
Chileanthicus Werner, 1966
|
publication ID |
https://doi.org/ 10.11646/zootaxa.2180.1.1 |
|
persistent identifier |
https://treatment.plazi.org/id/3949251E-FFA5-776C-FF63-D9D0FC50FA96 |
|
treatment provided by |
Felipe |
|
scientific name |
Chileanthicus Werner, 1966 |
| status |
|
( Figs 1–8 View FIGURES 1–8 )
Chileanthicus Werner, 1966: 222 . Type species: Formicomus lafertei Solier, 1851 , fixed by original designation.
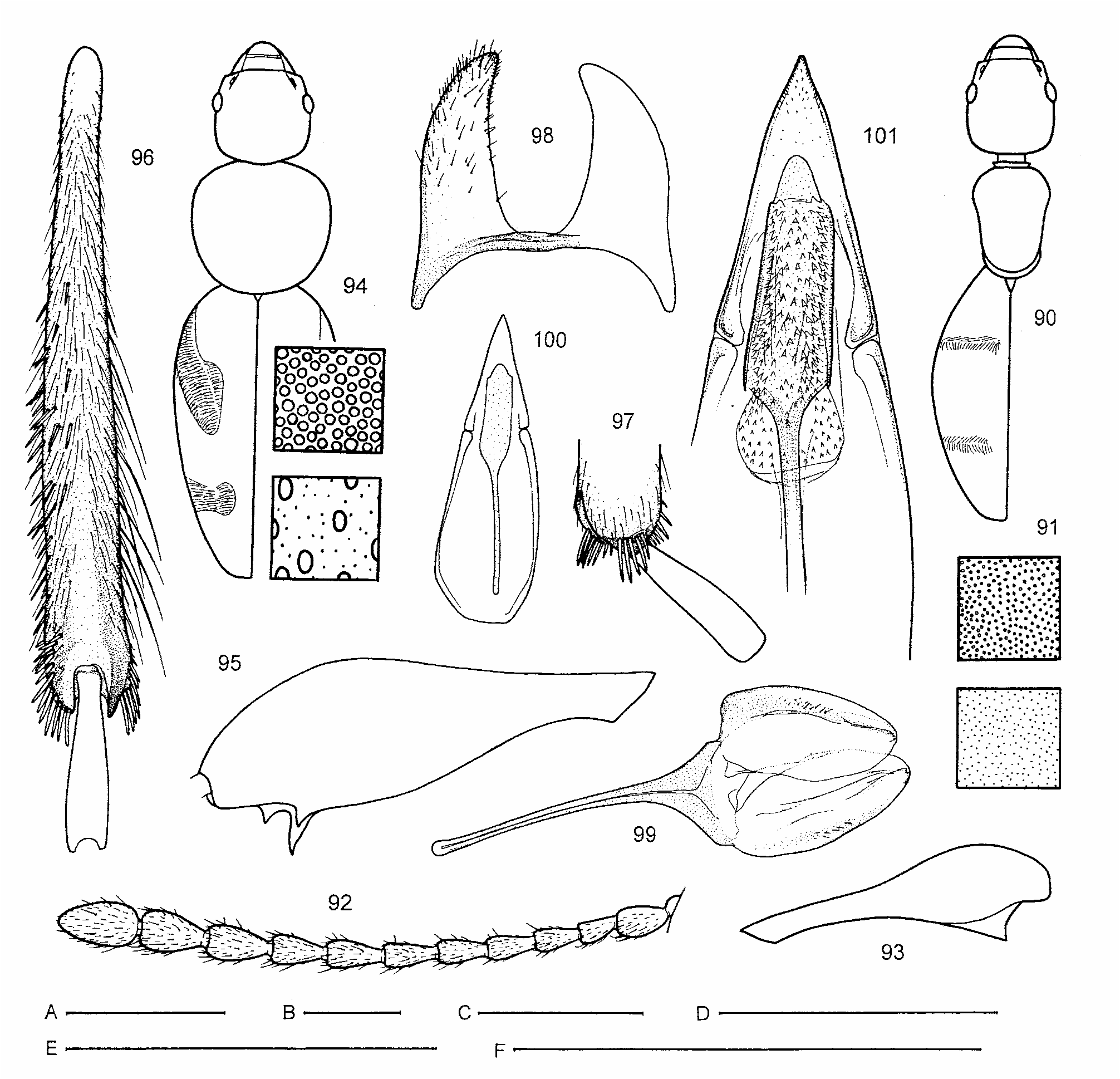
Redescription. Anthicids of small to medium size (2.4–6.2 mm). Head with frontoclypeal sulcus fine but distinct; antennal insertion exposed and clearly visible; neck short and thick, always well-differentiated from head; eyes small to medium in size; the last maxillary palpomere rather small, triangular ( Fig. 5 View FIGURES 1–8 ).
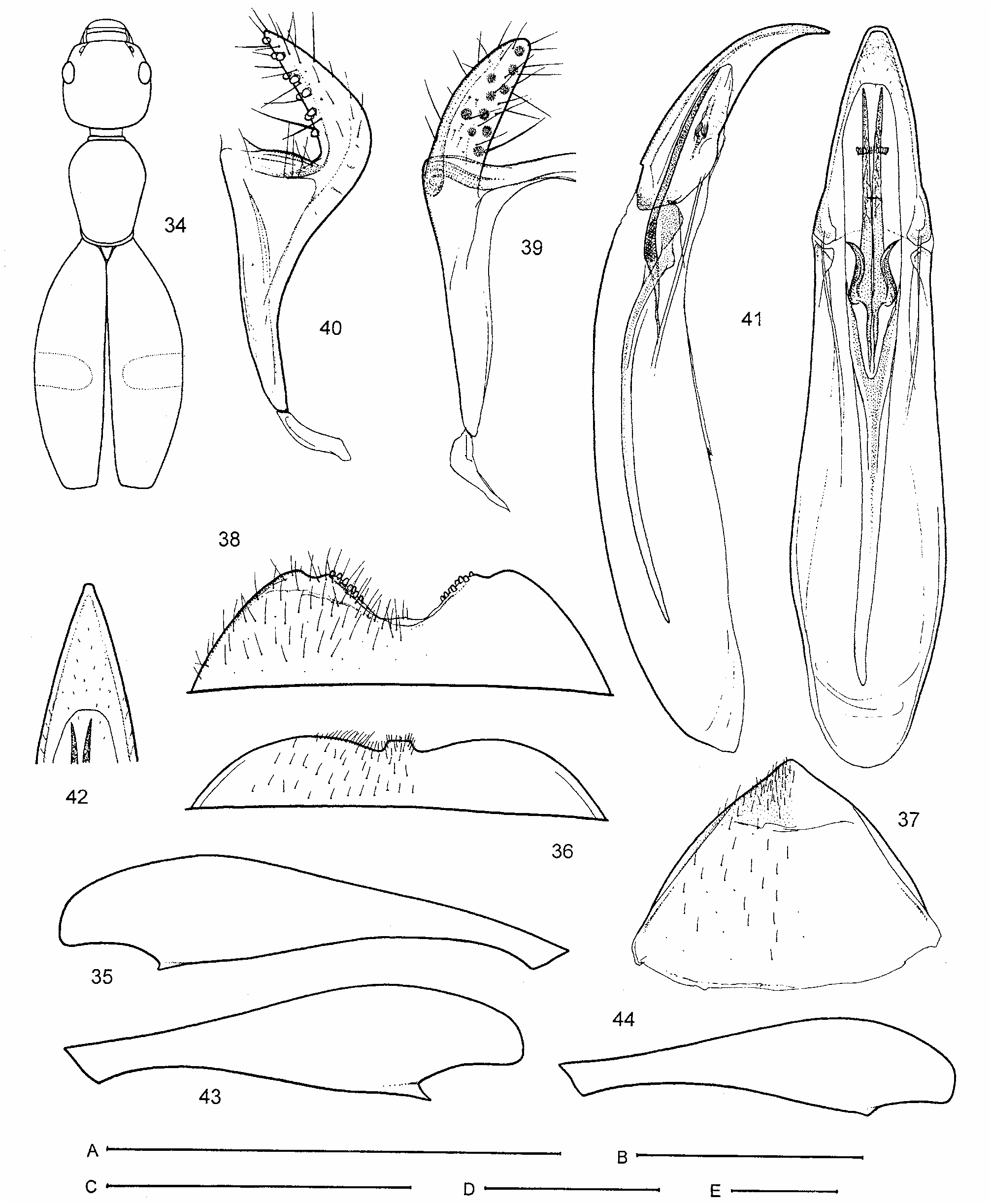
Pronotum with distinct apical flanged rim (collar) and basal margin distinctly bordered by thin, welldefined sulcus originating at lateral foveae ( Fig. 2 View FIGURES 1–8 ), the latter may be hidden in dorsal view under posteriorly strongly convex/bulging base of pronotum; dorso-lateral margins of pronotum often angulate, forming rounded to sharp edge close behind widest point, and pronotal disc thus well-delimited and more conspicuous (most Australian species). Mesosternum triangular, with lateral margins nearly straight, its anterior angle separated from anterior margin of mesothorax by very narrowly joined mesepimera ( Fig. 1 View FIGURES 1–8 ); intercoxal process of mesosternum fully separating mesocoxae (its apex mostly touching metasternum, unlike in Fig. 1 View FIGURES 1–8 ). Lateral margins of mesepisterna simply connected (not raised) with mesepimera, bare, without distinct fringe of setae. Mesoscutellum triangular, tapering posteriorly ( Fig. 3 View FIGURES 1–8 ). Prosternum area beneath coxae wellsclerotized and extended posteriorly as distinct sclerite; posterior margin of this sclerite shallowly emarginate and angulately produced medially ( Fig. 1 View FIGURES 1–8 ). Metendosternite with short stalk, arms narrow and obsolete ventrolateral projections/lamina ( Fig. 4 View FIGURES 1–8 ). Elytra more or less convex, mostly rather strongly declivous at scutellar area towards base, postscutellar impression and elytral humeri indistinct; apical notches in males absent; sutural striae indistinct. Posterior wings strongly reduced to absent (all know species are flightless).
Metafemora armed with 1–3 pointed teeth on inner side subapically; penultimate tarsomere simple (not clearly bilobed), with terminal tarsomere articulated apically, especially in meso- and metatarsi; paired terminal spurs mostly well-developed and distinct, rarely reduced for metatarsi (a few Australian species).
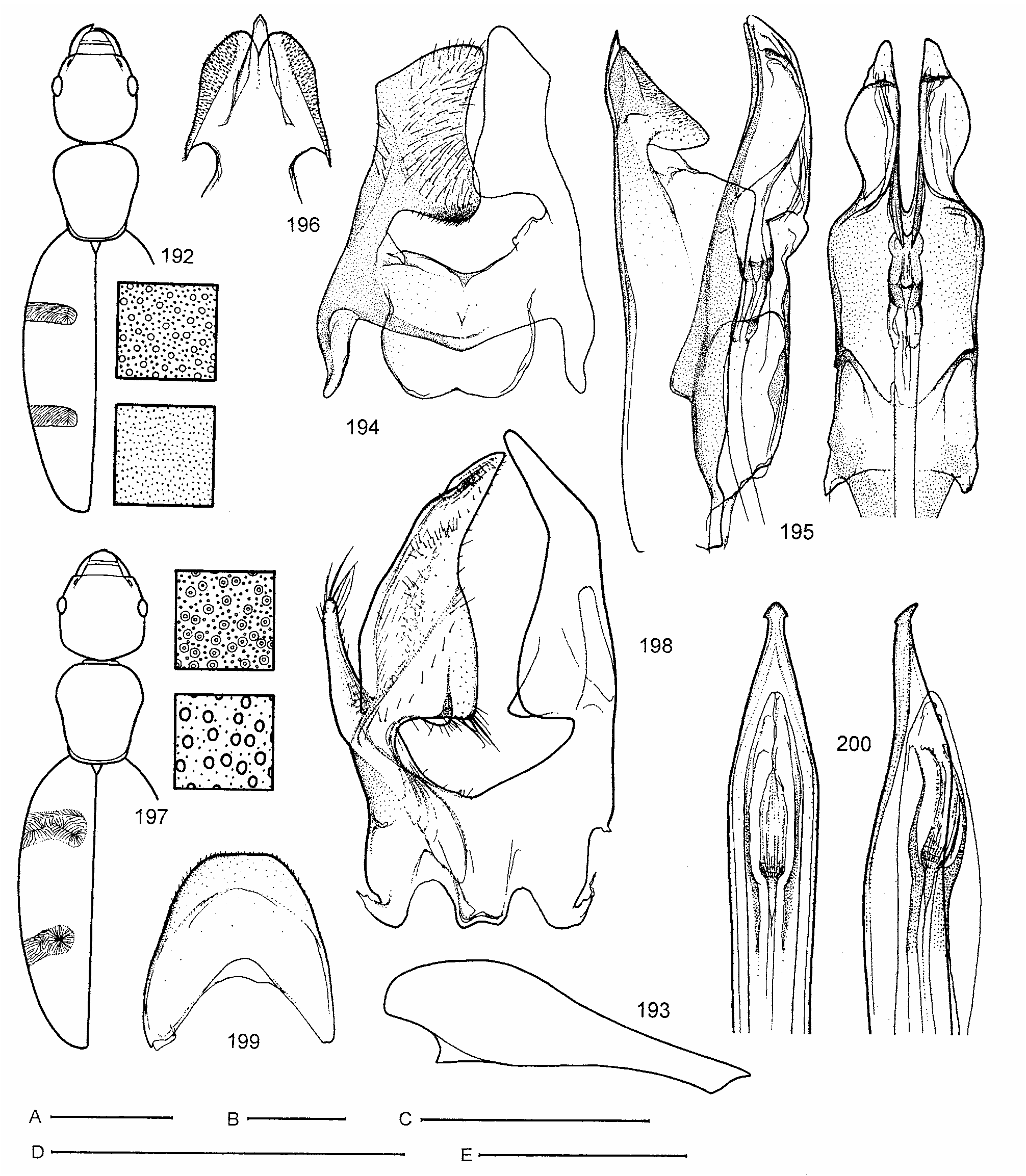
Abdominal sternum III without foveae immediately behind metacoxae, its intercoxal process truncate apically, incompletely bordered (only laterally) in ventral view ( Fig. 1 View FIGURES 1–8 ). Male sternite VIII modified, with pair of posteriad projecting prongs; prongs more or less tightly joined medially, latero-basal plates very small and inconspicuous. Male tergite VIII forming single sclerite ( Figs 25 View FIGURES 17–26 , 30 View FIGURES 27–33 , 138 View FIGURES 135–145 , 199 View FIGURES 192–200 , 217, 219 View FIGURES 209–219 ). Male segment IX (spiculum) forked distally, mostly T-shaped, with a pair of apical sclerites ( Figs 51 View FIGURES 45–53 , 84 View FIGURES 81–89 , 99 View FIGURES 90–101 , 126 View FIGURES 123–134 , 139, 144 View FIGURES 135–145 , 158 View FIGURES 155–159 , 163 View FIGURES 160–170 , 181 View FIGURES 171–182 , 203 View FIGURES 201–208 , 218 View FIGURES 209–219 ).
Aedeagus with tegmen clearly divided into apical portion (formed by fused parameres) and basal-piece; median lobe strengthened by sclerotized apodeme, gonopore situated in apical portion of aedeagus. Female ovipositor without apparent styli and conspicuously setose ( Fig. 8 View FIGURES 1–8 ).
Sexual dimorphism. Males of Chileanthicus are usually somewhat smaller and more slender than females, sternum VII is sometimes modified, legs are more robust with more or less produced inner apical margin of meso- and/or metatibiae, and there is different setation of the metatibiae, with longer, stiffer, or sometimes tufted setae (numerous Australian species).
Females rarely exhibit modifications that are not present in males: tapering elytral apices ( C. acutipennis sp. nov., Fig. 240 View FIGURES 232–241 ), sternum VII with single or paired median process ( C. lafertei , C. femineus sp. nov., Figs 21 View FIGURES 17–26 , 36 View FIGURES 34–44 ), or bulging tergum VII ( C. penai , Fig. 53 View FIGURES 45–53 ). In most females, tergum VII appears less widely rounded posteriorly than in the males, rather triangular, angularly produced at apex.
Immature stages. Unknown.
Biology. The life history of members of Chileanthicus is essentially unknown. Based on the label data, specimens have been collected by sifting leaf litter in forests, by beating shrubs, looking under stones (most Chilean specimens; F. Ramírez, J. E. Barriga pers. comm.) or on fresh dung, and by using pitfall traps.
Lea (1922) recorded two Australian species as being “found in ant nests”: C. quadrimaculatus ( King, 1869) with Pachycondyla lutea (Mayr, 1862) , and C. speciosus ( King, 1869) with Iridomyrmex conifer Forel, 1902 . I have examined the respective specimens, and the two females of the former species proved to be misidentified and belong probably to C. pumilio sp. nov. (see Remarks under this species). Another two specimens regarded as inquilines were found in the McMillan collection (WAM): C. audax sp. nov. was found with a ponerine species (probably Hypoponera Santchi, 1938 ), and C. speciosus with a species of Camponotus Mayr, 1861 . Many anthicids are known to occur with ants, mimicing their appearance and movements for (probably) protective reasons ( Hingston 1925, Wasmann 1898, Werner & Chandler 1995). True myrmecophily of the above-mentioned species is therefore uncertain, and must be corroborated by more exact observations.
Distribution. Gondwanian; so far known from Chile (Atacama and Coquimbo), Australia, and Madagascar ( Fig. 242 View FIGURE 242 ). This genus appears to be more speciose in semi-arid and arid regions. In Chile it is presently restricted to the arid, northern part of the Chilean Coast Range, between Copiapo in the north and Ovalle in the south. With this information, the occurrence of C. cursor sp. nov. in the montane rainforest area of eastern Madagascar is surprising. This species is based on two specimens and a labeling mistake cannot be excluded.
Relationships. Chileanthicus is currently placed in the Formicomini Bonadona, 1974 , together with Andrahomanus Pic, 1903 , Stenidius LaFerté-Sénectère, 1847 and Anthelephila Hope, 1833 (see Kejval, 2003). Within the tribe Chileanthicus can be recognized by the following characters: 1) basal sulcus of pronotum distinct both dorsally and laterally, reaching as far as to lateral foveae; 2) anterior angle of mesosternum separated from anterior margin of mesothorax by very narrowly joined mesepisterna; 3) posterior membranous wings reduced (all known species are flightless); 4) metafemora armed with 1–3 pointed tooth-like protrusions on inner side subapically (equally developed in both sexes); 5) male abdominal sternite VIII modified, but less differentiated, paired prongs of sternite VIII more or less tightly joined medially, latero-basal plates inconspicuous; 6) male tergite VIII forming single sclerite; 7) male segment IX with a pair of distinct apical sclerites; 8) gonopore situated rather in apical portion of aedeagus; 9) ovipositor rather short and wide, without apparent styli and is conspicuously setose.
At least four characters of Chileanthicus (2, 5–7) appear to be primitive and suggest, along with the Gondwanian distribution, the rather basal position of this genus within the Formicomini . The modified metafemora (4) and the peculiar morphology of the ovipositor (8) can be regarded as derived character states. The first character is an autapomorphy for the genus. The second may represent a possible synapomorphy supporting a close relationship to Andrahomanus Pic, 1903 , which is known from Madagascar and Southern Africa (Kejval, in prep.), and differs only by the simple metafemora for both sexes.
As for other genera of Formicomini , Stenidius shares several characters (2, 5–7), and a similar body form (in the apterous species), but differs by the reduced basal sulcus of the pronotum (varying from slightly indicated dorso-medially to absent), simple metafemora, and ovipositor with well-developed styli (see Kejval, 2002). Anthelephila , the most derived member of the tribe, differs in many respects, mainly by the medially widely joined mesepimera, simple metafemora, and ovipositor with well-developed styli (see Kejval, 2003). It also displays a reduced basal sulcus of the pronotum (indistinct laterally), and a more complicated structure of abdominal segment VIII (the sternite is differentiated into five discrete parts, the tergite is composed of two sclerites); however the latter character is unstable and the condition in some species may resemble that in Chileanthicus (see Kejval, 2007).
Remarks. Werner (1966) mentioned the similarity of Chileanthicus to the Australian Formicomus quadrimaculatus King, 1869 in sharing aptery, dentate metafemora in both sexes, and similar form and setation of the female ovipositor. However, he was not really familiar with Formicomus nor the Australian Anthicidae , and he placed great importance on differences in body setation, position of the metafemoral process, and to form and inner structure of the aedeagus. Based on my studies ( Kejval, 2003, 2005), F. quadrimaculatus and its relatives differ clearly from Anthelephila (= Formicomus ), both morphologically and in distributional pattern. On the other hand, they share all the above-mentioned characters of Chileanthicus . The differences pointed out by Werner (1966) are, in my opinion, less important, variable characters, which is evident once more extensive material is studied. The Australian species, unlike the Chilean ones, are extremely diversified both in external and male characters, which is probably related to their greater diversity of habitats and extent of their distributional range. Even the two species known from Madagascar, undoubtedly very closely related, differ mutually in body setation and structure of the median lobe of the aedeagus. Remarkably, one of them ( C. dentatus ; Fig. 61 View FIGURES 54–62 ) displays the same peculiar peg-like setation of the prongs of male sternite VIII, that is typical for all the Chilean species.















No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
Chileanthicus Werner, 1966
| Kejval, Zbyněk 2009 |
Chileanthicus
| Werner, F. G. 1966: 222 |