
Parabrachyodus hyopotamoides, (LYDEKKER, 1883)
|
publication ID |
https://doi.org/ 10.1093/zoolinnean/zlac111 |
|
DOI |
https://doi.org/10.5281/zenodo.7927134 |
|
persistent identifier |
https://treatment.plazi.org/id/3E5A87FC-FFAB-9D12-4824-F94EFB15B63D |
|
treatment provided by |
Plazi |
|
scientific name |
Parabrachyodus hyopotamoides |
| status |
|
PARABRACHYODUS HYOPOTAMOIDES ( LYDEKKER, 1883)
( FIGS 4–8 View Figure 4 View Figure 5 View Figure 6 View Figure 7 View Figure 8 )
Selected synonymy (exhaustiƲe synonymy list of 100 synonyms in Supporting Information, Appendix S1):
Upper and lower molars of a gigantic species of Hyopotamus ; Lydekker, 1882: 107.
Anthracotherium hyopotamoides Lydekker ; Lydekker, 1883: 152–154, pl. 24, fig. 2, pl. 25, figs 1, 3.
Hyopotamus giganteus Lydekker ; Lydekker, 1883: 160–164, fig. 1, pl. 24, fig. 3, pl. 25, fig. 2.
Brachyodus giganteus (Lydekker) ; Depéret, 1895: 407–408. Brachyodus hyopotamoides (Lydekker) ; Andrews, 1899: 484.
Brachyodus africanus Andrews ; Pilgrim, 1912: 59–62, pl. 22, figs 2–4 (no fig. 1, 5) (in part).
Telmatodon bugtiensis Pilgrim ; Pilgrim, 1912: pl. 24, figs 2, 3b (no pl. 24, fig. 4), pl. 25, fig. 6 (in part).
Brachyodus pilgrimi Forster-Cooper ; Forster-Cooper, 1913: 516–517, fig. 3 (viewed).
Parabrachyodus obtusus (Forster-Cooper) ; Forster-Cooper, 1915: 404–406, figs 1, 2 (viewed).
Brachyodus gandoiensis Forster-Cooper ; Forster-Cooper, 1924: 27–28, pl. 1, figs 3–6 (viewed).
Brachyodus platydens Forster-Cooper ; Forster-Cooper, 1924: 29–31, pl. 2, figs 2, 3 (viewed).
Brachyodus orientalis Forster-Cooper ; Forster-Cooper, 1924: 32, pl. 3, figs 3, 4 (viewed).
Brachyodus indicus Forster-Cooper ; Forster-Cooper, 1924: 32, pl. 4, fig. 1 (viewed).
Gonotelma major Forster-Cooper ; Forster-Cooper, 1924: 49–50, pl. 5, fig. 1 (viewed).
Parabrachyodus Forster-Cooper ; Viret, 1961: 948–949.
Brachyodus manchharensis Prasad ; Prasad, 1964: 9–12. Anthracotherium bugtiense Pilgrim ; Pickford, 1987: 309–311 (in part).
Parabrachyodus hyopotamoides (Lydekker) ; Pickford, 1987: 316–319 (in part).
Telmatodon Pilgrim ; Pickford, 1987: 320–323 (in part).
Parabrachyodus hyopotamoides (Lydekker) ;Welcomme et al., 1997: 533, 535.
Parabrachyodus hyopotamoides (Lydekker) ; Lindsay et al., 2005: 16, table 1 (in part).
Parabrachyodus hyopotamoides (Lydekker) ; Bhandari et al., 2010: 76, fig. 7A.
Parabrachyodus hyopotamoides (Lydekker) ; Antoine et al., 2013: tabs 1, 2, fig. 4 (in part).
Holotype: Right maxilla with mesiolabially damaged M3/ and roots of M2/, GSI B426, figured by Lydekker (1883: pl. 24, fig. 2) and housed in the collections of the Indian Museum, Calcutta, India.
Type locality: Exact locality unknown, ‘ lower Manchhars of the Bhugti hills, north of Sind’ ( Lydekker, 1882: 107), Indus Basin, Pakistan.
Localities: Pakistan: Sulaiman Province: Dera Bugti, Lundo Chur, Kumbi, Samane Nala (levels 4 and 5), Tobah (level Q = 4), all in the Bugti Hills, Zinda Pir area (Z114, Z154, Lindsay et al., 2005; Safed Nala). India: Fategad, central Kutch.
Horizon: Chitarwata Formation (level 4, upper member, Métais et al., 2009), Vihowa Formation (level 5, lower member; Antoine et al., 2013), Khari Nadi Formation ( Patnaik & Prasad, 2016), seemingly Manchar Formation (lower member; Lydekker, 1882; Raza et al., 1984).
Age: Early Miocene, from c. 22 to 19–18 Mya ( Lindsay et al., 2005; Métais et al., 2009: Roddaz et al., 2011: fig. 4; Antoine et al., 2013).
Neae material: The whole new fossil material (22 specimens) used here is housed in the palaeontological collections of the Université de Montpellier ( UM). Material from Samane Nala, level 4 ( SAM 4 ) of the upper Chitarwata Formation, Dera Bugti ( DB) area, northern flank (19 specimens): right I2/ or I3/ (UM-SAM4-024); right P1/ (UM-SAM4-025); skull with left P3/-M3/ tooth row and P1/-P2/ alveoli (UM-SAM4-001); skull fragment with left P4/- M1/ (UM-SAM4-002); right maxilla with M2/-M3/ (UM-SAM4-003); crown of right M3/ (UM-SAM4-004); worn right M 3/ (UM-SAM 4 - 006); right M 3 / (UM-SAM4-007); worn left M3/ (UM-SAM4-008); lingual fragment of left M3/ (UM-SAM4-009); distal fragment of left M3/ (UM-SAM4-011); crown of left I/1 (UM-SAM4-026); left P/1 (UM-SAM4-027); fragment of left mandibular symphysis with P/2-P/3 alveoli ( UM DB SAM4-014); right M/2 (UM-SAM4-015); worn left M/2 (UM-SAM4-016); fragment of right mandible with M/3 and talonid of M/2 (UM-SAM4-013); fragment of left M/3 without hypoconulid (UM-SAM4-017); talonid and hypoconulid of left M/3 (UM-SAM4-020)
.
Material from Samane Nala, level 5 ( SAM 5) of the Vihowa Formation, Dera Bugti area, northern flank of the Zin anticline: left M3/ (UM-SAM5-001).
Material from Tobah, level Q of the Chitarwata Formation, Dera Bugti area, southern flank of the Zin anticline: right mandible with P/4-M/3 (UM-TOB-001).
Material from the Zinda Pir Dome ( Pakistan), Safed Nala: right M/3 (UM-SFN-001).
Other material: Material from the historical field expeditions in the Bugti Hills ( Pakistan): ‘ Dera Bugti’ , ‘ Chur Lando’ , ‘ Kumbi’ and ‘ Bugti’ (i.e. for ‘ Bugti’ , on the flanks of the Zin anticline; see Pickford, 1987: fig. 2), level unknown, Dera Bugti area (82 specimens) :
- Material collected by the Geological Survey of India ( GSI) , housed in the Indian Museum of Calcutta (21 specimens) . The revision of the following specimens was in part based on casts housed in the NHM, and on published illustrations. The attributions of some GSI specimens to Par. hyopotamoides not mentioned here but proposed by Pickford (1987) could not be verified in absence of illustrations or available casts. Loaeer dentition: right P/1-M/2 ( GSI B479 / M11076) ; left P/3-P/4 ( GSI B485 ) ; left P/3-M/3 ( GSI B478 ) ; right P/3-M/3 ( GSI B420 ) ; P/3-M/3 ( GSI B458 ) ; left M/1- M/3 ( GSI B521 / M11062) ; right M/2 ( GSI B429 ) ; left M/2-M/3 ( GSI B459 ) ; right M/3 ( GSI B428 / M1541) ; half right M/3 ( GSI B430 ) ; broken right M/3 ( GSI B431 ) ; M/3 ( GSI B460 ) . Upper dentition: left P2/ ( GSI B518 / M11065) ; left P3/-P4/ ( GSI B456 / M10598) ; left P4/ ( GSI B457 ) ; palate with right M1/-M3/ and left M3/ ( GSI B462 ; Pilgrim, 1912, pl. 22, fig. 2) ; skull with left M1/-M2/ and right M1/-M3/ ( GSI B461 / M11060; Pilgrim, 1912: pls 19, 20) ; skull with left M1/-M2/ and right M3/ ( GSI B454 / M11064) ; right M3/ ( GSI B426 ) ; left M3/ ( GSI B427 , GSI B433) .
- Material collected from several expeditions in the Bugti Hills, studied by Forster-Cooper (1924) and housed in the Bugti collection of the NHM (61 specimens). Loaeer dentition: mandible with left I/2, left P/4-M/3 and right P/4-M/3 (M12724); right P/1-M/1 (M12736); left P/2-M/1 (M12721); right P/3- M/3 (M12723); left P/3-M/3 (M12731); left P/4-M/3 (M12720); right M/1-M/2 (M12732); left M/1-M/3 (M12733, M12734); right M/2-M/3 (M12727); left M/2-M/3 (M12729, M12735); right M/3 (M12726); left M/3 (M12030, M12725, M12730); broken left M/3 (M12722); worn right M/3 (M12719). Upper dentition: left P1/-M3/ (M12034); left dP3/-M1/ (M12820); right P3/-M2/ (M96529); skull with left P3/-M2/ and right M3/ (M12821); palate with left dP4/-M1/ and right dP4/ (M43964); left P4/- M1/ (M43962); right P4/-M2/ (M12713); left P4/- M2/ (M12827); right P4/-M3/ (M11061, M12712, M12717, M12822, M43953); left P4/-M3/ (M12031, M12714, M12715, M12826); right M3/ with worn right P4/-M1/ (M43963); right M1/-M2/ (M43960, M43960); right M1/-M3/ (M12818); left M1/-M3/ (M12032, M12035, M12036, M12718); right M2-3/ (M12039); right M2/-M3/ (M12716, M12817, M12823, M12824), left M2/-M3/ (M12711, M12819, M12825, M43950, M43951); right M3/ (M12033, M43952, M43955, M43958); left M3/ (M43956, M43957, M43959); skull roof (M43949).
Material from the Zinda Pir Dome ( Pakistan), collected by the GSI, studied by Lindsay et al. (2005) and housed in the Peabody Museum of Harvard University (2 specimens); locality Z154, right M3/ (z2077); locality Z114, left P4/ (z258) .
Material from Fategad, central Kutch ( India), Khari Nadi Formation (latest Early Miocene), collected by the GSI, studied by Prasad (1964, 1967) and Bhandari et al. (2010) and housed in the Indian Museum of Calcutta: palate with left M2/-M3/ and fragment of right M2/ ( GSI 18097).
Emended diagnosis [modified from Pickford (1987)]: Medium-sized to large bothriodontine with a unique combination of characters among anthracotheres: main palatal foramen open in front of M1/, I/2 smaller than I/3, large alveoli for lower and upper canines, which are separated from premolars by short diastema, no diastema between lower and upper jugal teeth, P/1 biradiculate, preprotocristid of lower premolars with an accessory cusp, P/4 postprotocristid centred with respect to the mesiodistal axis and not fused with the endoprotocristid, P1/-P3/ with two mesial ridges (a preparacrista and an ectoparacrista) and a postparacrista with two accessory cusps, P3/ with distolingual cingular style, P4/ with an endoprotocrista and a large distostyle, pentacuspid upper molars with a continuous transverse valley and a low crown, pronounced flare of the lingual flank of the lingual cuspids on upper molars, developed loop-like labial styles on upper molars, parastyle formed by preparacrista, quadricrescentic protocone with an ectoprotocrista connected to a protostyle on the mesial cingulum, postectoprotocrista directed towards the lingual margin of upper molars along the transverse valley, deep ectoprotofossa and postectoprotofossa, lack of premeta-preprotocristid connection on lower molars, postentocristid often present, prehypocristid of M/3 with two mesial arms, pinched hypoconulid aligned with lingual cuspids, wrinkled and thick molar enamel with a large zone of radial outer enamel on the bizonal Schmelzmuster, dP3/ with bicuspid anterior lobe and entostyle.
Differential diagnosis: Differs from other bothriodontines by its additional styles and crests on lower and upper jugal teeth. Parabrachyodus differs from Elomeryx in the lack of junction between the upper molar protocristae and premetacristule. Among bothriodontines from the Bugti Hills: differs from Telmatodon and Gonotelma mainly in the presence of a paraconule on upper molars. It differs also from T. bugtiensis in a less prominent ectostylid (when present) on its M/3. Contrary to T. orientalis , the postectoprotocrista joins the lingual margin on its upper molars, and there is an endoprotocrista on P4/. Parabrachyodus differs from Gonotelma shahbazi in the presence of a postmetafossule, the less steep-sided lingual flank slope of the lingual cusps on upper molars, the alignment of the M/3 hypoconulid with lingual cuspids, and its much larger teeth. The paraconule is well distinct from the protocone compared to SiƲameryx palaeindicus , and the postparacristule reaches the transverse valley instead of the base of the paracone. Differs from advanced Merycopotamini , such as SiƲameryx and Hemimeryx blanfordi , in the configuration of the prehypocristid, which does not reach the lingual margin of lower molars, and the more robust mandible with a more obtuse angle between the symphysis and the jugal tooth row.
ComparatiƲe descriptions
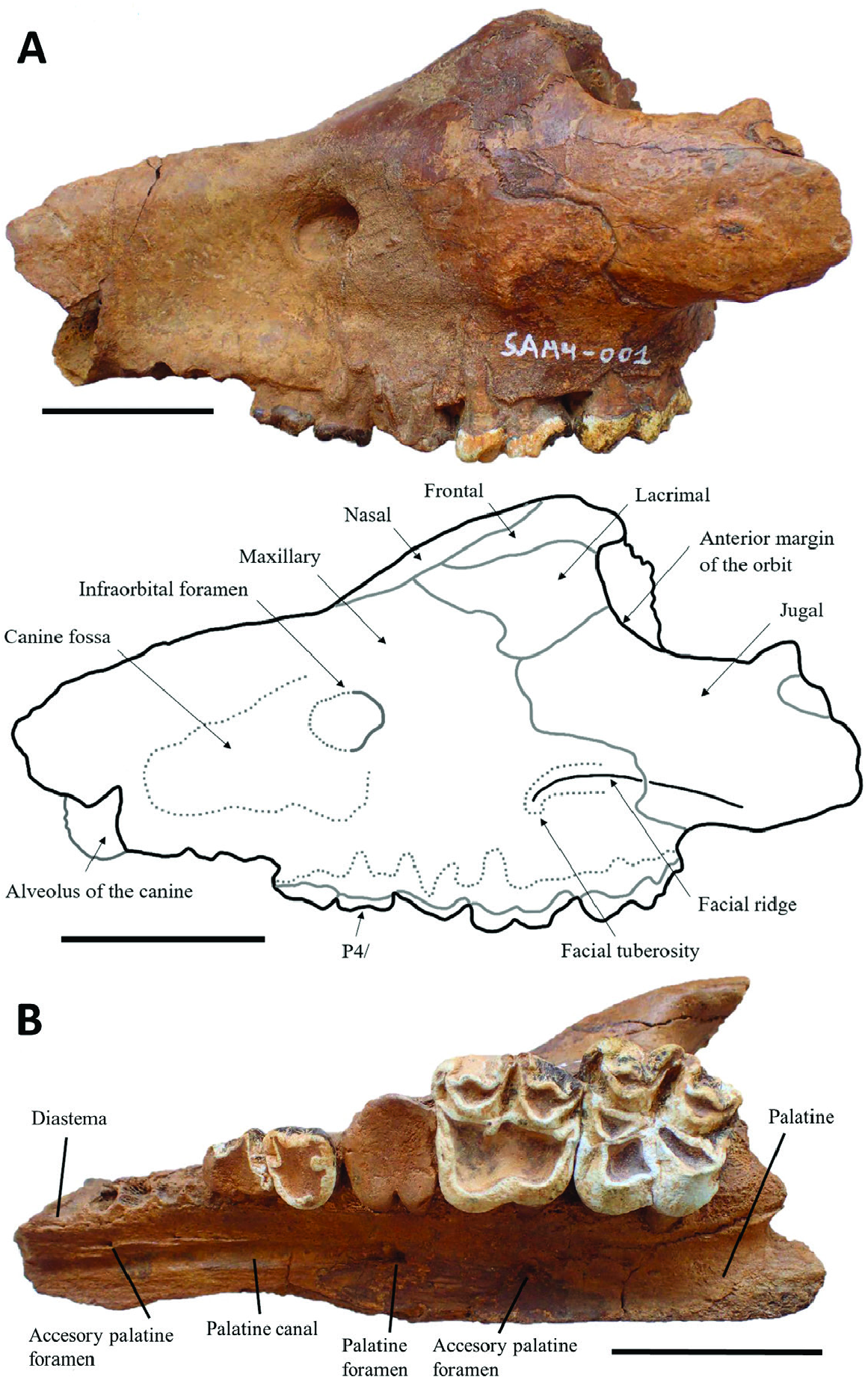
Cranium: In lateral view ( Fig. 4A View Figure 4 ), the skull is shallow and elongated as in Brachyodus onoideus ( Dineur, 1981; Orliac et al., 2013). The jugal appears relatively deep compared to that of other bothriodontines, such as Elomeryx ( Lihoreau et al., 2009) . The facial ridge runs ventrally along the jugal until the level of the infraorbital foramen. It intrudes on the maxilla where the maxillojugal suture delimits a brief concavity of the jugal, then bifurcates ventrally to reach the facial tuberosity above the M2/. The combination of these cranial characters is found on all available maxillae of Parabrachyodus (UM-SAM4-002, M12033, M12717 and M12718), but also on the sole cranium of SiƲameryx , which preserves this area ( Miller et al., 2014). A strong concavity of the face is anterior to the facial tuberosity, so that the snout is narrow at the level of the canine fossa ( Fig. 4A, M View Figure 4 12821), unlike the condition seen in Brachyodus aequatorialis ( MacInnes, 1951: pl. 1) and Br. onoideus ( Dineur, 1981) . The infraorbital foramen is located above the P4/ on UM-SAM4-001 and above the M1/ on UM-SAM4-002. There is also a reduced distance between this foramen and the anterior margin of the orbit in the latter with respect to UM-SAM4-001 (64 mm vs. 74.8 mm). The lacrimal contributes to the anterior margin of the orbit and joins a narrow nasal by a short process separating the frontal from the maxillary on UM-SAM4-001. This lacrimonasal contact is absent on other skulls reported to Par. hyopotamoides (M11064, M43949) and SiƲameryx ( Rowan et al., 2015) . The cranium of SAM 4 is thus an exception. Such a variation of the lacrimonasal configuration is unknown among other anthracotheres (i.e. Hippopotamoidea, excluding Hippopotamidae ) at the species level. This morphology was highlighted as a derived character in Hippopotamus , more generally regarded as a fixed and constant generic character in the whole Hippopotamidae ( Boisserie, 2005) . The orbits are slightly more dorsally elevated (M43949) than in SiƲameryx , and are similar to the condition seen in Brachyodus onoideus ( Orliac et al., 2013) , but are not as much elevated as those in Merycopotamus dissimilis Falconer & Cautley, 1847 (Lihoreau et al., 2007) or Libycosaurus bahri Lihoreau et al., 2014 ( Lihoreau et al., 2014). There is neither lacrimal foramen nor lacrimal canal, except on M43949. In ventral view ( Fig. 4B View Figure 4 ), the palatine advances between the maxillaries to the posterior part of the M1/, like in all anthracotheres.
The main palatine foramen is situated at the level of the M1/ on the cranium from SAM 4, whereas in Elomeryx it is located between the level of the P2/ to P4/ ( Lihoreau et al., 2009), the P2/ in Brachyodus onoideus ( Dineur, 1981: 25) and Merycopotamus (Lihoreau et al., 2007) , and is between the mesialmost upper premolar (PX/) and the canine in Libycosaurus ( Lihoreau et al., 2014) . Anteriorly, a wide palatal canal forms a groove over the rest of the dental row and has an accessory palatal foramen at the level of the P1/. A second accessory palatal foramen is located on the maxillopalatine suture in front of the M2/ ( Fig. 4B View Figure 4 ). This configuration of the palatal foramina is also exhibited by the M12714 ( Forster-Cooper, 1924: pl. 2, fig. 1) and M12826 specimens. Pilgrim (1912: 53, 58) noted a palatal foramen near the P3/ on some GSI specimens, but it is not appreciable on the available casts and illustrations of the referred skulls. Only the M12821 skull with Parabrachyodus -like molars clearly displays this character state. The choanae open behind M3/ at the level of the palatine process and the maxillary tuberosity (UM-SAM4-001, -003) as in all bothriodontines.
In dorsal view [only on material published by Pilgrim (1912)], the angle between the sagittal crest and the frontal lines is more obtuse on the skulls M11060 and M43949 (> 80°) than on M11064 (60°). Furthermore, the sagittal crests of M43949 and M11064 are longer than on M11060. In the absence of more material likely to quantitatively address this variation, it is not possible to conclude whether or not there is a marked sexual dimorphism like that observed in Merycopotamus and Libycosaurus (Lihoreau et al., 2007, 2014). The median occipital ridge (M11064) is surrounded by two nuchal fossae also found in Merycopotamus (Lihoreau et al., 2007) .
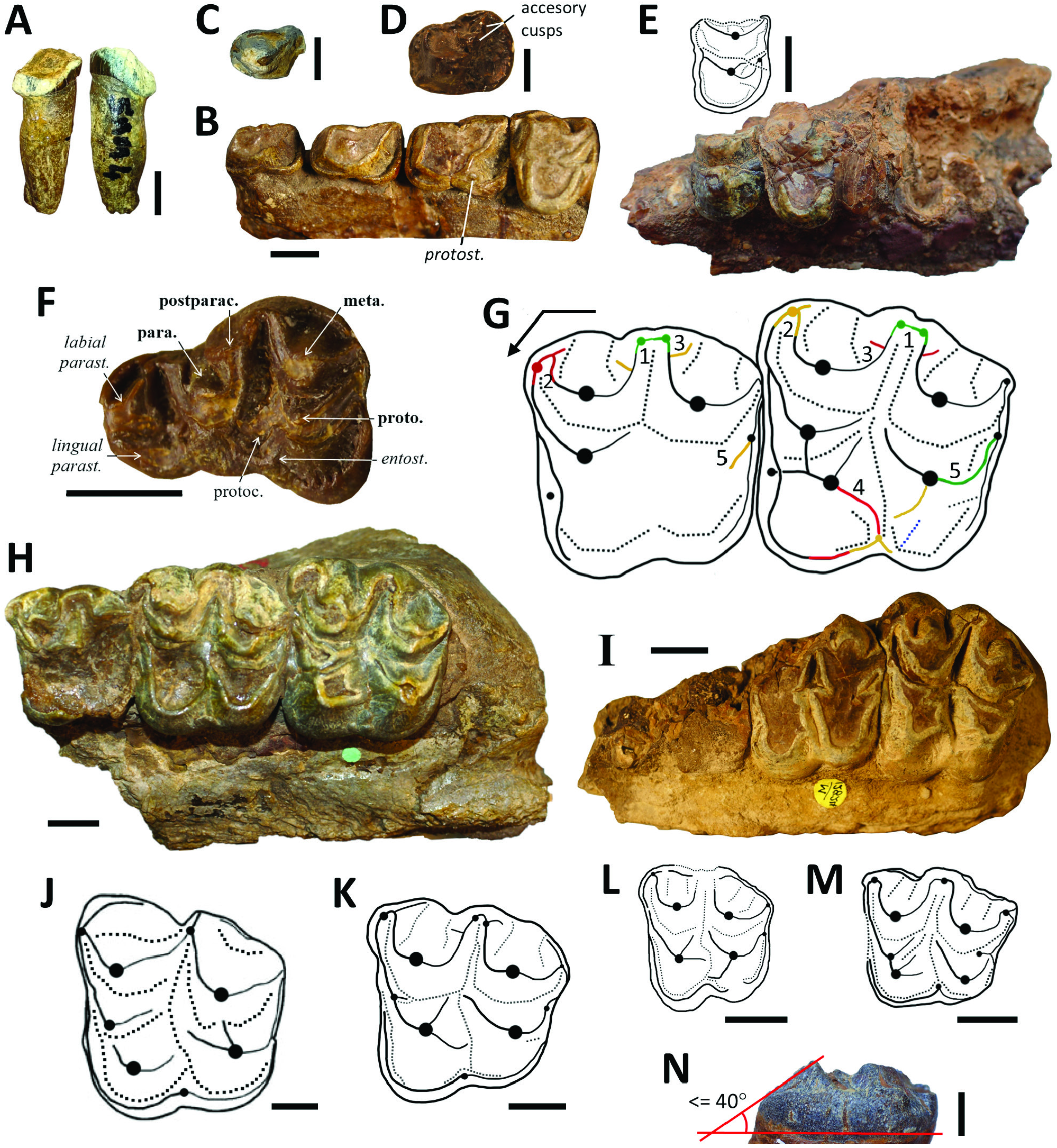
Rostral dentition: Of the premaxillary region and rostral teeth of Par. hyopotamoides , only an isolated right I2/ or I3/ is known, measuring 38 mm from root to crown (UM-SAM4-024; Fig. 5A View Figure 5 ). Its root is relatively short and smaller in diameter than the crown. The crown has a spatulate shape, a mesial and a distal pinched crest, and wrinkled enamel that is a characteristic feature of Par. hyopotamoides . The distal crest is higher than the mesial crest. A strong lingual cingulum is adjacent to the mesial border. On the skull UM-SAM4-001 ( Fig. 4A View Figure 4 ) the alveolus for the canine has a large diameter (18.3 mm) and it is transversely compressed. The canine is separated from the P1/ by a relatively short diastema only visible on the skull of SAM 4 (19.3 mm; Fig. 4B View Figure 4 ), while that of Brachyodus is long (e.g. Pickford, 1991).
Upper premolars: The upper jugal teeth are in a contiguous row (UM-SAM4-001 and M12034; Figs 4B View Figure 4 , 5B View Figure 5 ), which contrasts with Eocene and Oligocene bothriodontines displaying diastema between their premolars, such as Bothriodon and Bothriogenys ( Dineur, 1981: 159) or Elomeryx Geais (1934) . P1/ to P3/ are biradiculated and exhibit two mesial ridges: a preparacrista and an ectoparacrista. The occlusal pattern of P1/ is known from an isolated specimen from SAM 4 (UM-SAM4-025; Fig. 5C View Figure 5 ), which displays weak accessory cusps on the postparacrista and looks more like P1/ than P2/ of M 12034 in shape and size. The P3/ is larger than the P2/, with a more developed distolingual basin and a squarer shape (M12034; Fig. 5B View Figure 5 ). In addition, the paracone of the P3/ is flanked by a small distolingual cingulate style (protostyle, Fig. 5C View Figure 5 ), as in Anthracotherium ( Scherler et al., 2018: fig. 3). The P3/ has two accessory cusps on the postparacrista (GSI B456; Fig. 5D View Figure 5 ) and the two mesial ridges reminiscent of Merycopotamini , such as SiƲameryx palaeindicus ( Holroyd et al., 2010: fig. 43.4C), are present.
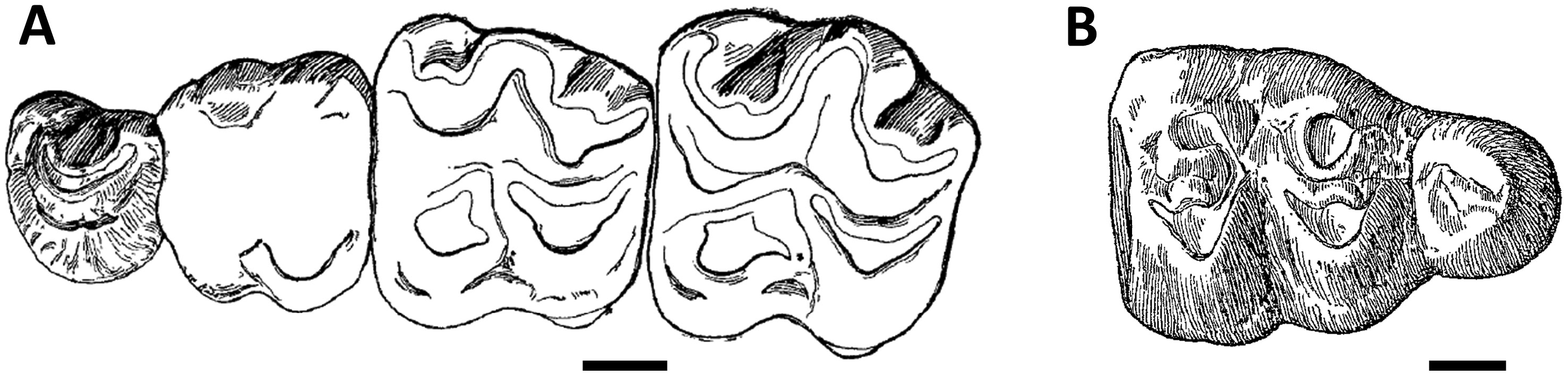
The P4/ is always bicuspidate, except for that of the maxilla, which Forster-Cooper (1915) erroneously interpreted as the ‘holotype’ of the genus Parabrachyodus (M12031; Fig. 1A View Figure 1 ), and for M12826 and M12827, which both lack the protocone (20%, 3/15 P4/ with a well-preserved occlusal surface). In the classic pattern of P4/ (UM-SAM4-002; Fig. 5E View Figure 5 ), three ridges arise from the protocone with a rounded contour and less selenodont than forms like S. palaeindicus . The pre- and post-paracristae are divergent in the same way as the latter, in contrast to Merycopotamus where they tend to parallelize (Lihoreau et al., 2007). A strong cingulum surrounds the entire tooth and opens distally into a large distostyle (M12712). The latter is in contact with the postprotocrista like in Telmatodon ( Forster-Cooper, 1924) , whereas it appears free most of the time in bothriodontines such as Bothriogenys orientalis Ducrocq, 1997 ( Ducrocq, 1997) . In Par. hyopotamoides , an additional crest is similar in position to the postprotocrista known in Myaingtherium Tsubamoto et al., 2011 ( Tsubamoto et al., 2011) and in some anthracotheriines ( Scherler et al., 2018). We interpret it as an additional ‘endoprotocrista’, regarding the more lingual position of the postprotocrista in Bothriodontinae (see Discussion). One-third of the P4/ specimens have an endoprotocrista with a vacant distal end as on the P4/ of SAM 4 with a preserved occlusal pattern ( Fig. 5E View Figure 5 ). The other two-thirds show an endoprotocrista that joins the base of the paracone (GSI B456, M11061 or M12822). It is worth noting that two P4/s attributed to S. palaeindicus from SAM 4 similarly show a variation in the orientation of the postprotocrista, directed towards the distostyle or the base of the paracone. This was also observed on some P4/ of S. palaeindicus studied by Forster-Cooper (1924: 33).
Upper deciduous dentition: The only known and unpublished dP3/ (M12820; Fig. 5F View Figure 5 ) bears four main cusps and three accessory cusps as Merycopotamus ( Gomes Rodrigues et al., 2020) , but it is larger. On the same specimen, the dP4/ is a smaller copy of the adjacent M1/, whose protocone possesses the characteristic features of Parabrachyodus (see also M43964). In addition to the four main cusps, the dP3/ is characterized by an entostyle and accessory cusps (lingual parastyle and postparaconule), whose appearance is delayed during the development of cusps ( Gomes Rodrigues et al., 2020: table 2). By contrast, only the postparaconule is present in upper deciduous premolars of Elomeryx armatus ( Scott, 1940: fig. 122).
Upper molars: The teeth from SAM 4 have a morphological disparity (illustrated in Fig. 5G View Figure 5 ) also encountered in other sites of the Bugti Hills, as described hereafter. The interindividual variation does not allow us to subdivide the material into coherent taxonomic units in a small series from a single locality (here SAM 4). Pilgrim (1912: 49) already underlined the difficulty of distinguishing, based on stable characters, the two species Brachyodus giganteus ( Lydekker, 1883) and Br. hyopotamoides then discriminated by himself and Lydekker (1883). Thus, the wide, looped mesostyle surrounding a deep, transverse valley without ectocristyle is divided into two slender apices connected to each other (issued from the postparacrista and premetacrista) in less than half of the specimens (43%, 13/30; Fig. 5G View Figure 5 ). The presence of a mesostyle with two apices is also common in Elomeryx armatus and E. borbonicus Geais, 1934 ( Scott, 1940: fig. 122). Each of these character states (mesostyle composed of one or two apices) here certainly depend on the degree of tooth wear.
For the whole upper teeth sample, the normality assumption is rejected only for the labiolingual length of the mesial lobe of M3/ (Shapiro-Wilk test with p <0.05, Table 2 View Table 2 ). On some M3/s (UM-SAM5-001 and M43951), this part of the tooth is excessively extended lingually, and as such it is reminscent of the more triangular shape of Telmatodon ( Fig. 5K View Figure 5 ) and Gonotelma ( Fig. 5L View Figure 5 ). In addition, the parastyle resulting from the preparacrista and not from the labial cingulum, unlike Brachyodus ( Fig. 5J View Figure 5 ; Lihoreau & Ducrocq, 2007), is larger than the mesostyle on one-third of the M3/s (5/15, e.g. M12034 and M12035). They are otherwise equal ( Fig. 5G View Figure 5 ), which gives a square shape to the teeth. Since the origins of the variability in labiolingual width are diverse, it is not possible to discriminate between individuals solely on such a difference in shape (see Fig. 5H View Figure 5 vs. Fig. 5I View Figure 5 ). The metastyle is reduced compared to the other two labial styles. The development of the cingula is not the most variable characters of Par. hyopotamoides ( Fig. 5G View Figure 5 ; contra Viret, 1961). In fact, the cingulum surrounding the protocone is only reduced in one specimen of SAM 4 (UM-SAM4-001; Fig. 4B View Figure 4 ) and we found a similar polymorphism in the entire sample for M3/ (77%, 17/22). When a labial wing-like cingulum is present (75%, 24/32 of M/2 and M/3; UM-SAM4-001 and -007; Figs 4B View Figure 4 , 5G View Figure 5 ), it joins the postparacrista and the premetacrista, and thus draws the loop of the mesostyle.
The paraconule is separated from the protocone by a small, transverse fossa, which contrasts with the advanced fusion of these cusps seen in SiƲameryx ( Fig. 5M View Figure 5 ; Grossman et al., 2019). In addition, the postparacristule joins the transverse valley in the same way as in Elomeryx , whereas it connects to the base of the paracone in S. palaeindicus ( Ducrocq & Lihoreau, 2006) . A protostyle adorns the mesial cingulum as observed in Anthracotheriinae ( Scherler et al., 2018) , Geniokeryx Ducrocq, 2020 and Epirigenys Lihoreau et al., 2015 . As in the latter, an accessory ridge could be occasionally noted in the distal transverse fossa (M12715 and M43959). The retention of both the protostyle and this crest underscores the primitiveness of Par. hyopotamoides compared to other Miocene bothriodontines. The species is unique in that a crest we identified as an ectoprotocrista constantly connects the protocone to the protostyle ( Fig. 5G View Figure 5 ). The ectoprotocrista is clearly separated from the preprotocrista at the apex of the protocone on the less worn specimens (M11061 and M43961). In addition, almost all M3/ have a postectoprotocrista running along the transverse valley to the lingual edge (88%, 22/25), where it meets or not an entostyle (68%, 17/25 M3/; Fig. 5G View Figure 5 ). Such orientation of the postectoprotocrista is half as common on M2/ (40%, 4/10). This character is otherwise found on the single known M3/ of Telmatodon bugtiensis ( Pickford, 1987) . The prominence of this crest always tends to deviate the transverse valley distally, which is observed to a lesser extent in Gonotelma ( Fig. 5L View Figure 5 ), SiƲameryx ( Fig. 5L View Figure 5 ) and even Brachyodus ( Fig. 5J View Figure 5 ), whilst it is almost absent in Hemimeryx and most Merycopotamini lacking this distal crest ( Lihoreau et al., 2016: fig. 2D, E). The ectoprotofossa and postectoprotofossa lingual to the ectoprotocrista and postectoprotocrista, respectively, protrude those additional cristae ( Fig. 5G–I View Figure 5 ), thereby implying a multiplication of the number of wear surfaces. The arrangement of the cristae and fossae on the conical protocone is hence different from the selenodont pattern exhibited by other bothriodontines. The distinction of this original pattern of the protocone is enough to recognize Par. hyopotamoides , even for worn or fragmented M3/ from SAM 4 (UM-SAM4-006 and -009). It should be noted that specimens attributed to SiƲameryx palaeindicus may occasionally exhibit some of the peculiarities observed on the protocone of Par. hyopotamoides ( Forster-Cooper, 1924: pl. 4, fig. 7; Lydekker, 1883: pl. 23, fig. 6).
We dis tingu is hed two mo rphs fo r the postmetacristule labial to the postmetafossule: one with a double curvature before a large distostyle (52%, 17/33 upper molars; UM-SAM4-001 and M12036; Fig. 4B View Figure 4 ) and another with a single curvature but less common here (UM-SAM4-011; Fig. 5G View Figure 5 on the M2/). Also, two molars along the same tooth row may vary in types of curvature for their postmetacristule (M12718; Fig. 5H View Figure 5 ), and this may be related to the differential wear of M2/ and M3/. The ectometacristule is either weak (UM-SAM4-009) or protruding (UM-SAM4-011) on M2/ and M3/, almost twice as present on M3/ (73%, 19/26) than on M2/ (40%, 4/10), and this character tends to disappear in other Miocene bothriodontines as well. Moreover, M1/ do not develop an ectometacristule, unlike BakaloƲia and Elomeryx from the Eocene– Oligocene ( Hellmund, 1991; Böhme et al., 2013). The specimens UM-SAM4-007 and UM-SAM4-011 show that a secondary ectometafossule, lingual to the ectometacristule, is sometimes present on M3/ (19%, 5/26; Fig. 5G View Figure 5 ). The M3/ UM-SAM4-004 – and most of the specimens to a lesser extent – is covered with thick wrinkled enamel on the lingual tubercles. This upper molar also shows the shallow aspect of the crown, which is characteristic of Par. hyopotamoides ( Fig. 5N View Figure 5 ; Bhandari et al., 2010). The two lingual roots of the M3/ from SAM 5 (UM-SAM5-001) are well-preserved and distinct, whereas the labial roots are fused. The angle between the lingual flank of the metaconule and the collet of the M3/ ( Fig. 5N View Figure 5 ) is on average closer to that observed on teeth of Telmatodon and Elomeryx (between 40 and 45°), whereas it is more obtuse (between 50 and 60°) in Brachyodus , Gonotelma and all Merycopotamini (Supporting Information, Table S2 View Table 2 ).
The M1/ are significantly smaller than M2/, and the M2/ are significantly smaller than M3/, even if the overlap between M1/-M2/ is lesser than the overlap between M2/-M3/ ( Table 2 View Table 2 ). The dimensions of the jugal teeth from SAM 4 (Supporting Information, Table S1) are included in the range of variation of all the specimens we attributed to Par. hyopotamoides from the collections of the Bugti Hills. In addition, no biometric discrimination has been retrieved, either considering the deposit of origin (i.e. Dera Bugti, Lando Chur, Samane Nala and Bugti Hills) or the ancient attribution (the museum species name of the specimens) with mesiodistal or labiolingual length 1 (Kruskal-Wallis test with p <0.05, Table 3 View Table 3 ).
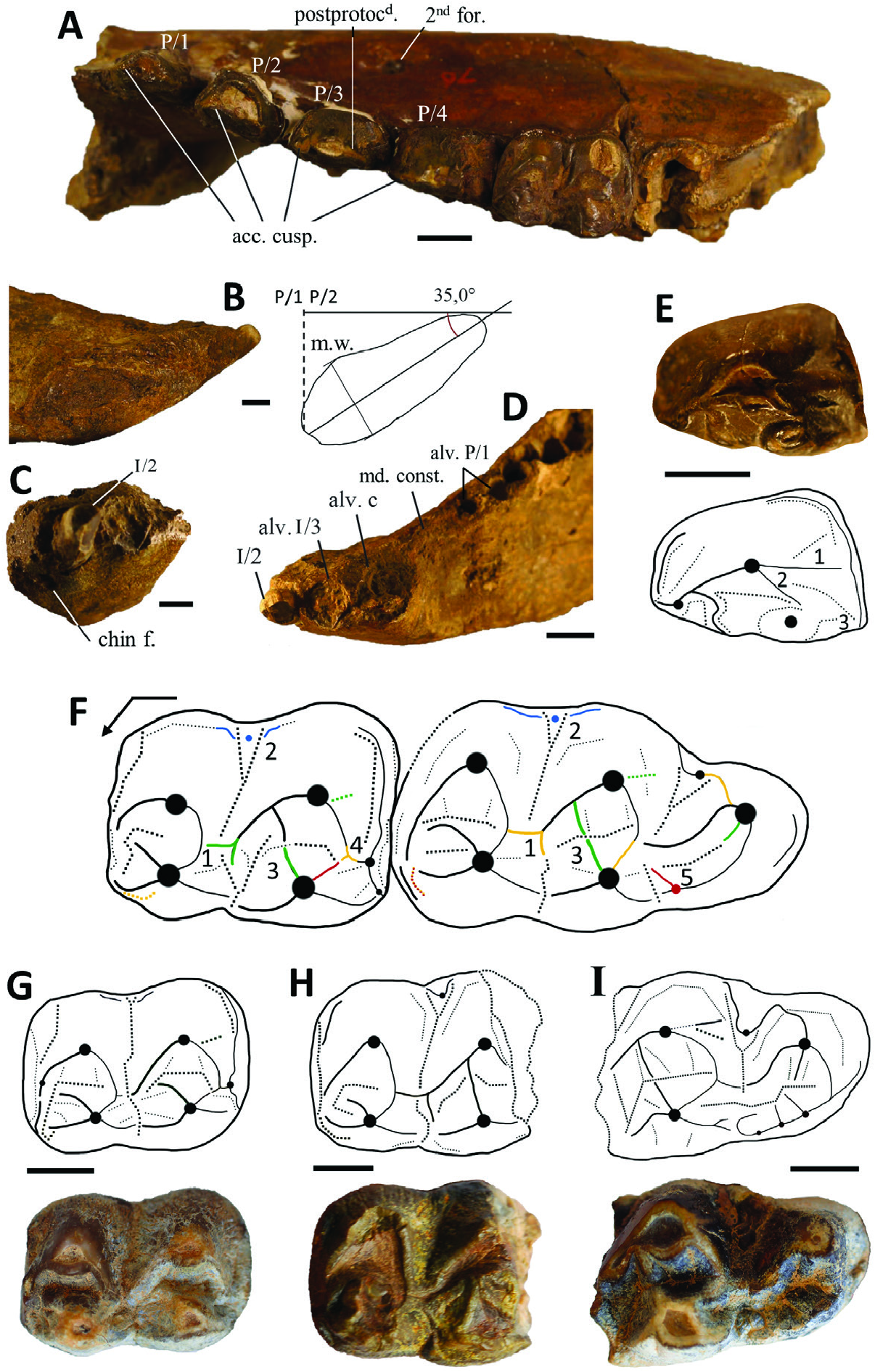
Mandible: The external flank of the mandible has two foramina: one below the P/1-P/2 contact, as observed on the symphysis UM-SAM4-014, and another below the P/3-P/4 contact (M12736; Fig. 6A View Figure 6 ). The mandibular height under the P/2-P/3 is greater on UM-SAM-014 (57.7 mm) than on UM-TOB-001 (47.3 mm), hence it is variable. So is the mandibular height below M/3, which is nonetheless normally distributed here (Shapiro-Wilk test with p <0.05, Table 4). The mandible corpus is wider at the M/3 than mesially (e.g. UM-TOB-001; Fig. 7A View Figure 7 ), and it is also much higher under the M/3 (e.g. 88.9 mm on UM-SFN-001; Fig. 7C View Figure 7 ). The mandibular symphysis ends up between P/1 and P/ 2 in all the specimens referable to Par. hyopotamoides (e.g. Fig. 6B View Figure 6 ). In sagittal section, its dorsal margin is convex, at least distally, on UM-SAM4-014 and M12724 ( Fig. 6B View Figure 6 ; Supporting Information, Appendix S5B) and its ventral margin (M11076, M12724 and M12731) is always straight (convex in Brachyodus ). The greatest distal height of the symphysis (31.6 mm) makes it more robust than that of Hemimeryx ( Lihoreau, 2003: fig.
I.25), a bothriodontine with similar dental proportions ( Lihoreau et al., 2016, table 1). The angle between the anteroposterior axis of the symphysis and the jugal tooth row is 35.0° (M12724, Fig. 6B View Figure 6 ), so it is much more obtuse than in Elomeryx , Hemimeryx or SiƲameryx ( Lihoreau, 2003: fig. I.16). All these characters bring the symphysis of Par. hyopotamoides closer to that of the small Libyan anthracothere Afromeryx zelteni Pickford, 1991 ( Pickford, 1991: pl. 6), except for its robustness and the absence of a genial spine. There is a marked mandibular constriction between c and P/1 without the intermediate protrusion characteristic of Hemimeryx and SiƲameryx ( Fig. 6D View Figure 6 ). Neither the coronoid process nor the angular process was preserved. The gonial region is slightly projected ventrally below the horizontal branch of the mandibles M12723, M12726 and M12732 ( Forster-Cooper, 1924: figs 28, 40), and thus resembled that of Brachyodus ( Dineur, 1981: pl. 9) more than that of Libycosaurus ( Lihoreau et al., 2014) .
Loaeer incisors and canines: The arrangement of the lower incisors is known from the M12724 mandible ( Forster-Cooper, 1924: fig.29), where a socket and a broken incisor are preserved ( Fig. 6C View Figure 6 ; Supporting Information, Appendix S5C). The incisor is interpreted as I/2, and the alveolus as the adjacent I/3. Indeed, in frontal view, a small ventral chin foramen, mesial to the preserved incisor, could be in the same position as the intermediate foramen between the I/1 and I/2 of Merycopotamus medioximus Lihoreau et al., 2004b ( Lihoreau et al., 2004b) and reveals the existence of the I/1. The diameter of the I/2 alveolus (8.7 mm) is smaller than that of the I/3 alveolus (12.4 mm) ( Fig. 6D View Figure 6 ), unlike in Elomeryx . Only the crown of a left I/1 from SAM 4 (UM-SAM4-026, Lmd = 12 mm) documents this dental locus. There is no diastema between c and I/3. On the M12736 mandible ( Forster-Cooper, 1924: fig. 41) and UM-SAM4-014 in frontal view, the outline of the canine socket is large and clearly elliptical, hence not premolariform, in contrast to Brachyodus ( MacInnes, 1951; Dineur, 1981).
Loaeer premolars: The occlusal structure of P/1-P/2 is best preserved on the M12736 mandible, where all premolars have an accessory cuspid on their preprotocristid ( Fig. 6A View Figure 6 ; Supporting Information, Appendix S5A) like in Hemimeryx ( Lihoreau et al., 2016) . The P/1 has two separate alveoli ( Fig. 6D View Figure 6 ), whereas those of Brachyodus and Afromeryx are single-rooted. An isolated crown of a P/1 from SAM 4 (UM-SAM4-027) also possesses a fused upper part of the roots. The postprotocristid is distolingually oriented on the P/3 of M12736 ( Fig. 6A View Figure 6 ), and is clearly straight and distal in other specimens (M12721, M12723 and M12731), unlike the condition observed in Elomeryx and Merycopotamini . This tooth displays a well-preserved endoprotocristid on GSI B479 ( Pilgrim, 1912: pl. 25, fig. 6). The P/4 has a rectilinear postprotocristid centred with respect to the median axis ( Fig. 6E View Figure 6 ). It is split from the protoconid without partially fusing with the distolingually directed endoprotocristid (M12720 and UM-TOB-001; Figs 6E View Figure 6 , 7A View Figure 7 ), which is a known primitive character also present in BakaloƲia and Elomeryx ( Hellmund, 1991) . Despite the lack of distolabial cristid, a deep postprotofossid is retained. An accessory mesiolingual ridge joining the preprotocristid is only visible on the M12720 P/4 ( Fig. 6E View Figure 6 ), but not on the specimens M12721, M12731 and M12736 ( Fig. 6A View Figure 6 ). The advanced occlusal wear of three other known P/4s, including that of the mandible from Tobah ( Fig. 7A View Figure 7 ), discard observing this feature. The lingual cingulid of the P/4 does not form a clear wall, unlike in Merycopotamini ( Lihoreau et al., 2019), but it begins near the distal edge of the tooth.
Loaeer deciduous dentition: The only mandible with a dP/4 we attribute to Par. hyopotamoides (GSI B478), for which only an original drawing is available, was initially considered as Telmatodon bugtiensis by Pilgrim (1912: pl. 24, fig. 2). This deciduous tooth has a paraconid mesial to the primoconid but no postectoparacristid. This combination of characters is only found on dP/4 of Afromeryx zelteni .
Loaeer molars: Labial cuspids of lower molars are broad and less crescentic than in Gonotelma , Hemimeryx and SiƲameryx known from the Bugti Hills. This aspect is reinforced by the broad labial ribs of Parabrachyodus . The lingual cuspids are also broad, whereas they are reduced to an apex in Merycopotamini . The metaconid is always quadricrescentic ( Fig. 6F View Figure 6 ) like in SiƲameryx and Afromeryx , and it is thus clearly distinct from that of Brachyodus ( Holroyd et al., 2010) . Sixty-five per cent (17/26) of the lower molars we observed possess an ectometafossid, whereas the premetacristid is consistently present. The preprotocristid does not reach the premetacristid, which is often considered as a character of Merycopotamini ( Lihoreau et al., 2016) . Labial to the trigonid, there is a small ectostylid on the M/1 (M12731), which is rarely present on M/2 and M/3 (11%, 3/ 28 specimens; Fig. 6G View Figure 6 vs. Fig. 6H View Figure 6 , summarized in Fig. 6F View Figure 6 ) where the cingulid is often discrete. In contrast, it is protuberant on the M/3 ascribed to Telmatodon bugtiensis ( Forster-Cooper, 1924: fig. 35).
The shape of the prehypocristid is variable. It is frequently divided into two mesial arms on the M/3 on which this character is observable (57%, 8/14), whereas it more often consists in a single cristid on the M/2 (33%, 4/ 12 specimens with a prehypocrisid divided into two arms). The main arm of the prehypocristid is either oriented lingually toward the postmetafossid (UM-SAM4-015, -016 and M12734; Fig. 6G View Figure 6 ) or reaches the distal wall of the trigonid (‘median’ configuration, UM-SAM4-017; Fig. 6H View Figure 6 ). The main arm of the prehypocristid is more often median on the M/3 (71%, 10/14) than on the M/2 (33%, 4/12). A similar polymorphism of the prehypocristid is known in other bothriodontines, such as Elomeryx armatus ( Macdonald, 1956: figs 5, 7), and for the two M/3 referred to as Hemimeryx blanfordi by Lihoreau et al. (2016) ( Forster-Cooper, 1924: fig. 44 vs. pl. 6). In addition, the lingual arm of the prehypocristid, although sometimes long (M12731 and M12734), never reaches the lingual margin of the lower molars inasmuch as it is restricted by the postectometacristid and the protruding ectoentocristid (UM-SAM4-016), which tend to close the transverse valley. In this sense, the crest pattern observed in Parabrachyodus is reminiscent to that of Brachyodus (contra Dineur, 1981; Pickford, 1987) in which the lingual arm of the prehypocristid is also limited by the postectometacristid. This is in contrast to the condition seen in Libycosaurus , Merycopotamus and SiƲameryx where the lingual arm of the prehypocristid meets the lingual margin of the tooth ( Lihoreau & Ducrocq, 2007).
On almost half of the lower molars on which the pre-entocristid can be seen (48%, 13/27 M/2 and M/3; Fig. 6F View Figure 6 ), this cristid is directed towards the apex of the hypoconid. It is otherwise oriented mesially towards the trigonid wall. A possible contact between the preentocristid and the prehypocristid is materialized on three specimens by the presence of an accessory ridge distinct from the ‘endohypocristid’ known in some species of Bothriogenys ( Ducrocq, 1997) , in that its basal part is linked to the prehypocristid instead of the apex of the hypoconid (UM-SAM4-020, GSI B521 and M12725; Fig. 6I View Figure 6 ). The posthypocristid can either directly join the prehypocristulid on the M/3 and the lingual distostylid on the M/2 (UM-SAM4-015 and M12729; Fig. 6G View Figure 6 ), or form a talonid wall with the postentocristid when present (69%, 18/26 lower M/2 and M/3; UM-SAM4-013 and -020). In Bothriogenys gorringei ( Andrews & Beadnell, 1902) , the M/3 shows the same polymorphism regarding the accessory ridge of the hypoconid, but also on the direction of the posthypocristid as evidenced by Ducrocq (1997: 15). The uncommon posthypofossid (33%, 9/27 of M/2 and M/3) found on UM-SAM4-020 is accompanied by a distal accessory ridge on this tooth ( Fig. 6I View Figure 6 ).
The hypoconulid is loop-like and transversely compressed, as in all Bothriodontinae. It is also pinched mesially and aligned with the lingual cuspids, unlike Telmatodon bugtiensis ( Pilgrim, 1907: pl. 12, fig. 4) and Brachyodus onoideus ( Ducrocq & Lihoreau, 2006: fig. 1G). The morphology of this cuspid is similar to that of Anthracotheriinae on some specimens (UM-SAM4-020, M12722 and M12725; Fig. 6I View Figure 6 ), in part due to the short mesiolingual ectohypocristulid occasionally leaving its apex (44%, 7/16 of M/3; Fig. 6F View Figure 6 ). Regarding the postectostylid, the labial cingulid connects to the hypoconulid on over two-thirds of the M/3. In lingual view, we frequently observed a postentostylid on the posthypocristulid (89%, 16/18 of M/3), accompanied by a ridge closing the transverse fossa (76%, 13/17 of M/3). The M/3 UM-SAM4-020 ( Fig. 6I View Figure 6 ) displays three of those stylids arising from the marked distal cingulid, which is also well developed on the M/2 and absent on the M/3 of Hemimeryx ( Lihoreau et al., 2016) . Here, the largest of the postentostylids is, therefore, not homologous to the entoconulid of anthracotheriines. It is worth noting that a similar multiplication of postentostylids is also observed on an M/3 of S. palaeindicus from Tobah. The combination of a low crown, a postentocristid, an ectohypocristulid and a pinched hypoconulid closed by accessory cristids allowed us to recognize the only known lower jugal tooth of Par. hyopotamoides from Zinda Pir (UM-SFN-001; Fig. 7B, C View Figure 7 ).
The lengths and widths of the lower molars of Par. hyopotamoides have higher unbiased coefficients of variation than the upper molars ( Table 4), and the range of measurements for M/3 reaches 20 mm for the length. Variations of this order are expected for an anthracothere of the size of Parabrachyodus , because the same observations can be made for the molars of Libycosaurus bahri , equivalent in size to Par. hyopotamoides based on the proportions of the molars ( Lihoreau et al., 2014: table 7). There is no significant difference between the mesial (Lll1) and distal width (Lll2) of the lower molars (t-test with p <0.05; N = 38, t = –0.50, d.f. = 74, P = 0.61) even though those of the small series from SAM 4 (Supporting Information, Table S1) display a smaller width mesially than distally. In the new specimens here described, some molars have a broader talonid ( Fig. 7A View Figure 7 ), while others display a broader trigonid ( Fig. 7B View Figure 7 ). The size of the M/1 (approximated as the labiolingual length of the talonid instead of the mesiolingual length, due to the presence of the hypoconulid lobe on M/3, more elongated) is reduced compared to the M/2-M/3, in the same proportion as that which we observed on the upper molars ( Fig. 7A View Figure 7 ; Table 4). Furthermore, the labiolingual length of the talonids of Telmatodon and Hemimeryx falls within the lower limit of that of Par. hyopotamoides with which they could be confused. Due to the paucity of specimens attributed to the formers, we consider that this difference cannot be assessed with confidence here.
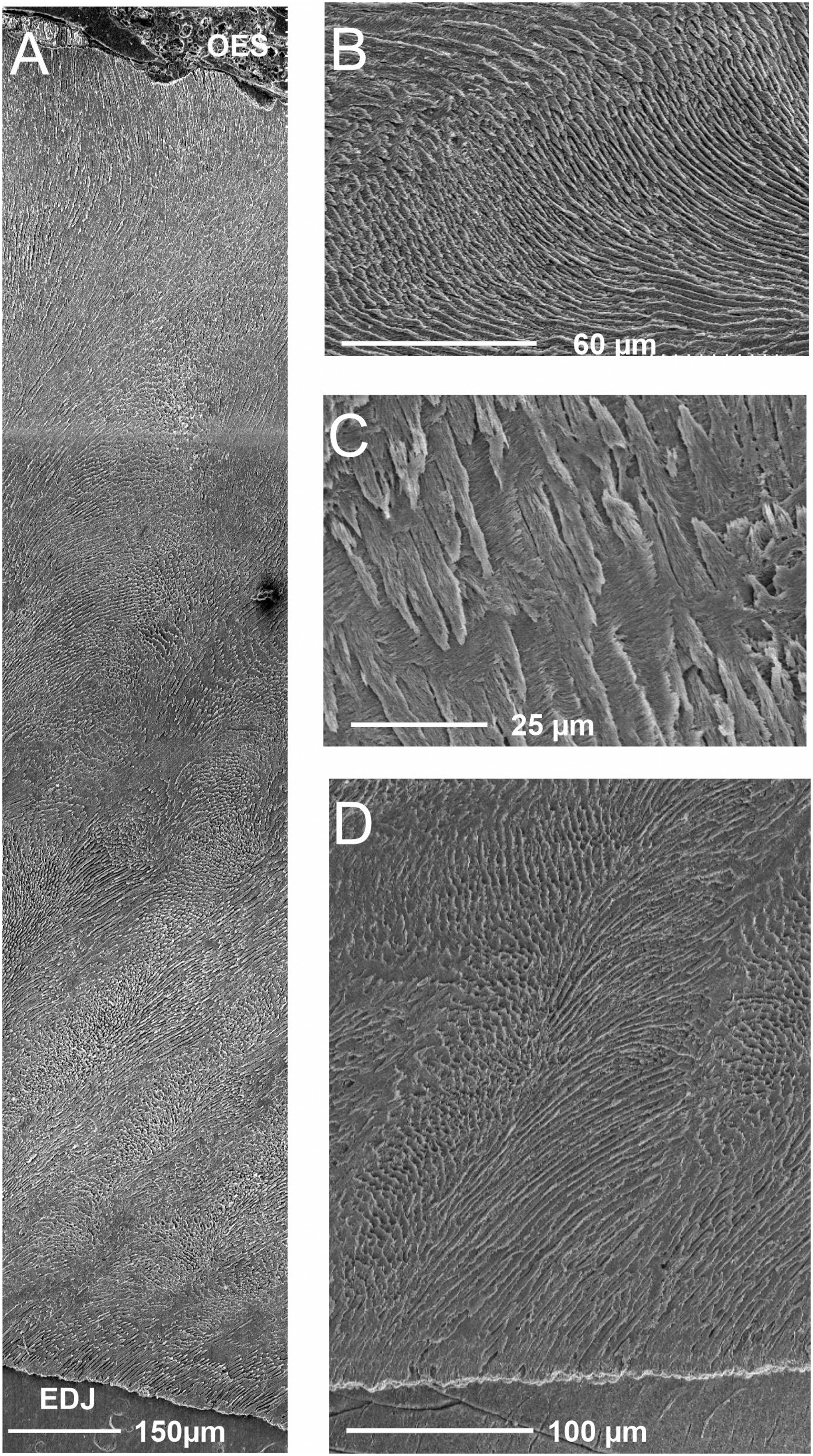
Enamel microstructure
On the vertical section, enamel thickness averages 2210 µm with maximal measured value of 2381 µm. This is higher than what has been observed so far in bothriodontines, notably with the thick enamel of Brachyodus onoideus , which is close to 1500 µm ( Alloing-Séguier et al., 2014). This enamel thickness is close to values measured for Paenanthracotherium bergeri Scherler et al., 2018 and the early hippopotamines Saotherium Boisserie, 2005 and Hexaprotodon Falconer & Cautley, 1836 ( Alloing-Séguier et al., 2014; see also: Scherler et al., 2018, for Paenanthracotherium attribution).The Schmelzmuster ( Fig. 8A View Figure 8 ) shows an inner zone with little development of inner radial enamel (4%), a large decussation zone with Hunter–Schreger Bands developed for 75% of the whole thickness, and a large outer zone of radial enamel (21%). Due to the weak development of inner radial enamel, the Schmelzmuster can be regarded as bizonal, following Alloing-Séguier et al. (2014). In the decussation zone, bands are from 45 to 117 µm (with a mean of 84.8 ± 25 µm) with about 12 to 17 prisms per band. There is a high decussation angle with several transitional prisms (almost five; Fig. 8B View Figure 8 ). Bands vary slightly in width from the EDJ to the OES ( Fig. 8A View Figure 8 ) displaying a regular aspect. They are strongly bent in the inner third of the enamel thickness, rising at an angle of 57 ± 8° with the EDJ and reaching the outer radial enamel perpendicularly to the OES. We did not observe anastomosis or bifurcation of the bands.
On horizontal sections, the Schmelzmuster displays radial enamel with some blurry decussations near the EDJ that quickly disappear to become strict radial enamel with some synchronous prism undulations (three) in the inner two-thirds of enamel thickness. Radial enamel then becomes straight to the OES.
Mean prism diameter is 4.5 ± 0.4 µm (between 4 and 5.2 µm). Prism angles with EDJ equal ~54.6 ± 6° and reach OES perpendicularly. IPM forms inter-row sheets in the HSB zone and in the short inner radial zone. IPM crystallites are nearly perpendicular to prisms ( Fig. 8C View Figure 8 ). In the outer radial portion, IPM forms closed coats.
Comparisons indicate more similarities with Merycopotamini than with Brachyodus , notably due to the marked development of the outer radial enamel, the large and blurry HSB, the low number of prism undulations in horizontal section and the development of inter-row sheets from the EDJ ( Fig. 8D View Figure 8 ).
PHYLOGENETIC ANALYSIS
The heuristic searches yielded 208 equally mostparsimonious trees of 1558 steps each. A prominence of homoplasic characters has been obtained (consistency index = 0.195) as expected for an intraordinal analysis, but the phylogenetic results are quite structured (retention index = 0.625), as summarized by the well-resolved strict consensus tree ( Fig. 9 View Figure 9 ). The major polytomies concern relationships in the Hip popotamidae + Bothriodontinae clade erected by Lihoreau et al. (2015). This clade is well supported [Bremer index (BI) = 5] by 24 non-ambiguous synapomorphies of which five are also non-homoplasic among artiodactyls, including the Schmelzmuster composed of three layers (1681, with intraclade reversions) and the presence of bent instead of straight HSB (1761). The trees differ from each other mainly in the resolution of the relationships between the four subclades ( Bothriodon + Aepinacodon ), (( Brachyodus spp. + Bothriogenys andreaesi ) + Bo. gorringei + Bo. fraasi ), ( Bo. orientalis + Epirigenys + Hippopota midae) and the clade A, including Parabrachyodus hyopotamoides . This uncertainty is consistent with the need for a revision of the evolutionary history of Brachyodus and Bothriogenys ( Lihoreau et al., 2019) whose monophylly is not supported here.
Parabrachyodus hyopotamoides is hence positioned among Bothriodontinae in a separate clade from the one including Brachyodus (clade A), supported by nine non-ambiguous traits (BI = 3). Its topology consists of a branching sequence with the Merycopotamini sensu Lihoreau et al. (2016) as crown group, with the enigmatic BakaloƲia as the earliest offshoot (node A), including the monophyletic genus Elomeryx (node B), Par. hyopotamoides and the other endemic taxa from the Bugti Hills (node D). It is worth noting that the Merycopotamini ( Afromeryx , Hemimeryx , Merycopotamus , Libycosaurus and SiƲameryx ) are resolved here as a paraphyletic group because of the newly added genus Gonotelma , which is the sistertaxon of Afromeryx (node H). These two genera are only united by synapomorphies of their lower molars (670, 801, 831). Furthermore, the non-ambiguous synapomorphies defining the tribe Merycopotamini ( Lihoreau et al., 2016) appear sequentially in our topology: the pinched hypoconulid (791), the labially connecting preparacrista of the parastyle (1302) and the absence of ectocristyle (1331) for clade A; the presence of accessory cuspids on the preprotocristid of lower premolars (191), which is also non-homoplasic at node B; and the loss of the connection between the premetacristid and the preprotocristid of lower molars (501), the loss of the paraconule of upper molars (1271) and the marked development of outer radial enamel (1711) for clade D.
ThecladeD,including( Parabrachyodus + Telmatodon ), Gonotelma and the Merycopotamini sensu Lihoreau et al. (2016) , is among the strongest ones (BI = 4). It is defined by 13 non-ambiguous synapomorphies (two of which are above-mentioned), including the more extensive development of the I/1 (42), the preparacrista composed of two mesial ridges on P1/-P3/ and the multiplication of accessory cusps on their postparacrista (931, 941), the loss of the junction between the protocone and the metaconule, characteristic of the upper molars of Elomeryx (1190) , or the presence of two external foramens on the mandible instead of one (1592). It is relevant to mention that the enamel microstructure and the occlusal morphology of the dP3/ strongly supports this clade with almost half of the non-ambiguous synapomorphies (six on a total of 13 synapomorphies). Hence, three of them are related to the enamel microstructure (1680, 1701 and 1711), namely that the Schmelzmuster is composed of two layers (1680), HSB occupy less than a quarter of its surface (1701) and the marked development of outer radial enamel (1711). Three non-ambiguous traits are related to character states of the dP3/ (2131, 2191, 2230), known by only one specimen for Parabrachyodus . Thus, there is an entostyle (2191) and no connection between the protocone and the metacone (2230). The bicuspidate anterior lobe of the dP3/ (2131) is also a non-homoplasic trait.
The resolution in clade D is good despite the low scoring for Gonotelma shahbazi (79/224 characters), Telmatodon bugtiensis (75/224 characters) and T. orientalis (90/224 characters). Altogether, Parabrachyodus and its ‘relatives’ from the Bugti Hills appear paraphyletic in this first attempt to include them in a cladistic analysis. Telmatodon forms a clade with Par. hyopotamoides (node E), supported by four non-ambiguous synapomorphies (631, 1081, 1431 and 1520), such as the gentle slope of the metaconule (1520) compared to all other anthracothere groups (non-homoplasic in this context). Parabrachyodus hyopotamoides is related to Telmatodon bugtiensis (node F) rather than to T. orientalis (both Telmatodon species are here considered valid due to the nature of the material attributed to each of them, see Supporting Information, Appendix S2) by only two non-ambiguous synapomorphies, one of which is also non-homoplasic and concerns the lingual development of the postectoprotocrista on the M3/ (1171).
The next node (G, BI = 2), grouping Gonotelma shahbazi , Afromeryx zelteni and the rest of Merycopotamini , is supported by four non-ambiguous synapormorphies, including the non-ambiguous anterior position of the main palatal foramen (1661), the abrupt change in direction of the preprotocristid of P/4 (442) and, especially, the lingual position of the postprotocristid of the P/4 (411), which is non-homoplasic at the artiodactyls scale. There are also two non-ambiguous synapomorphies supporting the clade I (BI = 4) that are related to the P/4 morphology: a fusion of the postprotocristid and endoprotocristid (322) converging with the subclade ( Brachyodus spp. + Bothriogenys in part) and the presence of a hypoconid (391). It is worth noting that SiƲameryx palaeindicus and S. africanus are brought together (clade J) only by the presence of a paraconule (1270), which is considered in this topology as a reversion within clade D, occurring also for Parabrachyodus hyopotamoides . Specifically, it is one of the six non-ambiguous autapomorphies of Par. hyopotamoides , notably including the already mentioned convergences with Anthracotheriinae , such as an ectohypocristulid developed on the M/3 (772) and the presence of a protostyle on upper molars (1511).


| UM |
University of Marburg |
| SAM |
South African Museum |
| GSI |
Geological Survey of India |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Parabrachyodus hyopotamoides
| Gernelle, Killian, Lihoreau, Fabrice, Boisserie, Jean-Renaud, Marivaux, Laurent, MéTais, Grégoire & Antoine, Pierre-Olivier 2023 |
Parabrachyodus hyopotamoides (Lydekker)
| Bhandari A & Mohabey DM & Bajpai S & Tiwari BN & Pickford M 2010: 76 |
Parabrachyodus hyopotamoides (Lydekker)
| Lindsay EH & Flynn LJ & Cheema IU & Barry JC & Downing K & Rajpar AR & Raza SM 2005: 16 |
Parabrachyodus hyopotamoides (Lydekker)
| Pickford M 1987: 316 |
Telmatodon
| Pickford M 1987: 320 |
Brachyodus manchharensis
| Pickford M 1987: 309 |
| Prasad KN 1964: 9 |
Parabrachyodus
| Viret J 1961: 948 |
Brachyodus gandoiensis
| Forster-Cooper C 1924: 27 |
Brachyodus platydens
| Forster-Cooper C 1924: 29 |
Brachyodus orientalis
| Forster-Cooper C 1924: 32 |
Brachyodus indicus
| Forster-Cooper C 1924: 32 |
Gonotelma major
| Forster-Cooper C 1924: 49 |
Parabrachyodus obtusus (Forster-Cooper)
| Forster-Cooper C 1915: 404 |
Brachyodus pilgrimi
| Forster-Cooper C 1913: 516 |
Brachyodus africanus
| Pilgrim GE 1912: 59 |
Brachyodus giganteus (Lydekker)
| Andrews CW 1899: 484 |
| Deperet C 1895: 407 |
Anthracotherium hyopotamoides
| Lydekker R 1883: 152 |
Hyopotamus giganteus
| Lydekker R 1883: 160 |