
Caecilia tentaculata Linnaeus, 1758
|
publication ID |
https://doi.org/ 10.5281/zenodo.203509 |
|
DOI |
https://doi.org/10.5281/zenodo.5635406 |
|
persistent identifier |
https://treatment.plazi.org/id/3E0BA842-4D55-2A44-BD96-D48D1D5AFB44 |
|
treatment provided by |
Plazi |
|
scientific name |
Caecilia tentaculata Linnaeus, 1758 |
| status |
|
Caecilia tentaculata Linnaeus, 1758 View in CoL
Caecilia tentaculata Linnaeus, 1758: 229 View in CoL ; Dunn, 1942: 480; Taylor, 1968: 555; Frost, 1985: 622; Lescure & Marty, 2000: 292; Coloma et al, 2004; Frost, 2008.
Diagnosis. Maximum known TL 850 mm ( Taylor, 1973). PA 112–131. SG 12–54. Eye visible; not covered by bone. Dermal scales present. SI between annulus 6 and 47, at most two rows of dermal scales per fold. Terminal shield present.
Description. TL 22.2–46.8 times (33.6 ± 5.3; n = 58) BW. Head slightly narrower than body. Snout projecting distinctly beyond mouth (e.g. 3.9 mm in MPEG 18160). Nuchal grooves may be distinct dorsally, laterally and ventrally, except third nuchal groove which ventrally is incomplete; second collar ventrally is fused with first primary annulus; a dorsal transverse groove is present on each collar, shorter and less distinct on first; some specimens have indistinct or partially distinct nuchal grooves so that collars are indistinct. Body subcylindrical, slightly wider than deep, (e.g. BW = 14.1 mm; BH = 13.4 mm in MPEG 18160). Primary annular grooves incomplete, interrupted dorsally and ventrally, except posteriormost primary grooves (at most eight, 4.9 ± 1.7; n = 47) that are complete; from general region where SG appear, to level of vent, PA dorsally complete. At most five secondary grooves complete (2.4 ± 1.3; n = 69). Vent transverse, may be slightly irregular; AD 10–17 (13.2 ± 6.2; n = 49); generally approximately similar number in posterior and anterior edge of vent (e.g. eight anterior, eight posterior in MPEG 23542). Paired anal papillae may be present in both juveniles and adults; found in one of two juveniles, 31 of 42 males and 5 of 25 females. Unsegmented terminal shield posterior of vent; in some specimens depressed. SI between annulus 6 and 47; scales and rows per fold increasing in size and number to at most two rows per annulus, posteriorly. Tongue anteriorly not completely attached to floor of mouth. PMT maximally 22, with notable variation in size, posterior maxillary teeth smaller, extending posteriorly of level of choanae as for PPT series. PPT maximally 20, smaller than PMT, posterior ones smallest. DT maximally 20, approximately same size as PMT; posterior teeth smaller. ST at most four, approximately same size as PPT.
Colour. In preserved and in live specimens body shows different shades of gray or blue. Lateral and ventral surface of body slightly paler than dorsum. Dark colour of dorsum extends laterally and ventrally in primary grooves of most specimens. There are two different patterns of ventral colouration that occur in all populations here studied: (1) a whitish venter with scattered dark blotches of irregular size and shape approximating colour of dorsum; (2) a uniform venter slightly paler than dorsum ( Fig. 9 View FIGURE 9 ). Both patterns also occur in specimens from Suriname ( Nussbaum & Hoogmoed, 1979).
Variation. There are no significant differences between the sexes for any morphometric or meristic character. A similar result was obtained in a study of a population of C. gracilis from Maranhão, Brazil (MAciel et al, in press).
Comparison of meristic and morphometric characters of populations reveals no great variations between populations sampled here ( Table 5 View TABLE 5 ). Variation in primary and secondary annuli is similar among the studied populations, and the total variation in these characters (PA 114–131; SG 23–44) is similar to that for fifteen C. tentaculata from Napo, Ecuador (PA 115–122; SG 23–45) ( Taylor, 1973).
The stepwise discriminant analysis of morphometric characters showed four variables (TL, HL, TMM, NN) that contribute most to the OTUs divergence ( Fig. 3 View FIGURE 3 ; Table 6), and a significant difference among OTUs (Wilks’s Lambda 0.577 p = 0.000). The first discriminant function explains 74.8 % and the first and second together 100% of the total variation. A total of 60% of classifications were correct, 55% Jackknifed ( Table 7). The first axis of the SDFA is better represented by HL and NN, reflecting a weak separation of the OTU MT (Mato Grosso) from PAB (Maraj-Belém-Carajás) and PAGS (Brazilian Guiana) ( Fig. 3 View FIGURE 3 ). The second axis is better represented by TL and TMM, but it does not separate OTUs.
Distribution. Brazil (states of Acre, Amapá, Amazonas, Maranhão, Mato Grosso, and Pará), Colombia, Ecuador, French Guiana, Guyana, Peru, Suriname and Venezuela ( Fig. 10 View FIGURE 10 ).
Remarks. Taylor (1972) removed C. albiventris Daudin, 1803 from the synonymy of C. tentaculata on the basis of the presence of diminutive subdermal scales in C. albiventris and the colour pattern of the belly. Taylor & Peters (1974) provided photos of a specimen of C. albiventris from eastern Ecuador. Frost (1985, 2008) listed C. albiventris as a valid species with a distribution in Suriname and eastern Ecuador. The meristic data for C. albiventris are well within the range known for C. tentaculata , and we observed a large variation in the pattern of the belly, which thus does not separate the two supposed taxa either. Apparently only the presence or absence of subdermal scales seems to differentiate these two species.
Lynch (1999) doubted whether C. guntheri Dunn, 1942 and C. volcani Taylor, 1969 were really different from C. tentaculata . Lynch (1999) provided some localities for C. tentaculata in Chocoan Colombia, which we did not use in our map, because we consider these most likely to be C. isthmica Cope, 1877 (cf. Savage & Wake, 2001; Frost, 2008), and C. tentaculata is considered as having a cis-Andean distribution.
continued next page Acre Mato Grosso PAGS PAB Manaus region continued next page Acre Mato Grosso PAGS PAB Manaus region DT 9– 11 10–20 12–17 10–15 17 (1)
10±4.3 (3) 15.4±3.8 (24) 14.4±1.5 (12) 13.6±6.4 (5)
ST 0–4 (2) 2–4 1–3 3–4
3±1.1 (23) 2.5±1 (4) 3.4±1.6 (5)
TL/BW 27.5–33.1 27.5–46.8 22.2–43.8 30.1–38.8 26.5–30.4
30.6±2 (4) 38.1±4.1 (23) 30.5±4.8 (20) 33.9±2.8 (8) 28.4±1.5 (3) SI 23–28 20–47 6 –29 34–35 28–31 (2)
25.7±2.2 (4) 30±8.5 (7) 17.2±8.4 (8) 34.3±14.8 (3)
Axis 1 Axis 2 TL 0.411 1.029 HL 0.681 0.184 TMM - 0.364 0.646 NN - 0.590 0.043
MT PA1 PA2 % correct Jackknifed % correct MT 18 7 0 72 68 PA 1 8 10 2 50 40 PA2 3 3 7 54 54 Total 29 20 9 60 55
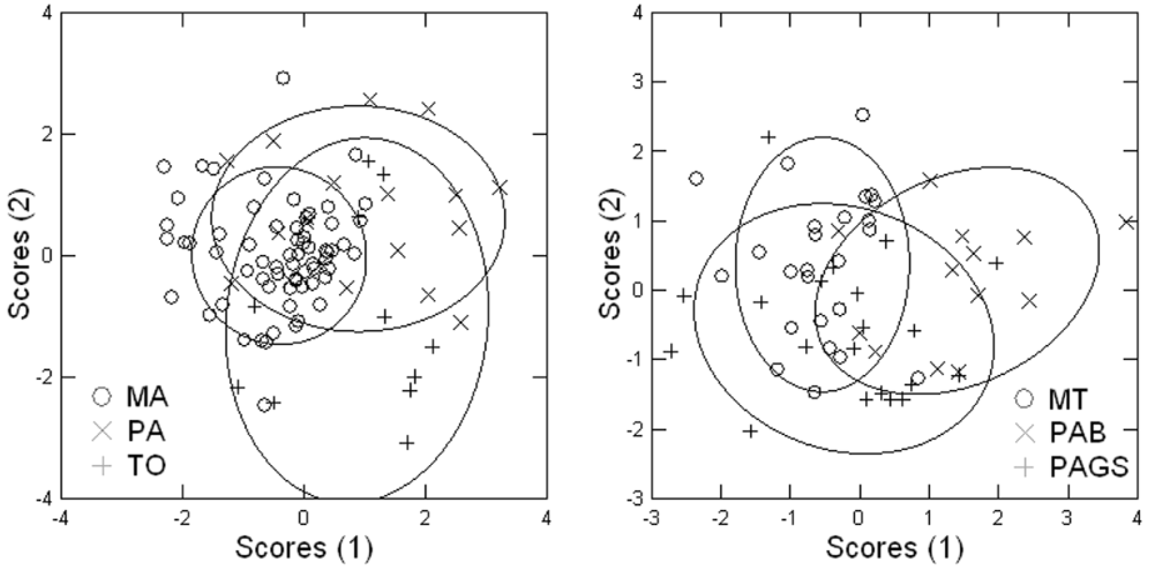


TABLE 5. Morphometric (in mm) and meristic data (range, mean ± standard deviation and sample size in parenthesis) of Caecilia tentaculata. PAGS: data of specimens from state of Pará in the Guiana shield; PAB: data of specimens from state of Pará from the region of Belém and other localities south of the Amazon.
| Acre | Mato Grosso | PAGS | PAB | Manaus region | |
|---|---|---|---|---|---|
| TL | 485–783 677±114.6 (4) | 177.5–500.5 365.6±98.9 (25) | 340–720 513.6±98.9 (20) | 345–566 460.7±65 (15) | 202–820 605.5±238.7 (4) |
| HW | 11.3–17.8 15.1±2.3 (4) | 5.5–9.5 7.7±1 (25) | 8.9–16.1 11.7±2.3 (20) | 7.7–11 9.8±3.4 (13) | 5.7–17.1 12.4±6.8 (3) |
| HL | 15.8–22.4 19±2.3 (4) | 6.5–13 9.9±1.5 (25) | 11–21.9 15.7±2.7 (20) | 12.6–16.9 14±5.6 (13) | 7.9–21.6 16.3±8.8 (3) |
| HH | 8.7–13 10.6±1.5 (4) | 3.7–6.2 5.3±2.4 (7) | 6.1–13.5 9±2.2 (20) | 6.3–9.6 7.4±3 (13) | 3.9–13.7 9.7±5.5 (3) |
| BW | 17.6–25.9 22±3 (4) | 5.5–12.5 9.6±3.1 (23) | 10.5–29.9 17.2±4.6 (20) | 11.6–18.8 13.7±7 (8) | 7.1–25.4 18.7±10.8 (3) |
| BH | 16.2–22.6 19.8±2.4 (4) | 5.1–10.1 7.9±3.2 (21) | 9–26.5 15.3±4.2 (20) | 10.4–15.5 12.4±6.3 (8) | 5.6–23 17.1±10.2 (3) |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Caecilia tentaculata Linnaeus, 1758
| Maciel, Adriano O. & Hoogmoed, Marinus S. 2011 |
Caecilia tentaculata
| Lescure 2000: 292 |
| Frost 1985: 622 |
| Taylor 1968: 555 |
| Dunn 1942: 480 |