
Callomyia proxima Johnson
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4111.5.1 |
|
publication LSID |
lsid:zoobank.org:pub:1286E111-8C60-47AB-B2A2-36D3BFB6CA3F |
|
DOI |
https://doi.org/10.5281/zenodo.5621990 |
|
persistent identifier |
https://treatment.plazi.org/id/3C7A0266-7362-9727-C2B1-2E3A1845FE29 |
|
treatment provided by |
Plazi |
|
scientific name |
Callomyia proxima Johnson |
| status |
|
Callomyia proxima Johnson View in CoL
( Figs 12 View FIGURES 9 − 14 , 22 View FIGURES 19 − 24 , 30 View FIGURES 29 − 32 , 38 View FIGURES 37 − 40 , 49, 51 View FIGURES 49 − 54 , 62 View FIGURES 59 − 62 , 75 View FIGURE 75 )
Callomyia proxima Johnson, 1916: 32 View in CoL . Type locality: Mt. Washington, New Hampshire, USA.
Callomyia liardia Kessel & Buegler, 1972: 262 . Type locality: Liard Hot Springs, British Columbia, Canada. syn. nov.
Diagnosis. This apparently disjunct eastern and western Nearctic species is characterized by its abdominal colour patterns, mid tibial setae, and male terminalia with a bifid surstylus that has long outer and inner processes and a bifid hypandrial process. The female is distinguished by a long median dorsal seta on the mid tibia and a distinctive abdominal colour pattern (yellow on tergites 1 and 2, silver-yellow or silver-blue on posterior portion of tergites 3 and 4—all interrupted by a median dorsal dark band—and tergite 6 entirely silver-blue). The male is distinguished by a median anterodorsal seta that is usually present above a median dorsal seta on the mid tibia, a dark brown to velvety black abdomen usually with posterolateral or posteroventral silver-grey markings on tergites 3 and 4, and features of the terminalia mentioned above. The male of C. proxima is similar to C. velutina from eastern and western North America, but differs by the silver-grey abdominal markings, a median anterodorsal seta on the mid tibia, and terminalia, as indicated in the key to species. The male terminalia of C. proxima are also very similar to the southwestern Nearctic species C. bertae , with differences indicated under that species.
Description. Male ( Figs 12 View FIGURES 9 − 14 , 22 View FIGURES 19 − 24 ). Body length 3.4–4.1 mm; wing length 3.25–3.8 mm. Head silver-black; mouthparts including palpus brownish-yellow; antenna with scape, pedicel, first flagellomere and arista dark brown. Antenna with first flagellomere short-oval (as in Fig. 41 View FIGURES 41 − 44 ).
Thorax mainly velvety black, dark brown in some specimens; propleuron, mesopleuron, metapleuron, mediotergite and laterotergite silver-grey dusted. Scutum with 6 notopleural setae.
Fore leg and mid leg light brown, hind leg dark brown, apex of femur and base of tibiae brownish-yellow. Mid tibia with median anterodorsal seta present in most specimens, median dorsal seta present ( Fig. 49 View FIGURES 49 − 54 ); base of hind femur with long thin posteroventral seta (as in Fig. 54 View FIGURES 49 − 54 ). Hind tarsomere 1 long, slightly narrower than apical width of hind tibia, length approximately 3X width.
Wing hyaline with cell sc faintly yellow, whitish in some specimens. Halter with stem brown to dark brown, knob yellow to orange.
Abdomen mainly dark brown to velvety black with posterolateral to posteroventral silver-grey markings on tergites 3 and 4 in most specimens; tergite 7 entirely brown to grey; sternites light brown, sternite 8 brown to grey.
Terminalia ( Fig. 62 View FIGURES 59 − 62 ) with epandrium grey; surstylus dark brown to black; hypandrium and cercus brown. Epandrium with minute ventral lobe, barely apparent; apical process short, broad at apex. Surstylus bifid, with long outer and inner process, both rounded at apex; outer process broader and longer; inner process narrower and shorter. Hypandrium with moderately long apical process; process bifid, with 2 short apical projections; dorsal projection narrow and longer, ventral projection broad and shorter. Postgonite long and moderately wide, somewhat truncated and slightly pointed at apex. Phallus sharply hooked at apex. Cercus short.
Female ( Figs 30 View FIGURES 29 − 32 , 38 View FIGURES 37 − 40 ). Body length 3.35–3.85 mm; wing length 3.35–3.7 mm. Head silver-blue with occiput silver-black; mouthparts brownish-yellow with palpus yellow to orange; antenna with scape and pedicel brown, first flagellomere and arista brown to dark brown. Antenna with first flagellomere short-oval (as in Fig. 43 View FIGURES 41 − 44 ).
Thorax black to velvety black with silver-blue markings on entire lateral portion of presutural scutum, notopleuron, most of postsutural scutum and postalar callus; propleuron, mesopleuron, metapleuron, mediotergite and laterotergite silver-blue dusted; postpronotal lobe and supra-alar area of scutum brownish-yellow. Scutum with 2–3 presutural intra-alar setae.
Fore leg and mid leg yellow to yellowish-brown with tarsomeres 3–5 brown to dark brown; hind leg darker with apex of femur, tibia and tarsomeres dark brown. Mid tibia with long median dorsal seta present ( Fig. 51 View FIGURES 49 − 54 ).
Wing hyaline with cell sc bluish-white in some specimens. Halter yellow.
Abdomen dark brown to black with tergites 1 and 2 yellow, both interrupted by median dorsal dark band; posterior silver-yellow or silver-blue markings on tergites 3 and 4, both interrupted by median dorsal dark band; tergite 6 entirely silver-blue; sternites white to pale yellow.
Terminalia with segment 8 and epiproct yellowish-brown to brown, silver-grey dusted; hypoproct and cercus brownish-yellow.
Type material. Callomyia proxima Johnson , LECTOTYPE (designated by Kessel & Buegler 1972), ♀ labelled: “Mt Washington/ betw. 2 & 3 mile [hand written] N H [New Hampshire]/ VII.16.1915 [hand written]; “ Paratype / No. [red label]; “C.W. Johnson/ Collector; “M.C.Z./ Paratype / 26892 [red label]; “ MCZ ENT / 0 0 304201 ( MCZ). PARALECTOTYPE: USA: NEW HAMPSHIRE: Mount Washington, 2–3 mi. post Carriage Road, 16.vii.1915, C. W. Johnson (1 ♀, MCZ).
Callomyia liardia Kessel & Buegler , HOLOTYPE, ♂ labelled: “Liard Hot/ Sprs.. B.C./ VII.26.62/ E.L.Kessel; “ HOLOTYPE / Callomyia / liardia / Kessel &/ Buegler [pink label]; “California Academy/ of Sciences/ Type No. 11265 [dissected] ( CAS). PARATYPES: CANADA: BRITISH COLUMBIA: same data as holotype (1 ♂, CAS); Kleanza Creek, 12 mi. E. of Terrace, 13.viii.1965, E.L. Kessel (1 ♂, CAS); Liard Hot Springs, Mile Post 496, Alaska Hwy, 8.viii.1959, E.L. Kessel (1 ♂, CAS); USA: ALASKA: 27 miles S. of Livengood, 28.vi.1962, E.L. Kessel (2 ♂, CAS); Mile Post 1231, E. of Tetlin Junction, Alaska Highway, 14.vii.1962, E.L. Kessel (1 ♂, CAS); Mile Post 1277, E. of Tetlin Junction, Alaska Highway, 14.vii.1962, E.L. Kessel (1 ♂, CAS); Peters Creek Camp Ground, 1.vii.1962, E.L. Kessel (1 ♂, CAS); Richardson Highway, 21 miles N. of Delta Junction, 29.vi.1962, E.L. Kessel (1 ♂, CAS); Salcha River Camp Ground, 28.vi.1962, E.L. Kessel (1 ♂, CAS); Spenard, 24.viii.1957, E.L. Kessel (1 ♂, CAS), same except 25.viii.1957 (1 ♂, CAS), same except 17.viii.1959 (1 ♂, CAS), same except 25.viii.1959 (1 ♂, CAS), same except 26.viii.1959 (1 ♂, CAS); 9 mi. E. of Valdez, 12.vii.1962, E.L. Kessel (1 ♂, CAS).
Additional material examined. CANADA: BRITISH COLUMBIA: Kleanza Creek, 12 mi. E. of Terrace, 14.viii.1965, E.L. Kessel (4 ♀, CAS); Liard Hot Springs, Mile Post 496, Alaska Hwy, 2.ix.1957, E.L. Kessel (1 ♂, CAS), same except 8.viii.1959 (2 ♀, CAS); Liard Hot Springs, 26.vii.1962, E.L. Kessel (1 ♂, CAS); Mile Post 104, Alaska Highway, 5.viii.1957, E.L. Kessel (3 ♀, 3 ♂, CAS), same except 5.viii.1959 (4 ♂, CAS); Mile Post 350, Alaska Highway, 6.viii.1957, E.L. Kessel (1 ♀, CAS); Mount Thornhill, near Terrace, 30.vii.1960, W.R. Richards (2 ♂, CNC); Whiskers Province Camp Ground, Hart Hwy., 29.vii.1962, E.L. Kessel (1 ♂, CAS); ONTARIO: Bruce Peninsula NP, Cameron Lake Road, 4.vii.1998, 4512.5N8133.5W, K.N. Barber, sweep veg./grasses, mixed forest (1 ♂, DEBU); Bruce Peninsula NP, Bruce Trail nr. Crane Lake, 29.viii.1999, S.A. Marshall (1 ♂, DEBU); Bruce Peninsula NP, Dunks Bay, 20.vii.1996, S.A. Marshall (1 ♂, DEBU), same except 27.vii.1997 (1 ♂, DEBU), same except 4.vii.1998, 4514’N8138’W, DEBU 0 0 0 75059 (1 ♀, 5 ♂, DEBU), same except 1-2.viii.1999 (1 ♀, DEBU); Crow Lake, Marmora area, 18.viii.1959, L.K. Smith (1 ♀, CNC); QUEBEC: Gatineau Park, King Mountain/Black Lake Trail, 29.viii.2012, 4529’20”N7551’53”W, H.J. Cumming, JSS26016 (1 ♂, LEMQ); Gatineau Park, Waterfall Trail, 3.ix.2012, 4528’42”N7551’06”W, H.J. Cumming & S.H. Cumming, JSS26018, JSS26017 (1 ♀, 1 ♂, LEMQ), same except 24.viii.2013, J.M. Cumming, CNC 287574 (1 ♀, CNC); Old Chelsea, 31.viii.1958, J.R. Vockeroth (1 ♀, CNC); Old Chelsea, Summit King Mountain, 1150’, Gatineau Park, 9.viii.1985, J.R. Vockeroth, CNC DIPTERA 192199 (1 ♀, CNC); USA: ALASKA: Anchorage, 19.vii.1921, J.M. Aldrich (1 ♀, 3 ♂, USNM), same except 21.vii.1921 (5 ♂, USNM), same except 20.vii.1951, R.S. Bigelow (2 ♀, CNC); 6 mi. W. of Circle city, 23.vi.1962, E.L. Kessel (1 ♂, CAS); Haines, 11.viii.1959, E.L. Kessel (1 ♀, CAS); Johnson Lake, 16 mi. S. of Soldatna, 5.vii.1962, E.L. Kessel (1 ♂, CAS); 4 mi. S. of Livengood, 27.vi.1962, E.L. Kessel (1 ♂, CAS); 27 miles S. of Livengood, 28.vi.1962, E.L. Kessel (1 ♂, CAS); Mile Post 1231, E. of Tetlin Junction, Alaska Highway, 14.vii.1962, E.L. Kessel (3 ♀, 1 ♂, CAS); Mile Post 1277, E. of Tetlin Junction, Alaska Highway, 14.vii.1962, E.L. Kessel (2 ♂, CAS); Mosquito Lake, 27 miles N. of Haines, 17.vii.1962, E.L. Kessel (1 ♂, CAS); Richardson Highway, 21 miles N. of Delta Junction, 29.vi.1962, E.L. Kessel (3 ♂, CAS); Soldatna, 5.vii.1962, E.L. Kessel (1 ♂, CAS); Spenard, 24.viii.1957, E.L. Kessel (1 ♂, CAS), same except 17.viii.1959 (2 ♀, 8 ♂, CAS), same except 19.viii.1959 (2 ♂, CAS), same except 7.vii.1962, E.L. Kessel (2 ♂, CAS); 9 mi. E. of Valdez, 12.vii.1962, E.L. Kessel (1 ♀, 1 ♂, CAS); Wasilla, 10.viii.1959, E.L. Kessel (1 ♂, CAS); MASSACHUSETTS: Chester, 5.viii.1914, C.W. Johnson (1 ♂, MCZ, paratype of C. velutina ); NEW HAMPSHIRE: Bretton Woods, 1.vii.1936, A.L. Melander (1 ♀, USNM); Mount Washington, 6.vii.1914, C.W. Johnson (1 ♂, MCZ, paratype of C. velutina ), same except betw. 1 & 2 mile, 24.vii.1915 (1 ♂, MCZ, paratype of C. velutina ); Mount Washington, Raymond Path, 28.vii.1915, C.W. Johnson (1 ♂, MCZ, paratype of C. velutina ); NEW YORK: Ithaca, vi, R.C. Shannon (1 ♂, USNM); St. Lawrence County, Cranberry L., 25.vi.1963, W.W. Wirth, swamp (1 ♂, USNM); Trumansburg, 19.vii.1983, D. Grimaldi (1 ♂, AMNH); NORTH CAROLINA: Macon County, Wayah Gap, 3500’, 10.viii.1957, J.G. Chillcott (1 ♀, CNC); UTAH: Cache County, W. Hodges Canyon, 14-18.vii.1978, Hanson & Knowlton (1 ♀, LACM); WEST VIRGINIA: Pocahontas County, Island Cpg, E. fork Greenbriar, 1.vii.1982, Mathis & Flint (1 ♂, USMN).
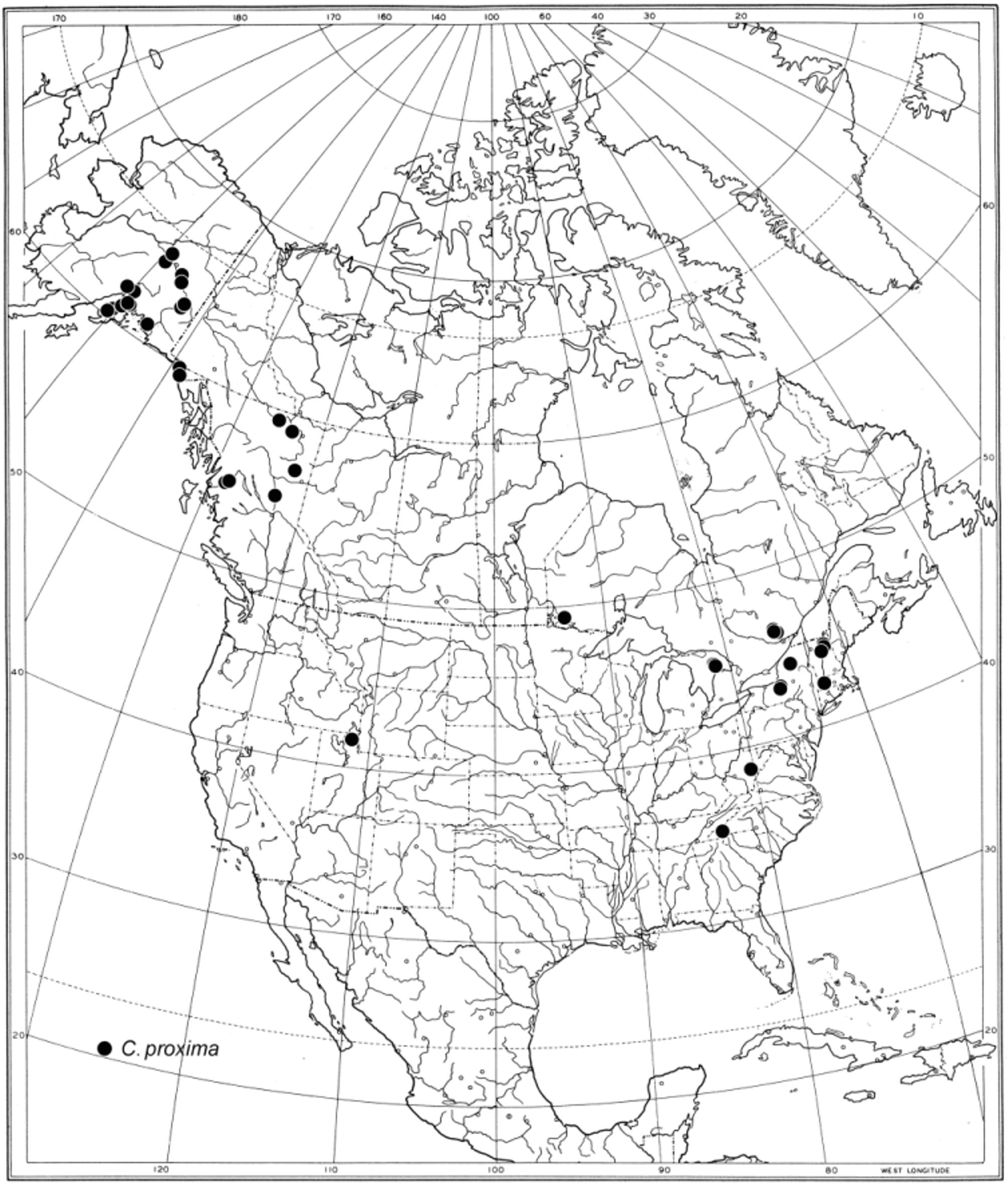
Geographical distribution and seasonal occurrence ( Fig. 75 View FIGURE 75 ). Callomyia proxima is currently distributed in both western North America (Alaska, British Columbia and Utah) and eastern North America (Ontario, Quebec, New York, New Hampshire, Massachusetts, West Virginia and North Carolina). Adults have been collected from late June to early September.
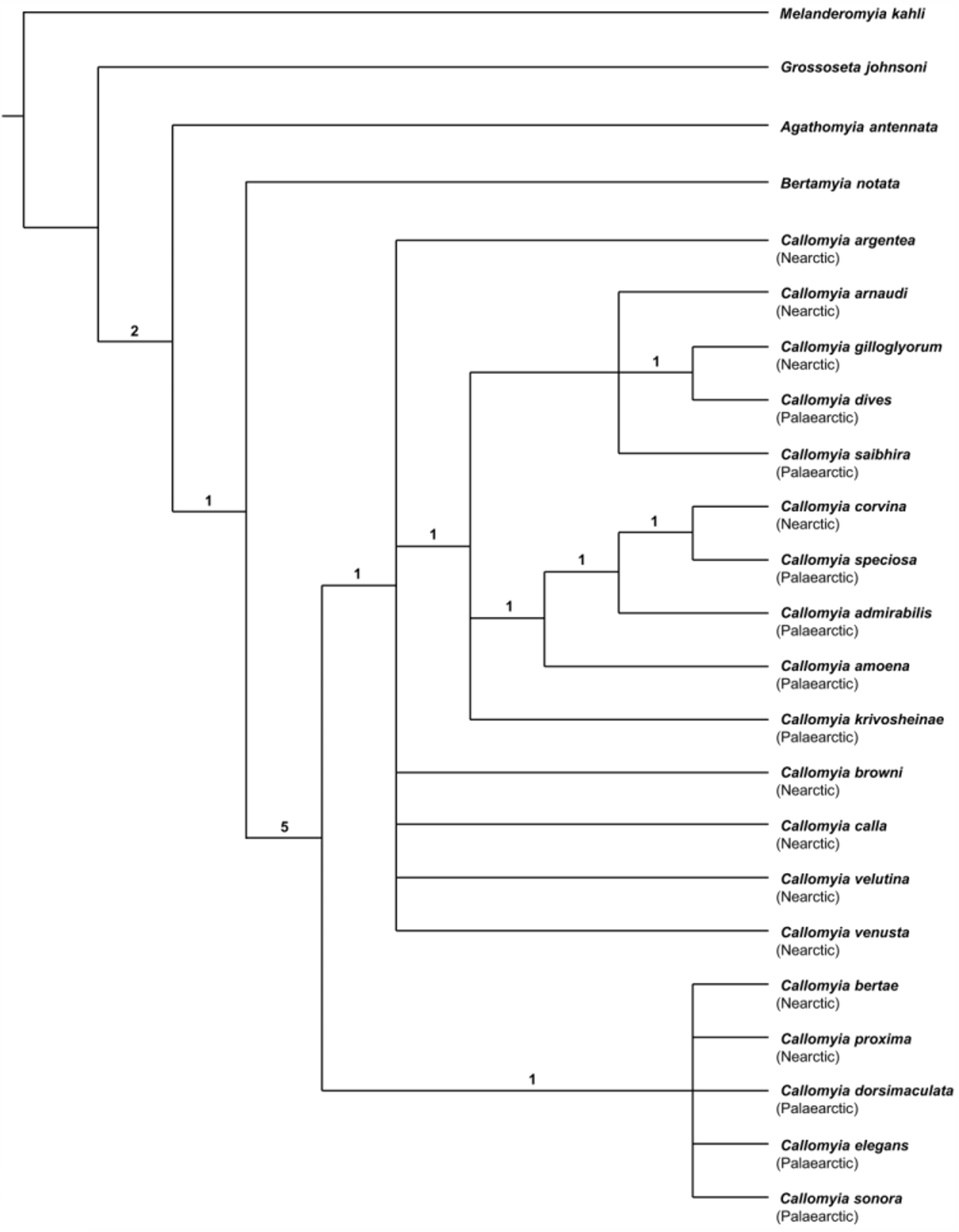
Remarks. Callomyia proxima is closely related to the Nearctic species C. bertae and the Palaearctic species C. dorsimaculata , C. elegans , and C. sonora , as shown in the strict consensus tree ( Fig. 79 View FIGURE 79 ). Within this clade, C. proxima is most similar to C. elegans because of similar male terminalia, setae on the mid tibia, and thoracic and abdominal colour patterns. However, it is more robust than C. elegans and its male terminalia differ by having a wider ventral projection on the hypandrial process and a shorter and wider postgonite.
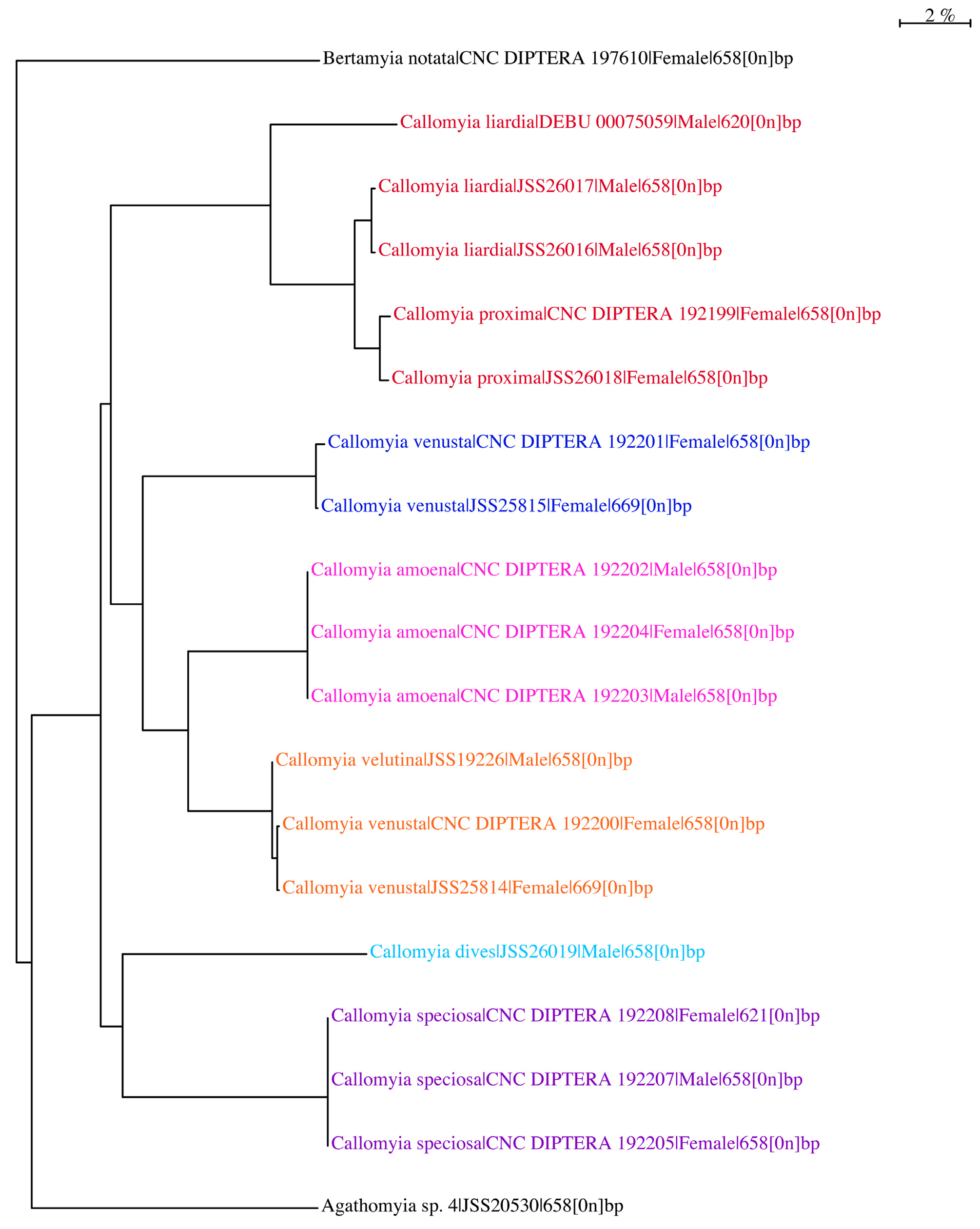
Two female specimens of C. proxima yielded COI barcode sequences (CNC DIPTERA 192199 and JSS26018) as did two male specimens, initially identified as C. liardia (JSS26016 and JSS26017) ( Table 1 View TABLE 1 ). In the Neighbourjoining tree ( Fig. 78 View FIGURE 78 ), these four specimens clustered closely together (<2% genetic divergence), suggesting that C. proxima and C. liardia are conspecific. A third male specimen of C. liardia yielded a barcode (DEBU 00075059) ( Table 1 View TABLE 1 ) and also clustered with the four specimens mentioned above, but had 6% genetic divergence from these specimens. Although this specimen has greater genetic divergence from the others, it appears to be morphologically identical to C. liardia (= C. proxima ). Only 620 bp of COI were sequenced for that specimen (658 bp were sequenced from the others), which may explain the higher divergence from the rest of the C. proxima cluster.
Callomyia proxima View in CoL was described from females only ( Johnson 1916) and Callomyia liardia from males only ( Kessel & Buegler 1972). Both species are considered to be conspecific because they have been collected together from multiple localities, including both type localities. In addition, DNA barcode data support the new synonymy proposed.
Kessel & Buegler (1972) considered four of the seven paratypes of C. velutina to be conspecific with C. liardia (= C. proxima ). Three of these four paratypes are listed under “ Type Material of C. velutina and have been added to “Additional material examined of C. proxima , above. The fourth paratype was not examined. Another paratype of C. velutina (from Raymond Path, Mount Washington, New Hampshire) was also discovered to be conspecific with C. proxima and is listed under “ Type Material of C. velutina .







| MCZ |
Museum of Comparative Zoology |
| ENT |
Ministry of Natural Resources |
| NEW |
University of Newcastle |
| CAS |
California Academy of Sciences |
| CNC |
Canadian National Collection of Insects, Arachnids, and Nematodes |
| DEBU |
Ontario Insect Collection, University of Guelph |
| LEMQ |
McGill University, Lyman Entomological Museum |
| USNM |
Smithsonian Institution, National Museum of Natural History |
| AMNH |
American Museum of Natural History |
| LACM |
Natural History Museum of Los Angeles County |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Callomyia proxima Johnson
| Cumming, Heather J. & Wheeler, Terry A. 2016 |
Callomyia liardia
| Kessel 1972: 262 |
Callomyia proxima
| Johnson 1916: 32 |