
Echinolittorina omanensis, Published, 2007
|
publication ID |
https://doi.org/10.11646/zootaxa.1420.1.1 |
|
publication LSID |
lsid:zoobank.org:pub:B2E8D420-9177-44DB-9807-12472877F48F |
|
persistent identifier |
https://treatment.plazi.org/id/3B108794-5915-FFE1-BAF1-5F1DEEB64AF3 |
|
treatment provided by |
Felipe |
|
scientific name |
Echinolittorina omanensis |
| status |
sp. nov. |
Echinolittorina omanensis View in CoL new species
( Figures 6G, H View FIGURE 6 , 16 View FIGURE 16 , 17 View FIGURE 17 , 18 View FIGURE 18 )
Littorina (Tectus) trochoides — Nevill, 1885: 156–157 (in part, includes E. marquesensis , E. malaccana ; not Gray, 1839 = E. pascua ).
Nodilittorina (Nodilittorina) trochoides — Reid, 1992: 202 (in part, includes E. malaccana , E. austrotrochoides , E. cecillei; not Gray, 1839).
Tectarius nodulosus — Melvill & Standen, 1901: 364 (in part, includes E. malaccana ; not Gmelin, 1791 = E. malaccana group or E. pascua ).
Nodilittorina nodulosa — Fischer, 1969: 119–129 (in part, includes N. pyramidalis , E. natalensis , E. malaccana , E. austrotrochoides , E. cecillei , E. marquesensis , E. wallaceana ; not Gmelin, 1791). Fischer, 1971: 31–32 (in part, includes E. natalensis , E. malaccana ; not Gmelin, 1791).
Nodilittorina (Nodilittorina) natalensis — Rosewater, 1970: 489–490, pl. 377 (map) (in part, includes E. natalensis ; not Krauss in Philippi, 1847). Mienis, 1973: 59–61, fig. 3 (not Krauss in Philippi, 1847). Bosch et al., 1995: 46, fig. 117 (not Krauss in Philippi, 1847).
Nodilittorina natalensis — Reid, 2002a: 259–281 (in part, includes E. natalensis ; not Krauss in Philippi, 1847).
Echinolittorina natalensis — Williams et al., 2003: 83 (in part, includes E. natalensis ; not Krauss in Philippi, 1847).
Echinolittorina subnodosa B—Williams & Reid, 2004: 2227–2251.
Types: Holotype BMNH 20040218 ( Fig. 17J View FIGURE 17 ); 1 dry paratype BMNH 20040219; 6 alcohol paratypes BMNH 20040220; Wadi Sayq , Jabal Qamr, Dhofar, Oman, 16°44’N, 53°20’E. GoogleMaps
Etymology: Latin, from Oman.
Taxonomic history: Nodulose shells from southern Arabia and Somalia have sometimes been identified as E. natalensis in the past (Rosewater 1970; Mienis 1973; Bosch et al. 1995), probably on the basis of geographical proximity. Nevertheless, the narrow shells, often with two rows of nodules, more closely resemble those of the E. malaccana group, with which they have also been classified (Nevill 1885; Melvill & Standen 1901; Fischer 1969, 1971; Reid 1992). They were recognized as distinct following molecular study (Williams & Reid 2004).
Diagnosis: Shell high-conical, 2–3 rows of rounded nodules on last whorl, peripheral nodules axially elongated and crossed by 3–4 spiral ribs; 10–16 spiral ribs on last whorl (including base); grey or black with white nodules. Somalia, southern Arabia, Pakistan. COI: GenBank AJ623059 View Materials , AJ623060 View Materials .
Material examined: 38 lots (including 15 penes, 1 sperm sample, 5 pallial oviducts, 2 radulae).
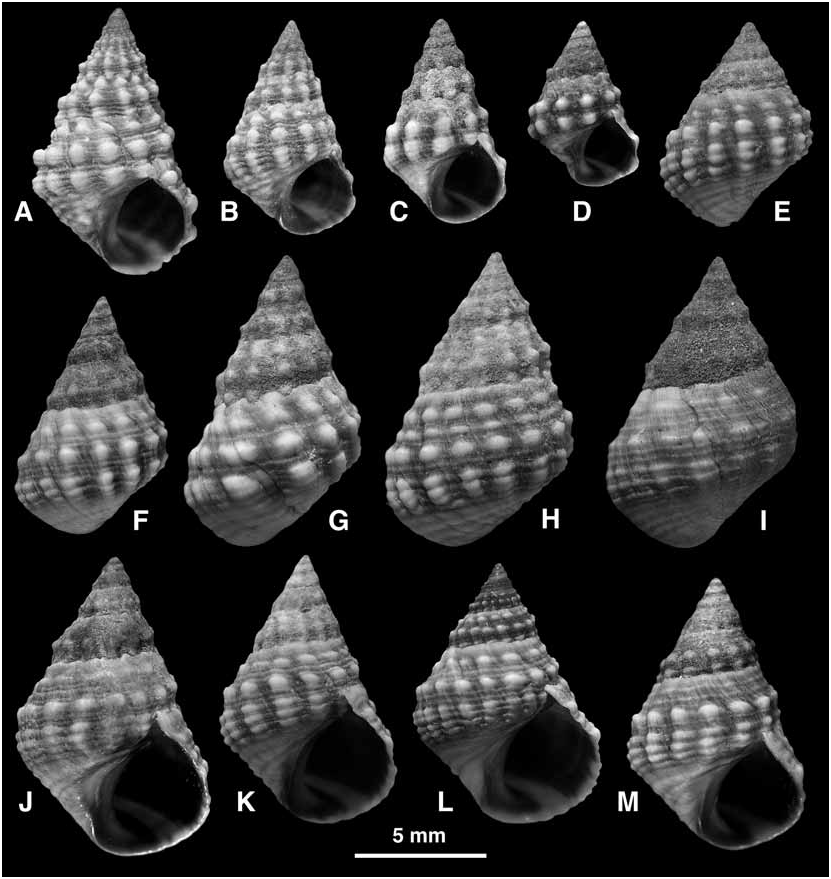
Shell ( Fig. 17 View FIGURE 17 ): Mature shell height 4.3–17.2 mm. Shape conical to high-conical (H/B = 1.36–1.75; SH = 1.61–2.27); spire whorls lightly rounded, suture distinct; spire profile almost straight; periphery of last whorl weakly angled. Columella short, concave, hollowed at base; small eroded parietal area. Sculpture of last whorl: 2 rows of rounded nodules at periphery and mid-whorl, nodules rarely pointed ( Fig. 17A View FIGURE 17 ), often an additional row of smaller nodules at shoulder ( Fig. 17A, B, E–H, K, L View FIGURE 17 ); nodules aligned to form 9–17 axial series; entire surface with spiral ribs, 7–10 at and above periphery, and microstriae; peripheral nodules axially elongated and crossed by 3–4 ribs; base with 4–6 ribs, occasionally with weak nodules; rarely the nodules on last whorl become weak ( Fig. 17I View FIGURE 17 ). Protoconch approximately 0.25 mm diameter. Colour: black to purplebrown (fading to blue-grey), nodules white; aperture brown to black with pale band at base; columella purplebrown.
Animal ( Fig. 18 View FIGURE 18 ): Head black, no unpigmented stripe across snout, tentacle pale around eye and sometimes also inner side of base, with two longitudinal grey to black lines, usually partly fused and extending half to full length of tentacle; sides of foot grey to black. Opercular ratio 0.43–0.54. Penis ( Fig. 18A–H View FIGURE 18 ): filament gradually tapering to pointed tip, with fine annular wrinkles for most of its length, filament 0.5–0.6 total length of penis, sperm groove extends to tip; mamilliform gland about equal to glandular disc, borne together on projection of base; penis unpigmented or slightly pigmented at base. Euspermatozoa not known; paraspermatozoa spherical to oval, 10–14 µm diameter, filled with large round granules, containing single short rectangular rod-piece, hexagonal in section and not projecting from cell. Pallial oviduct ( Fig. 18I View FIGURE 18 ): bursa opening at half length of straight section and extending back to albumen gland. Development predicted to be planktotrophic.
Radula ( Fig. 6G, H View FIGURE 6 ): Relative radula length 2.45–9.51. Rachidian: length/width 1.41–1.69; tip of major cusp pointed. Lateral and inner marginal: tips of major cusps rounded; major cusp of lateral slightly larger than that of inner marginal. Outer marginal: 7–8 cusps.
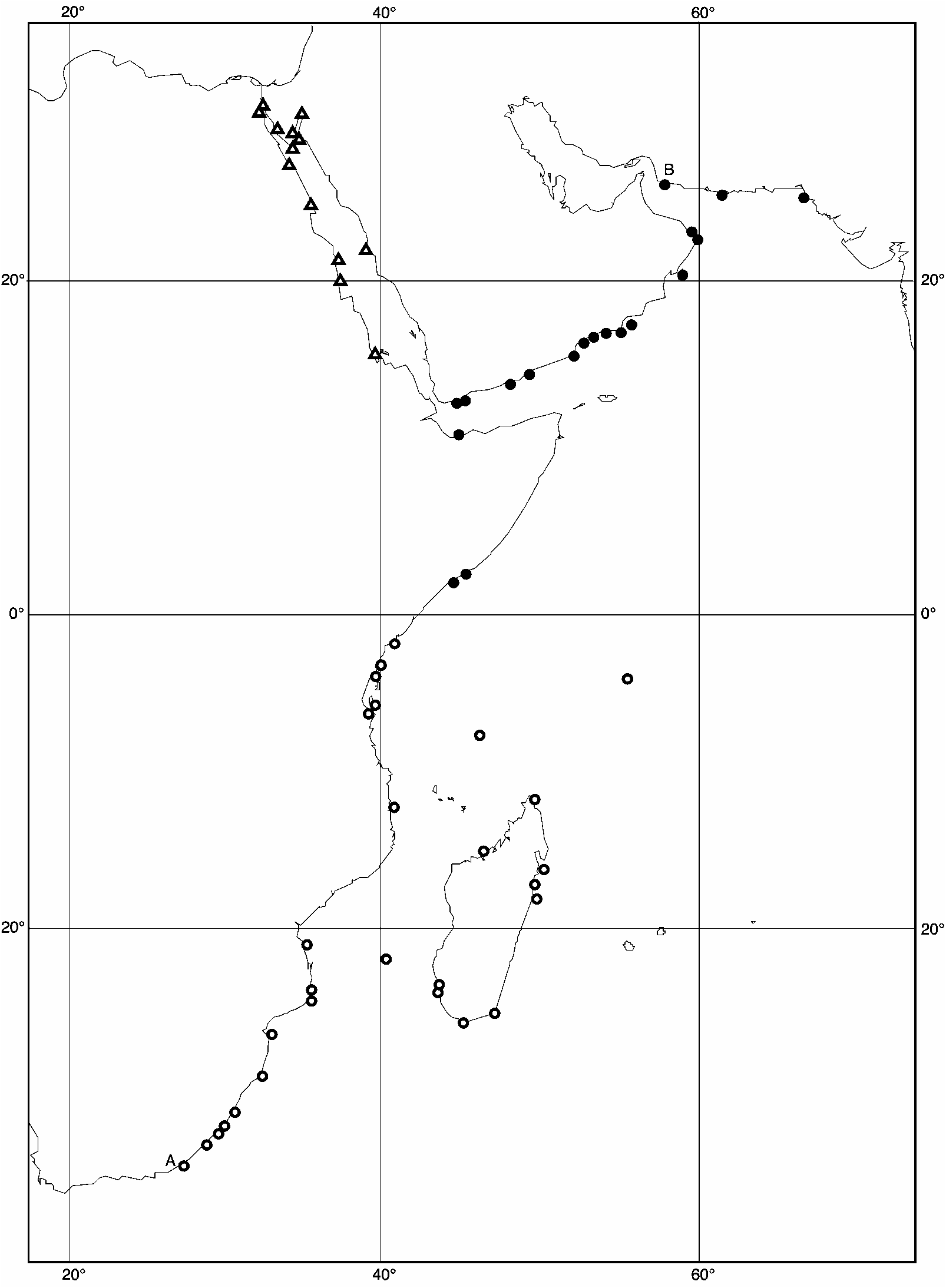
Range ( Fig. 16 View FIGURE 16 ): Somalia, Gulf of Aden, northwestern Arabian Sea to Pakistan. Range limits: Merka, Somalia (HUJ 30002); Mogadishu, Somalia (HUJ 30005, 30967); Berbera, Somalia (BMNH); Little Aden, Yemen (BMNH); Sur, Oman (BMNH); Ras al-Junayz, Ras al-Hadd, Oman (BMNH); Jask, Iran (Melvill & Standen 1901); Ramin, Iran (ZMA); Karachi, Pakistan (BMNH 20040215). The absence of records from most of the coast of Somalia is explained by the inaccessibility of the region. The absence from the southern Gulf of Oman is probably real (there are 10 records of E. arabica from Muscat and further north on the coat of Oman, but none of E. omanensis ), as is that from the Red Sea (there are three records of E. marisrubri from the west coast of Yemen).
Habitat: This species occurs in the littoral fringe on rocks and cliffs, on sheltered and more commonly on wave-exposed coasts (G.R. Feulner pers. comm.). It has been recorded on limestone, basalt and concrete sea walls.
Remarks: The geographical distribution of this species corresponds closely with the area of seasonally high primary productivity associated with the upwelling induced by the southwestern monsoon of the boreal summer (Wilson 2000; Wilson & Klaus 2000; Rutgers University Primary Productivity Study). This coastline is also one of generally high wave exposure. Further south than Mogadishu and Merka (the southernmost records of the species) the coast is protected by coral reefs (Carbone & Accordi 2000). Similarly, the coast of the Gulf of Oman between Ras al-Hadd and Muscat supports reefs and mangroves, while further west a sandy beach stretches to Fujairah (Wilson 2000). The species is present on the exposed, nutrient-rich Makran Coast of Iran and as far east as Karachi, but its eastward distribution is then limited by the sedimentary or estuarine conditions of the Gulfs of Kutch and Khambha, where no Echinolittorina species have been reported. Echinolittorina omanensis has not been recorded from the Red Sea, despite known inflow from the Gulf of Aden and consequent high summer nutrient levels in the southernmost Red Sea (Sheppard 2000).
The coastlines of Yemen and Oman have been relatively well collected (material in BMNH, ZMA; G.R. Feulner pers. comm.) and, consequently, it is clear that between Aden and Ras al-Hadd E. omanensis is the only abundant species of Echinolittorina . Also occurring on this coastline is E. millegrana which, while common at Aden and Ras al-Hadd, is scarce at intervening localities; E. millegrana occupies a slightly lower tidal level. Predatory intertidal crabs are common on the Arabian coastline and may restrict the microdistribution of E. omanensis and its congeners (G.R. Feulner pers. comm.); repaired shell breakages that may have been caused by unsuccessful attacks by crabs are frequent in some samples.
Echinolittorina omanensis overlaps barely or not at all with the remaining Echinolittorina species of the western Indian Ocean. In northern Oman, E. omanensis has been recorded together with E. arabica at only two localities: Ras al-Junayz, Ras al-Hadd (BMNH; both species common) and Sur (BMNH; E. omanensis rare, E. arabica sparse). At these localities their respective microdistributions are not known, but the habitat characteristics of these species elsewhere suggest that both occupy similar high levels in the littoral fringe, with E. arabica restricted to microhabitats that are more sheltered from wave action. In the vicinity of Aden, both E. omanensis and E. marisrubri have been recorded from Little Aden and Conquest Bay (BMNH), but the latter species is rare. Both species show three rows of nodules on the shell, but the intervening beaded ribs distinguish E. marisrubri . At Ras al-Hadd E. omanensis is sympatric with rare migrants of E. leucosticta .
Sequence data from the mitochondrial COI gene show that the sister species of E. omanensis is E. subnodosa from the Red Sea (Williams & Reid 2004; see Remarks on E. subnodosa ).
Separation from members of the conchologically similar E. malaccana group is discussed in the Remarks on the E. natalensis group and on E. malaccana (Fig. 27). Among the other members of the E. natalensis group, the shells of E. omanensis are most similar to those of E. natalensis ( Fig. 13 View FIGURE 13 ; see Remarks on E. natalensis ) although the two are not known to occur sympatrically. There is some variation in the proportions of the shell of E. omanensis , and in the development of a third row of nodules towards the suture, but no entirely smooth forms are known. Three samples have been seen from southern Somalia (see Range above) and in these the shells have more pointed nodules ( Fig. 17A, B View FIGURE 17 ) and thus the ‘prickly’ appearance of E. natalensis . These samples are geographically closer to E. natalensis than to the nearest records of E. omanensis , but to the south they are isolated from E. natalensis by 800 km of unsuitable sedimentary coastline. They are identified as E. omanensis because of their narrow profile, 3–4 ribs crossing the peripheral nodules, and 15–16 spiral ribs on the last whorl. Further collecting is required to establish whether, as expected, this species occurs on the rocky shores of the northern and northwestern coasts of Somalia.



No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |