
Erebussau profundus, Bezerra & Pape & Hauquier & Vanreusel, 2021
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4903.4.4 |
|
publication LSID |
lsid:zoobank.org:pub:6BFD34BE-F34B-4E77-99ED-9B833006B267 |
|
DOI |
https://doi.org/10.5281/zenodo.4431405 |
|
persistent identifier |
https://treatment.plazi.org/id/3D096322-D433-564E-FF5B-FCBD7815FD9A |
|
treatment provided by |
Plazi |
|
scientific name |
Erebussau profundus |
| status |
sp. nov. |
Erebussau profundus View in CoL sp. nov.
( Figs 5 View FIGURE 5 , 6 View FIGURE 6 and 7 View FIGURE 7 ; Tables 1 and 2)
Material examined for description: One male, three females and one juvenile. One holotype male (Inventory No. UGMD 104402 View Materials ) and one paratype female (Inventory No. UGMD 104403 View Materials ) are deposited in the Zoology Museum of Ghent University—Museum voor Dierkunde ( UGMD) .
Type habitat and locality: Deep-sea with sediment grain size dominated by silt, containing nodules. Male holotype was found in a sample from the GSR license area in the CCFZ during expedition SO239, Station 141, Core 1, 0–1 cm, at an average of 4509 m depth .
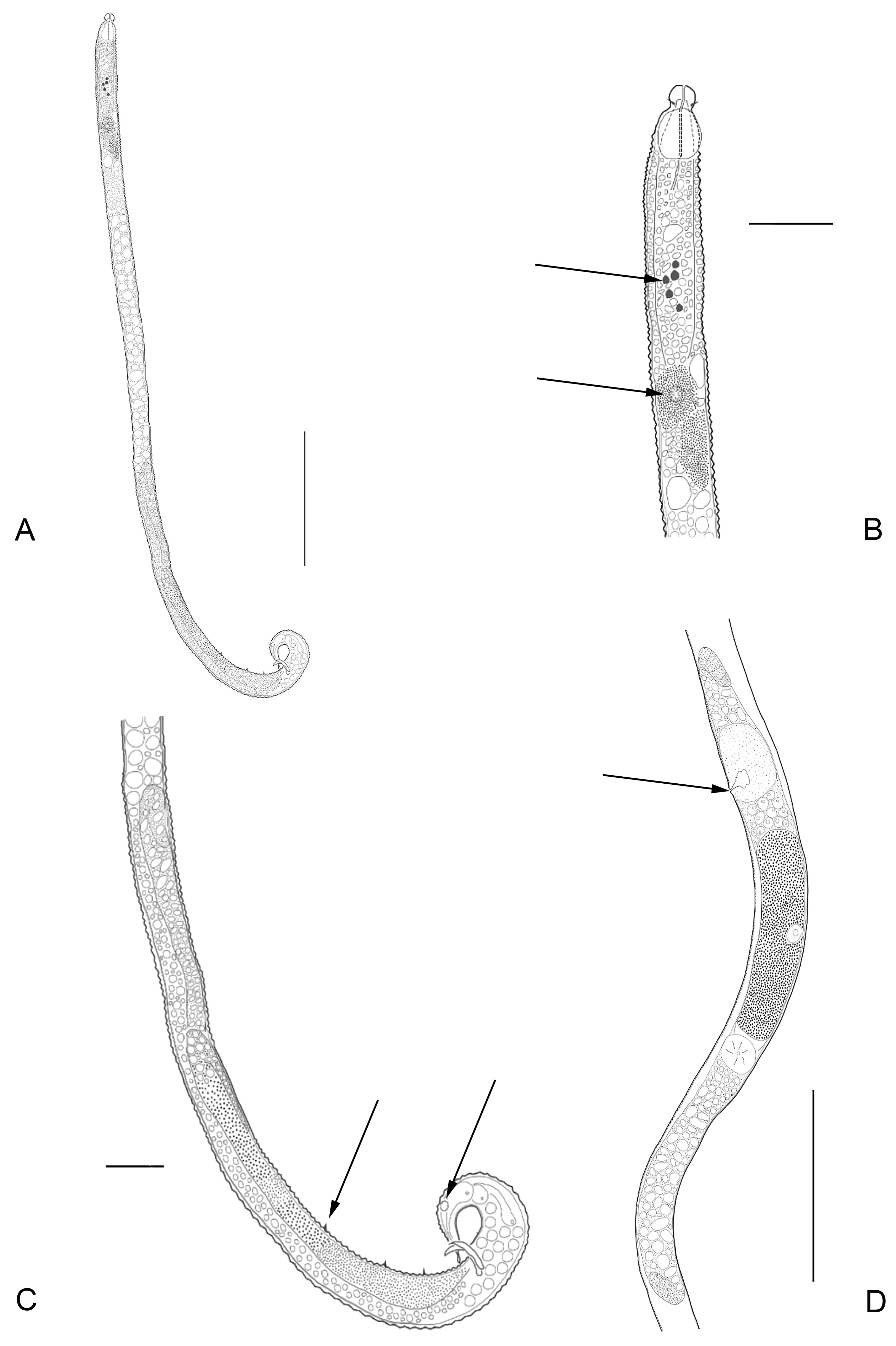
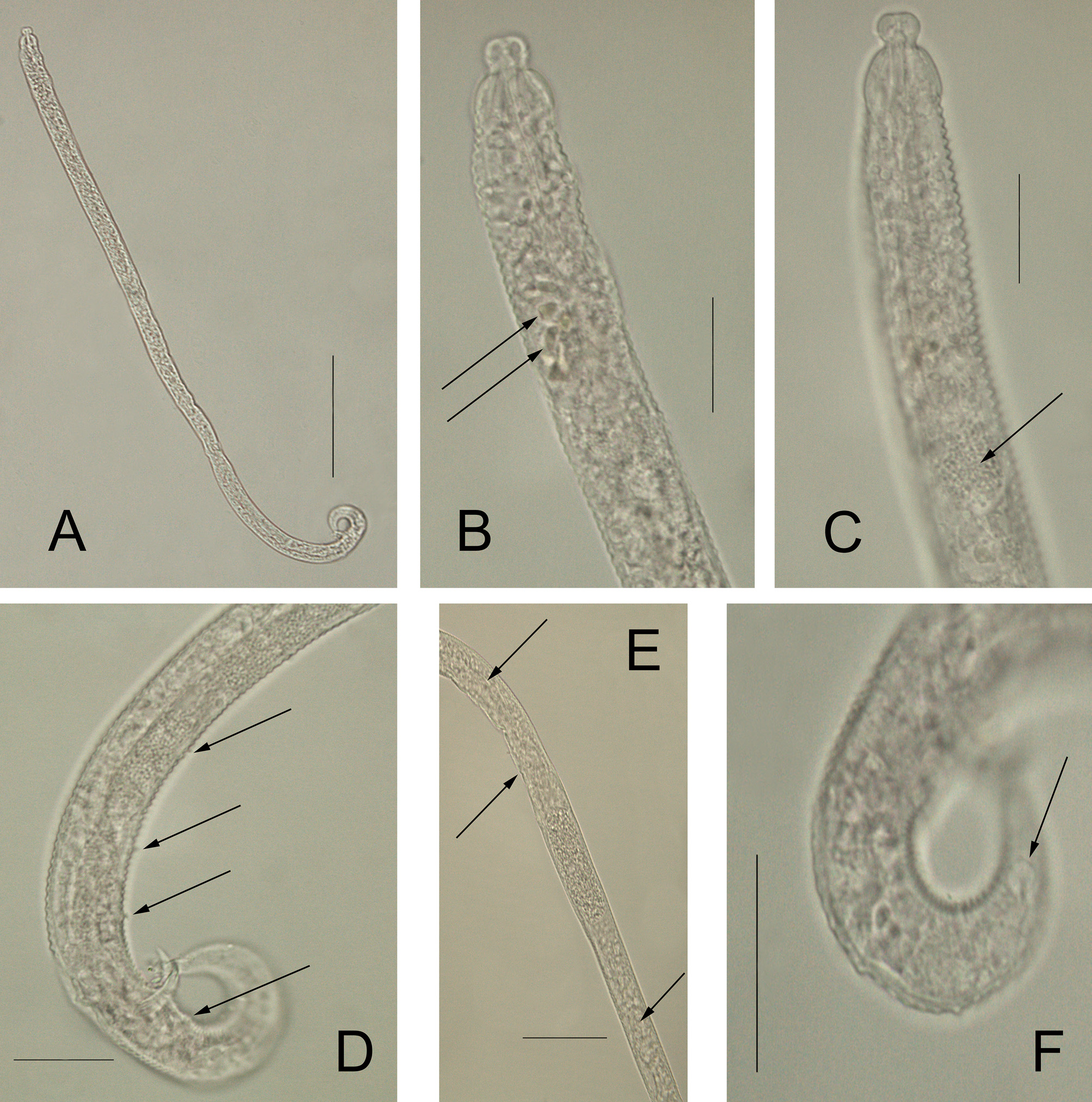
Description: Males: Body elongated, strongly annulated until mid-body. Posterior end also strongly annulated. Annulations of the anterior part of the body with triangular smooth outline. No somatic setae are visible. Cephalic region clearly offset followed by a narrow constriction about 50% narrower than the section behind this constriction. Four cephalic setae are present. Amphids vesicular, with a blistered amphidial fovea. Buccal cavity small and unarmed. Pharynx surrounded by many glands. Many pigment spots present, located at the end of the pharynx and beginning of intestine. Pseudocoelomocytes present, at the end of the pharynx and beginning of the intestine. Testes reflexed. Anterior testis well developed, posterior one much smaller. Spicules just slightly curved. Gubernaculum with caudal apophysis. Three setae inserted on elevations of the body cuticle, present as pre-cloacal supplements, plus one additional post-cloacal supplement. Small phasmata-like structures located laterally at the last posterior third of the tail region, on each side of the body. These structures can easily be overlooked. Three caudal glands contained in the tail. Tail conical, long with spinneret.
Females: Similar to males in most aspects, although generally longer. No sexual dimorphism detected. Reproductive system didelphic, amphidelphic, with reflexed ovaries. Reproductive system with a clear posterior ovary and a smaller anterior ovary. Vulva with non-protruding lips, positioned close to the mid body, slightly posteriorly in most cases. No perivulvar glands observed. Vagina transverse. Uterus not well-distinguished, appearing to be uniform. Oviduct indistinct; ovaries antidromously with reflexed tips. Phasmata-like structures, laterally at the last posterior third of the tail region, on each side of the body, larger than in males. Female tail conical, longer than in males. Terminal spinneret with same morphology as in males but slightly elongated.
Juveniles: Similar to adults in most morphological aspects.
Etymology: Profundus (Latin, adjective class I) = deep, profound.
Differential diagnosis: Erebussau profundus sp. nov. closely resembles Erebussau tenebricosus nom. nov., comb. nov., but males of Erebussau profundus sp. nov. are on average 27% longer in total length. This difference is reduced to 16% when females of both species are compared. Pigment spots are more numerous in Erebussau profundus sp. nov. Five to seven pigment spots were observed, in contrast with two in Erebussau tenebricosus nom. nov., comb. nov.. Other distinct characteristics are the presence of pre- and post-cloacal supplements and the shape of the spicules. The spicules are just slightly curved and 51 % longer when compared to the clearly curved and smaller ones of Erebussau tenebricosus nom. nov., comb. nov. ( Table 2). The triangular outline shape of cuticle annulation at the anterior part of the bod of Erebussau profundus sp. nov. is less pronounced than that in Erebussau tenebricosus nom. nov., comb. nov. and extends further until mid-body, although this could be the result of movement, such as contraction of the longitudinal body muscles and friction with the sediment. These movements can cause rings to telescope more or less, proving a more triangular outline.
General remarks: Analyses of different specimens allowed us to observe intra- and interspecific differences during the new denomination of Erebussau nom. nov., resulting in an update of the description of the family Meyliidae .
Blister-like amphids are a synapomorphy for Desmoscolecida ( Lorenzen, 1981, 1994) and the previous descriptions mentioned that members of Meyliidae differ from the other desmoscolecids by the vesicular fovea showing an inner spiral structure which is not the case for Erebussau nom. nov. Bussau (1993) described a blistered heartshaped amphid vesicular structure, this was not visible in the analyzed Erebussau nom. nov. specimens.
Another finding was the presence of pigment spots. Pigment spots with different sizes and shapes were found in the specimens of both species of Erebussau . The absence of lens-like structures refrains us from the use of the term ocelli. Bussau (1993) observed in Tricoma and Desmoscolex that the pigment spots were located at the level of the ampulla-like widening of the fusus amphidialis and hypothesized that the pigment spots are in fact concentrated secretions from the amphidial sheath cell, explaining their differences in size and shape among specimens. Lorenzen (1994) described pigment spots as resembling droplets of fluid and noted that their presence appears independent from the biotope in which the animals occur, stating that they were not present in deep-sea species. However, Decraemer (1989) found pigment spots in deep-sea Tricoma (Quadricoma) loricata specimens from the flat of Cap Vert (tropical Atlantic Ocean) revoking Lorenzen’s (1994) statement.
Phasmata were considered absent in Meyliidae , but phasmata-like structures are present in Erebussau nom. nov.
Pseudocoelomocytes are observed at the end of the pharynx, beginning of intestine, both in E. tenebricosus nom. nov., comb. nov. and E. profundus sp. nov., although they are more conspicuous in E. profundus sp. nov. To date, Erebussau comb. nov., together with Noffsingeria and Meylia vangoethemi , are the sole representatives of Meyliidae which possess pseudocoelomocytes.
The females of the described sibling species of Erebussau comb. nov. did not show striking features which can be used to separate the two species, but they could be distinguished by the sharp offset cephalic region, together with the greater length of E. profundus sp. nov. being 16 % longer on average, the triangular outline shape of cuticle annulation which extends further until mid-body and the well-developed anterior ovary observed in the two described specimens of E. profundus sp. nov.
| UGMD |
Zoology Museum of the University of Ghent |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.