
Madagascarhinus madagascariensis, Wesener, 2023
|
publication ID |
https://doi.org/ 10.11646/zootaxa.5278.1.9 |
|
publication LSID |
lsid:zoobank.org:pub:53B29C2D-C720-4F72-B10A-5DF61BA6812A |
|
DOI |
https://doi.org/10.5281/zenodo.7895190 |
|
persistent identifier |
https://treatment.plazi.org/id/3D6B87F2-FFEE-FFB1-DDBA-8CF4FEB13DB1 |
|
treatment provided by |
Plazi |
|
scientific name |
Madagascarhinus madagascariensis |
| status |
sp. nov. |
Madagascarhinus madagascariensis View in CoL n. sp.
Figures 1A View FIGURE 1 , 2 View FIGURE 2 and 3 View FIGURE 3
Material examined: Holotype: ♁, MHNG Mad 89/21; Madagascar, Province Antananarivo, Ankaratra massif, station Forestière Manjakatompo , près du sommet du Anosirivo, forêt primaire, prélèvement de sol dans une vieille souche, 1980 m; 26.xi.1989, leg. B. Hauser, extraction Berlese à Genève.
Paratypes: 1 ♁, 2 ♀♀, MHNG , same data as previous; 1 ♁, ZFMK MYR12534 View Materials , same data as previous, utilized for SEM .
Etymology: The new species is named after the type locality. Adjective.
Diagnosis: Long, slender, whitish-beige siphonorhinid without paranota. Up to 11 mm long with 62 tergites. Antennomere 2 as long as 3 and 4 combined, antennomere 6 only slightly longer than and as wide as antennomere 5. Sensilla basiconica on antennomere 5 and 6 located in a field. Anterior gonopod with seven podomeres, typical for the family, posterior gonopod with seven podomeres, podomere 7 apically with two bifurcated, apically thickening branches, basally with a spiniform process, typical for the genus. Differs from M. andasibensis n. sp. in the wider head (wider than long in M. madagascariensis n. sp., longer than wide in M. andasibensis n. sp.), the presence of a much larger lower branch of the claw at the anterior legs, reaching 2/3 of the length of the claw (1/3 of the claw in M. andasibensis n. sp.) at leg 1 and being even larger than the claw in leg 2, in the anterior and posterior gonopods possessing seven podomeres (six in M. andasibensis n. sp.), and the last podomere of the anterior gonopod being shorter and less strongly elongated than in M. andasibensis n. sp.
Description:
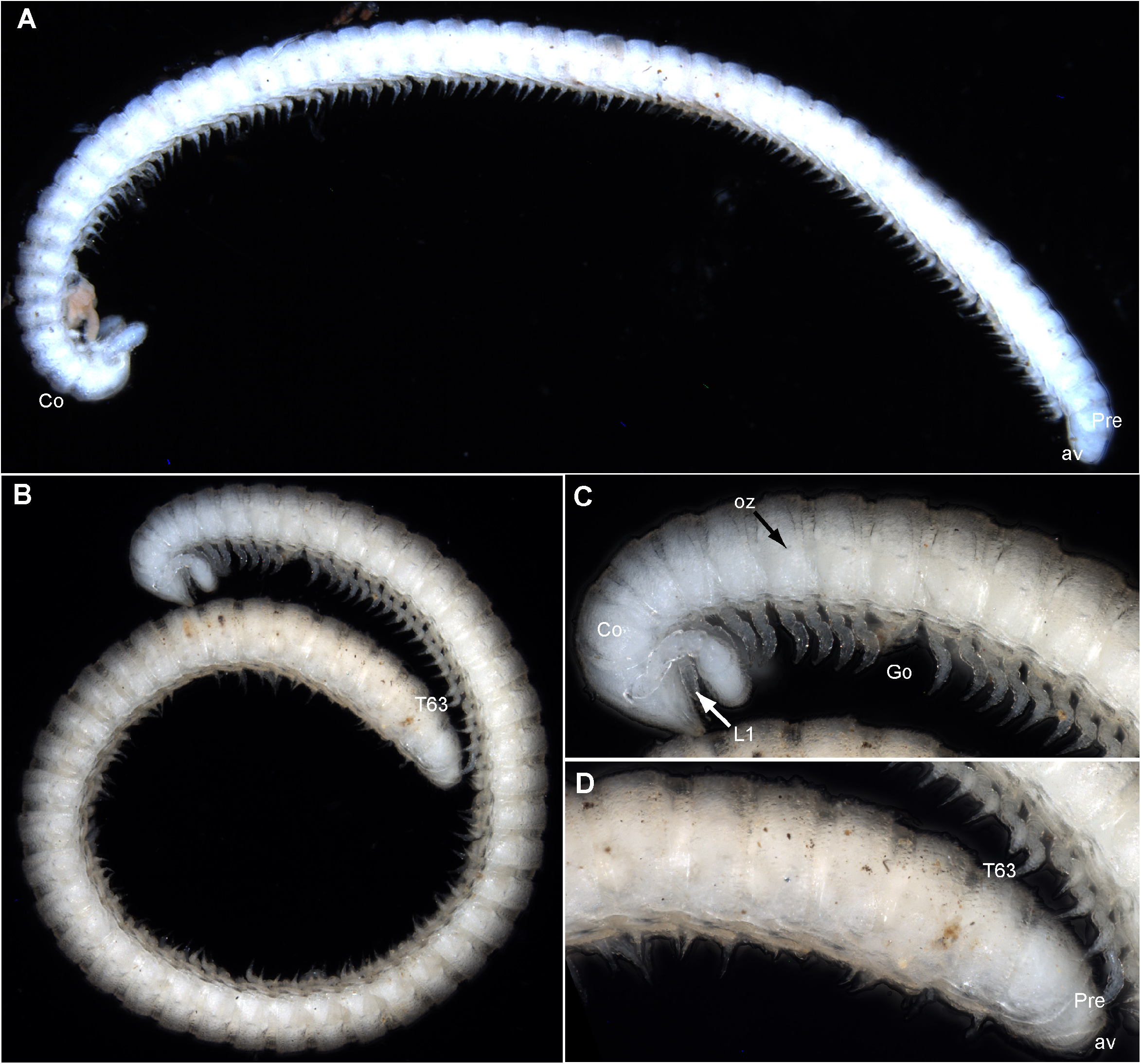
Measurements: all specimens of equal width, approximately 0.5 mm, holotype male with 49+1 tergites, ~ 8.5 mm long. Largest paratype male 61+1 tergites, ~ 11 mm long, smaller paratype male 27+1 tergites, ~ 4 mm long, females with 26+1 tergites, ~ 4 mm long and 19+1 tergites, ~ 3 mm long, respectively. Two males orange-whitish ( Fig. 1A View FIGURE 1 ), third male and females white (bleached?).
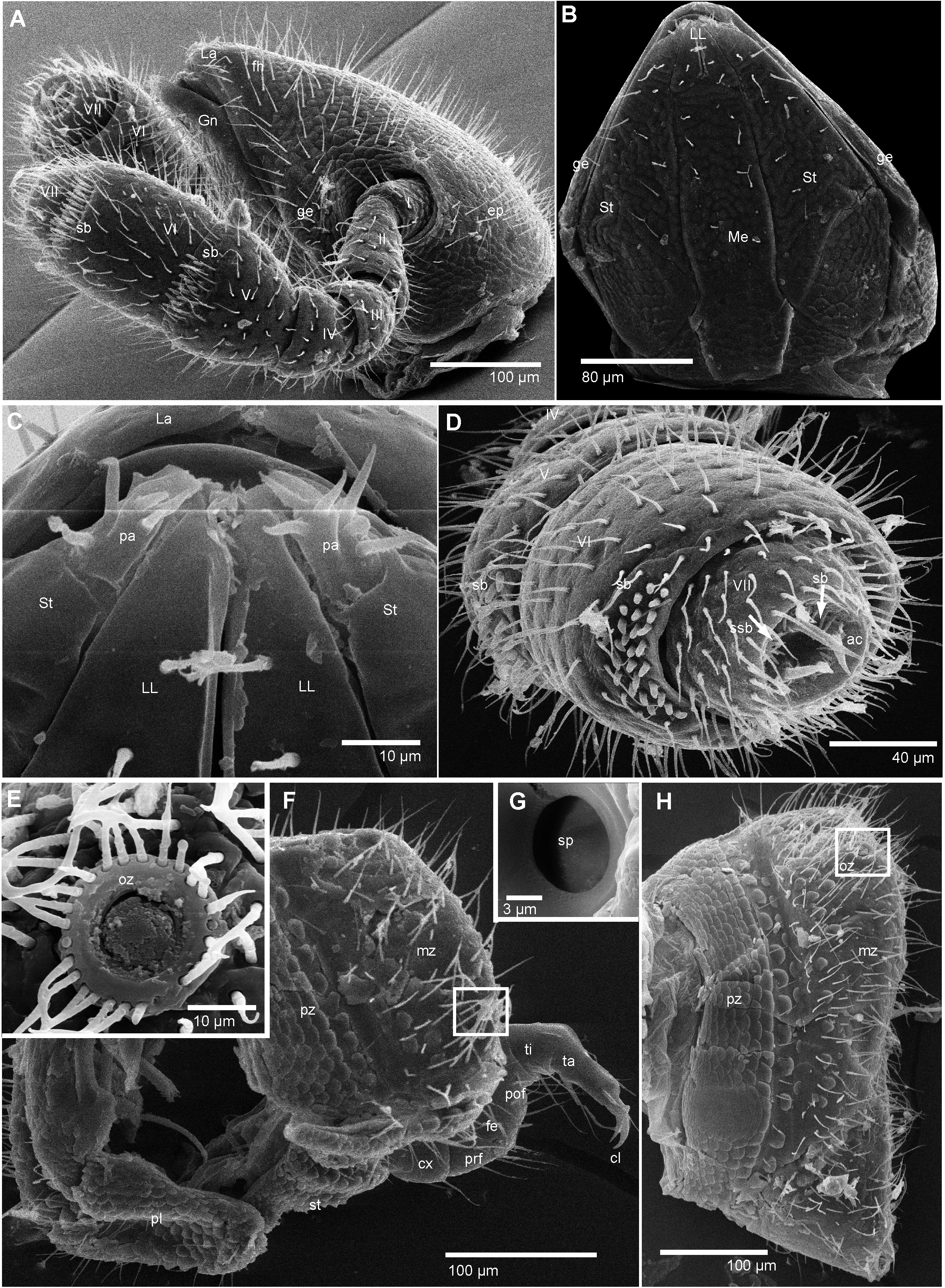
Head: pear shaped, densely setose, slightly wider than long ( Figs 2A, B View FIGURE 2 ). Gnathochilarium consisting of well distinguishable plates: mentum, stipes, lamellae linguales and a pair of lateral palpi origination from the stipes ( Figs 2B, C View FIGURE 2 ). Rudimentary palps as wide as lamellae linguales, with 4–7 sensorial cones ( Fig. 2C View FIGURE 2 ). Lamellae linguales towards one another with a sclerotized ledge, separated by a short gap ( Fig. 2C View FIGURE 2 ). Antennae elbowed between antennomeres 2 and 3, antennomere 2 elongated, narrower than other antennomeres, as long as 3 and 4 combined ( Fig. 2A View FIGURE 2 ). Antennomere 6 cylindrical, slightly longer but as wide as 5th. Antennomeres 5 and 6 latero-apically with rows of sensilla basiconica, 2 rows on 5th and 3 rows on 6th. Disc with four apical cones standing in a single field, a field of 5–10 sensilla basiconica as well as a semi-ring of isolated spiniform sensillae ( Fig. 2D View FIGURE 2 ).
Body-rings: collum slightly longer than following tergites, not overlapping head. Body rings clearly separated into tergite, pleurite and two sternites ( Fig. 2F View FIGURE 2 ). Tergites without paranota, ozopore located halfway between ventral and dorsal side, relatively close to the posterior margin, surrounded by a single dense row of setae ( Fig. 2E View FIGURE 2 ). Prozonites covered with scales, metazonites covered by long setae as well as large, well-rounded or spiny sclerotized spines ( Figs 2F, H View FIGURE 2 ). Pleurites separated into prozonite and metazonite, covered with scales like the prozonite of the tergite ( Fig. 2F View FIGURE 2 ). Sternites glabrous but covered with strong scales like the pleurites ( Fig. 2F View FIGURE 2 ), paired stigma openings covered by a plate ( Fig. 2G View FIGURE 2 ).
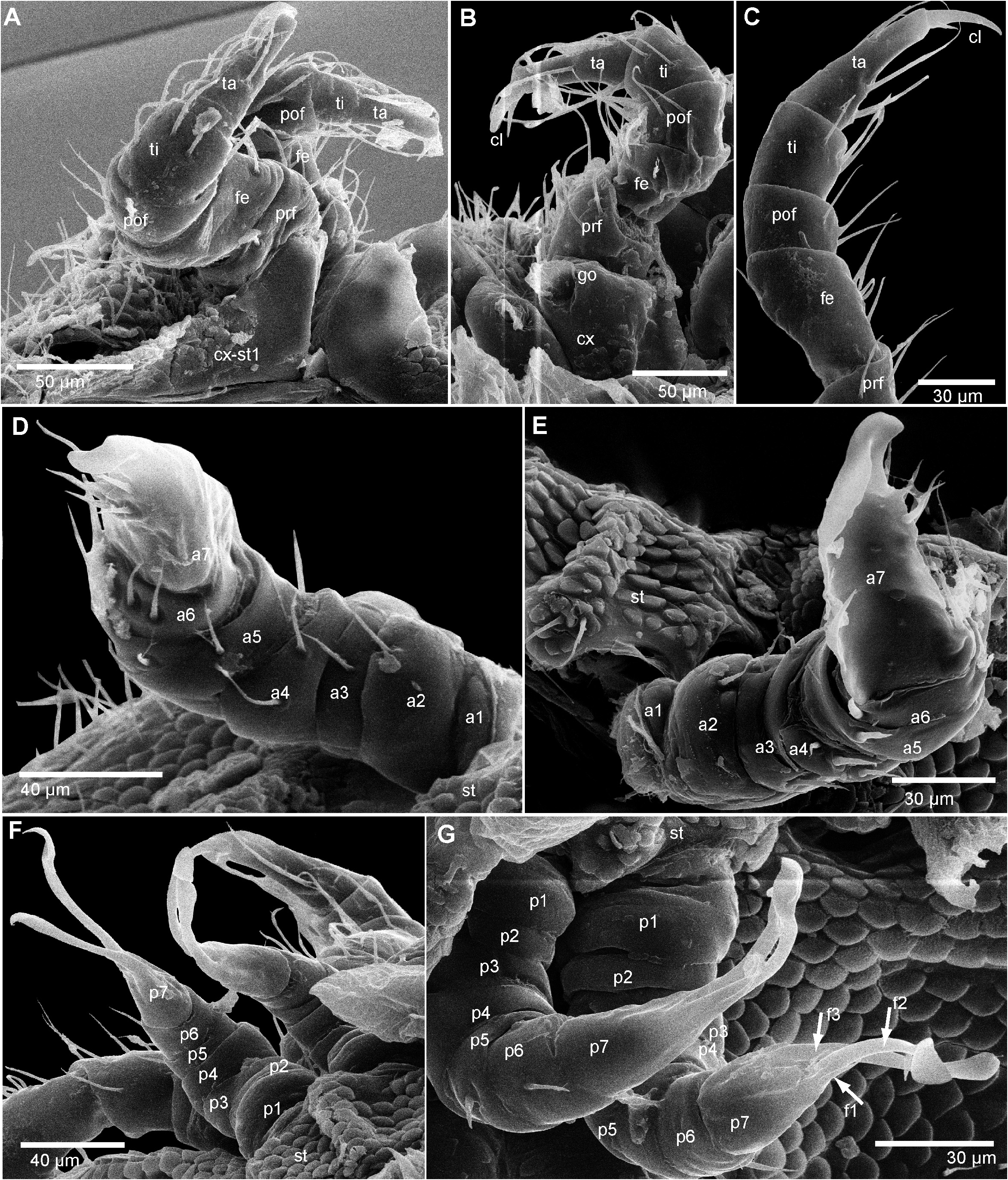
Legs: first leg pair larger and slightly thickened compared to following legs ( Fig. 3A View FIGURE 3 ). Triangular coxa fused to sternite. Second leg pair with greatly enlarged apical spine, surpassing length of claw ( Fig. 3B View FIGURE 3 ). First two leg pairs without trochanter or coxal pores, both trochanter and coxal pores starting at leg pair 3. Apical claw getting successively shorter in more posterior legs. Midbody leg ( Fig. 3C View FIGURE 3 ) with elongated femur, 1.5 times longer than wide, as long as postfemur and tibia combined. Each podomere at mesal margin with several long setae. Tarsus slender, apically tapering, 4 times longer than wide.
Telson: densely setose, shape of prenaal ring inconspicuous; anal valves well-developed; subanal scale small ( Fig. 1A View FIGURE 1 ).
Male sexual characters: gonopore inconspicuous hole with slightly projecting membranous part ( Fig. 3B View FIGURE 3 ).
Anterior gonopod sternite without any processes, inconspicuous. Anterior gonopod consisting of 7 podomeres, 1–6 being cylindrical, each carrying isolated setae ( Figs 3D, E View FIGURE 3 ). Podomere 7 strongly enlarged, as long as basal 3 or 4 podomeres combined, apically curved, forming a protection sheath for the posterior gonopod. apico-posterior edge with 5–8 long setae.
Posterior gonopod sternite inconspicuous ( Fig. 3F View FIGURE 3 ). Posterior gonopod consisting of 7 podomeres, 1–6 cylindrical. Podomere 7 with two long processes of equal length, being as long as podomeres 1–6 combined ( Figs 3F, G View FIGURE 3 ). In ventral (posterior) view with a third, setae-like process at mid-length, protruding up to half the length of longer processes. Longer processes forming a wide but thin, spoon-like apex.
Habitat: The forest of Manjakatompo is one of the last three (99% have been destroyed) indigenous high plateau forests in Madagascar. The forest was planted by humans a few centuries ago, when it was still connected to now no longer existing fragments of pristine highland vegetation. Manjakatompo harbors rare and endemic genera and species of mammals, but lacks other faunal elements (such as lemurs) most likely due to the artificial origin of the forest and the lack of any fruit-bearing trees ( Goodman et al. 1996).
Remarks: The same vial which contained the five Madagascarhinus madagascariensis n. sp. specimens also contained 3 Betscheuma sp. (Chordeumatida) and two indeterminable small-bodied Spirobolida (1 female and one juvenile).
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.