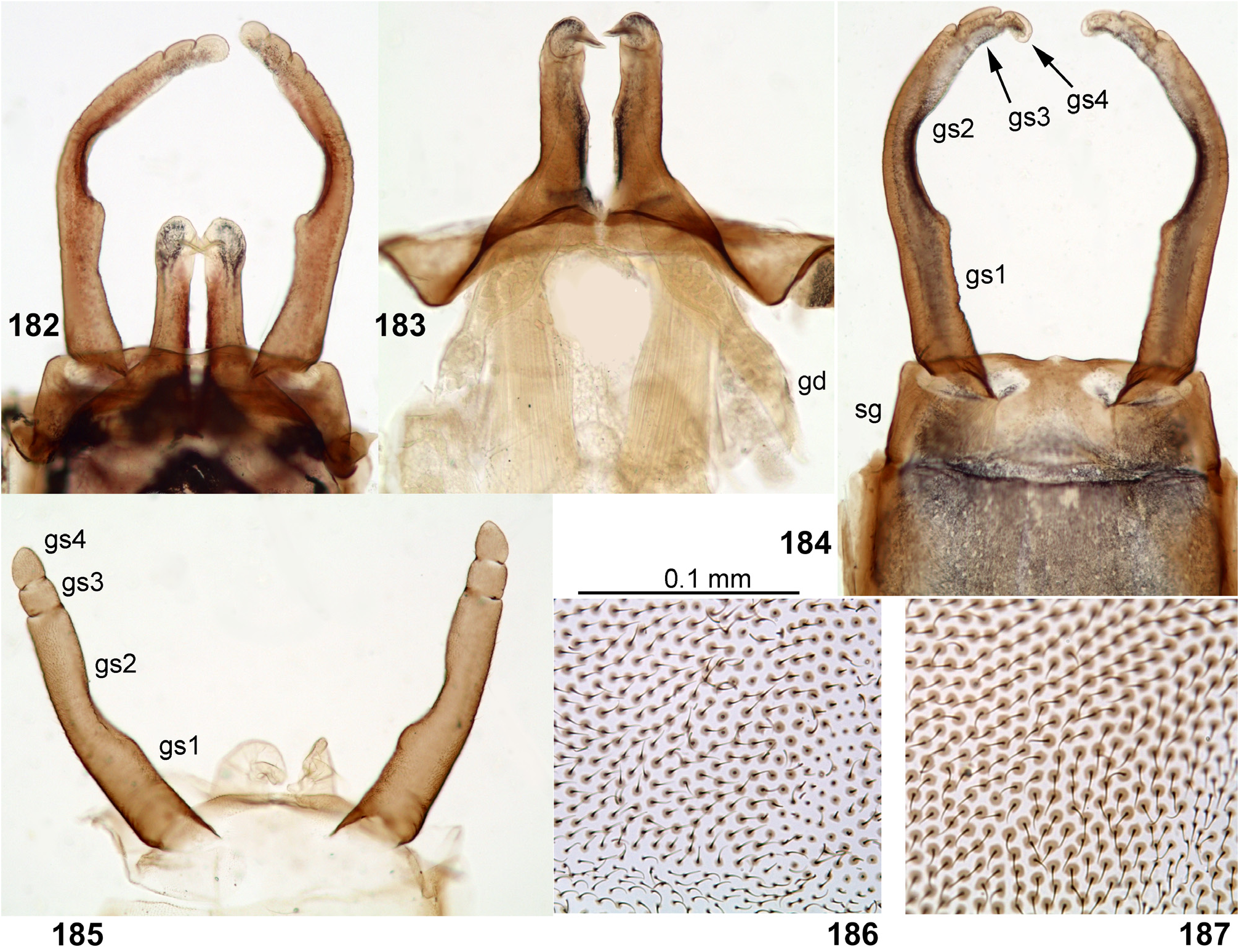
(20) Styliger and gonostyli. The posterior-dorsal margin of styliger is distinct, and its postero-ventral margin is not expressed, so the gonostyli are attached on ellipsoid membranous cavities faced ventro-caudally and well visible from ventral view ( Figs 181 View FIGURES 172–181 , 184 View FIGURES 182–187 ). In other respects styligers and gonostyli are significantly diverse: in Petersula and Kimminsula the styliger is short and simple, and the gonostyli retain division into 4 segments ( Figs 184 View FIGURES 182–187 , 376 View FIGURES 374–376 , 420 View FIGURES 417–420 , 472 View FIGURES 468–473 ); in Ceylonula femoralis the styliger is projected medially, and the gonostyli lack boundary between the initial 1st and 2nd segments ( Fig. 296 View FIGURES 294–299 ); in Hubbardula heterolepida sp. n. the styliger is elongated and modified, the membranous cavities of gonostyli attachments are connected medially, and the gonostyli lack boundary between the initial 2nd and 3rd segments ( Figs 689, 691 View FIGURES 688–693 , 698 View FIGURES 694–699 ).
Composition of the Kimminsula -complex
This complex includes two south Indian genera, Petersula Sivaramakrishnan 1984 and Ghatula gen. n., and three Ceylonese genera, Ceylonula gen. n., Kimminsula Peters & Edmunds 1970 and Hubbardula gen. n. Key characters of these genera are given in Table 1 View TABLE 1 .










































































No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |