
Huracan coffeyi ( Dalquest, 1986 )
|
publication ID |
https://doi.org/10.1206/3996.1 |
|
persistent identifier |
https://treatment.plazi.org/id/3F7387AE-FFFE-FFBB-FE1F-B357FB45FA6D |
|
treatment provided by |
Felipe |
|
scientific name |
Huracan coffeyi ( Dalquest, 1986 ) |
| status |
|
Huracan coffeyi ( Dalquest, 1986)
Agriotherium schneideri: Savage, 1941: 696 .
Agriotherium sp. : Harrison, 1983: 22.
Agriotherium schneideri (in part): Hunt, 1998: 184.
“ Agriotherium ” schneideri: Jiangzuo et al., 2021 .
TYPE SPECIMEN: MWSU 12147 , a partial mandible preserving c-m3, from Coffee Ranch .
REFERRED SPECIMENS: In addition to the holotype, and specimens listed in Frick (1926), Harrison (1983), and Dalquest (1986), the following specimens also now are referred to this species. Cranium: Guymon Formation: AMNH F: AM 30399 (with associated mandible, shortly mentioned by Frick, 1937) , AMNH F: AM 50030 ; Quiburis Formation: AMNH F: AM 49372 , AMNH F: AM 76001 , AMNH F: AM 76005 , AMNH F: AM 76006 , AMNH F: AM 76007 . Mandible: Coffee Ranch: AMNH F: AM 23364 , AMNH F: AM 23368 ; Guymon Formation: AMNH F: AM 50032 , AMNH F: AM 50033 ; Quiburis Formation: AMNH F: AM 76005 , AMNH F: AM 76008–10 , AMNH F: AM 76012 , AMNH F: AM 76012 a, AMNH F: AM 76020 , AMNH F: AM 76031 , AMNH F: AM 144551 . Isolated teeth or jaw fragments: Coffee Ranch: AMNH F: AM 23365 , AMNH F: AM 23367 , AMNH F: AM 23369 a, AMNH F: AM 23369 b, AMNH F: AM 23369 c, AMNH F: AM 23370 , AMNH F: AM 23371 , UCMP 30517 View Materials , UCMP V24025 , UCMP
A E1 E E3B
B3
C F2
D1 G D2 5 cm
V 24027, UCMP V 24026; Guymon Formation: AMNH F: AM 50034, AMNH F: AM 50034a,, AMNH F: AM 50035–50037, AMNH F: AM 50037a, AMNH F: AM 50038, AMNH F: AM 50039; Quiburis Formation: AMNH F: AM 145923, AMNH F: AM 76017, AMNH F: AM 76005, AMNH F: AM 76006, AMNH F: AM 76007, AMNH F: AM 49372, AMNH F: AM 76035, AMNH F: AM 76013, AMNH F: AM 76032, AMNH F: AM 76019, AMNH F: AM 76021, AMNH F: AM 76020, AMNH F: AM 76033, AMNH F: AM 76024, AMNH F: AM 76034, AMNH F: AM 76023, AMNH F: AM 76011, AMNH F: AM 76016, AMNH F: AM 76020, AMNH F: AM 76018C, AMNH F: AM 76018D, AMNH F: AM 76015; AMNH F: AM 146568; AMNH F: AM 146569; AMNH F: AM 146468–502; AMNH F: AM 146570; AMNH F: AM 146571; Big Sandy Formation: AMNH F: AM 49380, AMNH F: AM 49382, AMNH F: AM 49376, AMNH F: AM 49375, AMNH F: AM 49379, AMNH F: AM 49378. Humerus: Coffee Ranch: UCMP 31818, Quiburis Formation: AMNH F: AM 76106, AMNH F: AM 76070, AMNH F: AM 76071, AMNH F: AM 76072, AMNH F: AM 76073, AMNH F: AM 146513. Radius: Ash Hollow Formation: UNSM 76013; Quiburis Formation: AMNH F: AM 146513, AMNH F: AM 76078, AMNH F: AM 76077, AMNH F: AM 76008, AMNH F: AM 146512, AMNH F: AM 146456; Guymon Formation: AMNH F: AM 50042. Ulna: Quiburis Formation: AMNH F: AM 76082, AMNH F: AM 76081, AMNH F: AM 76080, AMNH F: AM 76079, AMNH F: AM 76008, AMNH F: AM 146511, AMNH F: AM 146457; Guymon Formation: AMNH F: AM 50042. Carpal bones: Guymon Formation: AMNH F: AM 146450 (scapholunar); Coffee Ranch: AMNH F: AM 146452 (cuneiform), AMNH F: AM 112588 (pisiform); Quiburis Formation: AMNH Red (Redington)117-1831 (pisiform). Femur: Quiburis Formation: AMNH F: AM 76090, AMNH F: AM 146513, AMNH F: AM 146503, AMNH F: AM 76105, AMNH F: AM 76085, AMNH F: AM 76086. Tibia and
Fibula: Quiburis Formation: AMNH F: AM 146513, AMNH F: AM 146514, AMNH F: AM 76092, AMNH F: AM 76090, AMNH F: AM 76087, AMNH F: AM 76097, AMNH F: AM 76096, AMNH F: AM 76095, AMNH F: AM 76008, AMNH F: AM 76014, AMNH F: AM 76014. Astragalus: Quiburis Formation: AMNH F: AM 76103, AMNH F: AM 146508, AMNH F: AM 76102, AMNH F: AM 146509, AMNH F: AM 76014, AMNH F: AM 76014; Coffee Ranch: AMNH F: AM 146454. Calcaneum: Quiburis Formation: AMNH F: AM 146510, AMNH F: AM 76097, AMNH F: AM 76008, AMNH F: AM 76014; Big Sandy Formation: AMNH F: AM 146467; Coffee Ranch: UCMP 31837. Mesocuneiform: Coffee Ranch: AMNH FM 146707. Cuboid: Quiburis Formation: AMNH F: AM 76014, AMNH F: AM 146459; Coffee Ranch: UCMP 30519. Metapodial: MC 1, Big Sandy Formation, AMNH F: AM 146563, AMNH F: AM 146564, AMNH F: AM 146458. MC 2, Quiburis Formation, AMNH F: AM 76084. MC 4, Quiburis Formation, AMNH F: AM 76101, AMNH F: AM 76008, AMNH F: AM 146505, AMNH F: AM 146564. MC 5, Quiburis Formation, AMNH F: AM 76100, AMNH F: AM 69325A, AMNH F: AM 69327. MT 1, Quiburis Formation, AMNH F: AM 76099, AMNH F: AM 76014, Big Sandy Formation, AMNH F: AM 146564. MT 2, Quiburis Formation, AMNH F: AM 146513, AMNH F: AM 76014, AMNH F: AM 146460. MT 3, Quiburis Formation, AMNH F: AM 146513, AMNH F: AM 76014. MT 4, Quiburis Formation, AMNH F: AM 146513, AMNH F: AM 76014, Big Sandy Formation, AMNH F: AM 146466. MT 5, Quiburis Formation, AMNH F: AM 76029, AMNH F: AM 76030, AMNH F: AM 76087, AMNH F: AM 76014.
EMENDED DIAGNOSIS: Huge ailuropodine bear with hypercarnivorous craniodental features. Cranial condylobasal length 380–515 mm. Mandible often with premasseteric fossa; anterior premolars single-rooted (P3/p3 occasionally double rooted); P4 longer than M1, variably with subdivided hypocone; p4 with distinct anterior and posterior cingulid cusps but no medial ridge; m1 with elongated paraconid and distinct metaconid, mesoconid and distal entoconid; humerus lacking entepicondylar foramen.
DIFFERENTIAL DIAGNOSIS: Differs from H. schneideri in having slightly larger dental size; narrower M2 with proportionally longer talon; p4 with less distinct anterior accessory cusp and weaker medial ridge; m1 with stronger mesoconid and larger posterior entoconid; m2 with stronger metaconid and better subdivided entoconid. Differs from Huracan qiui in having more reduced P4 anterior hypocone, shorter M1 and M2.
OCCURRENCE: Known from the late Hemphillian NALMA (Hh3). In the Great Plains of the U.S., this species has been found at USGS Locality Ml 430 in the Coso Formation in Inyo County, California; the Carlin High Quarry in the Hay Ranch Formation in Elko County, Nevada; the Clay Bank and Bird Bone Quarries in the Big Sandy Formation in Mohave County, Arizona; the Old Cabin, Redington, and Camel Canyon quarries in the Quiburis Formation in Pima and Pinal Counties, Arizona; and at Coffee Ranch, Guymon-Optima quarries, Axtel Quarry, Edson Quarry, Lost Quarry and the nearby UNSM Locality SM- 101 in the Ogallala Formation in Texas and Oklahoma ( Savage, 1941; Harrison, 1983; Dalquest, 1986; Hunt, 1998; Tedford et al., 2004). An isolated M2 from the Johnson Member of the Snake Creek Formation in Sioux County, Nebraska is also from Hh3 (R. Tedford, in note associated with this specimen in the AMNH collections).
major palatine foramen
C
DESCRIPTION: This species was previously known only by isolated teeth and several mandibles. Additional study of the extensive AMNH Frick Collection of Fossil Mammals brings many new specimens of this species to light, including seven crania, 14 mandibles, and numerous isolated teeth and postcranial bones. Unfortunately, nearly all the crania were found in Arizona and Oklahoma from sites in which most fossils are crushed. The crania and mandibles are generally large in size and vary substantially in morphology. Complete crania permitting measurement of total length and condylobasal length are rare, but those have an average of 490 mm for cranial total length (n = 2) and 440 mm for condylobasal length (n = 4; table 5). However, complete mandibles are abundant, with a mean length of 318.5 mm (n = 8; table 11). Based on the ratios of 1.47 and 1.37 for cranial total length/ mandibular length and condylobasal length/mandibular length, respectively, from an individual associated cranium and mandible of AMNH F:AM 76005, the range of cranial total lengths and condylobasal lengths in this species are estimated at 411–553 mm and 383–515 mm, respectively. The large size range and bimodal distribution of mandibular size suggest considerable sexual dimorphism in this species.
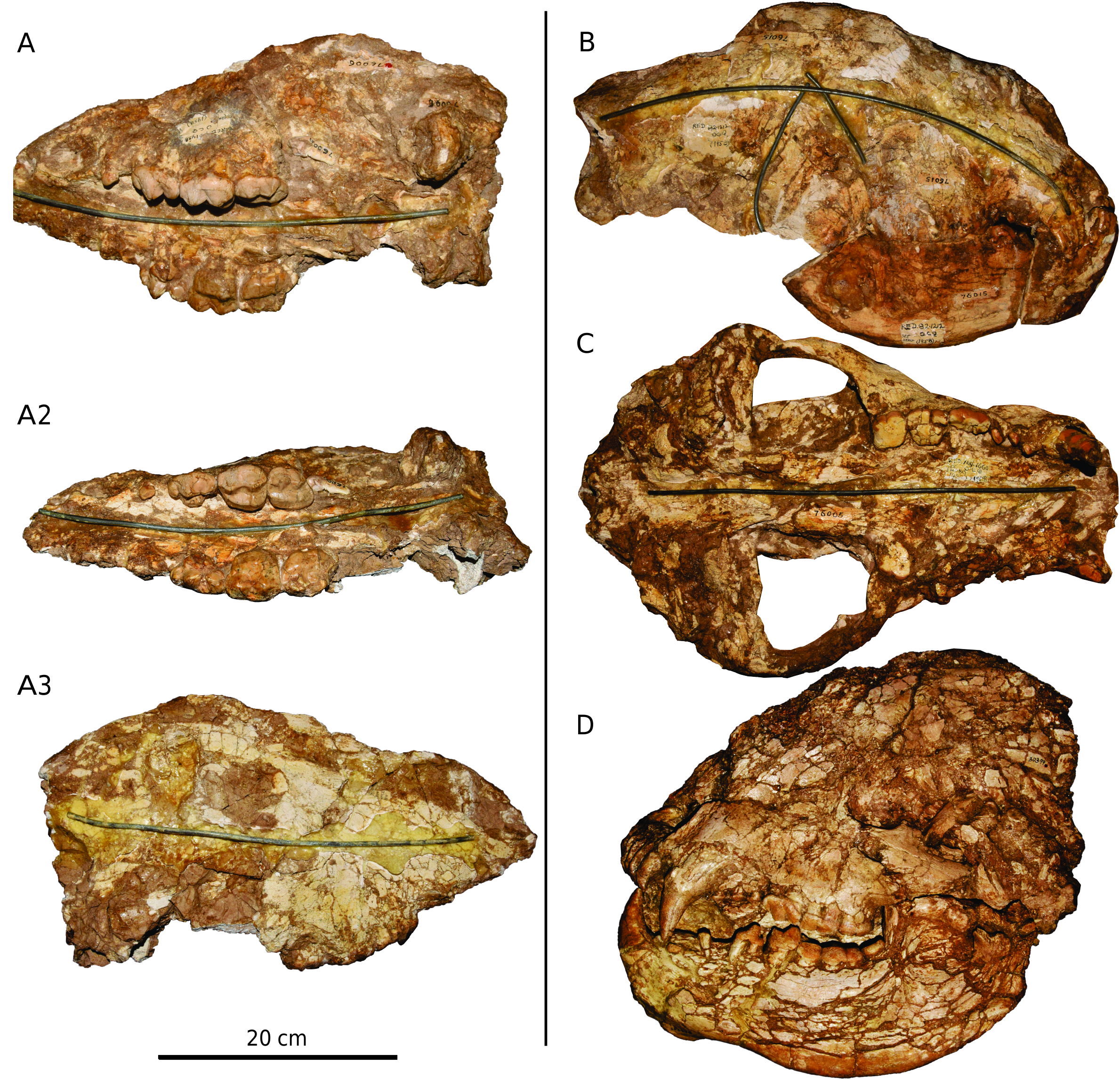
The only skull that is not severely compressed is AMNH F:AM 49372 from the Camel Canyon Quarry, Quiburis Formation, Arizona ( fig. 8 View FIG ). Our description of the cranium thus is based primarily on this specimen, although some of the other crushed crania are informative and figured for comparison (e.g., AMNH F:AM 76015, fig. 9B View FIG ).
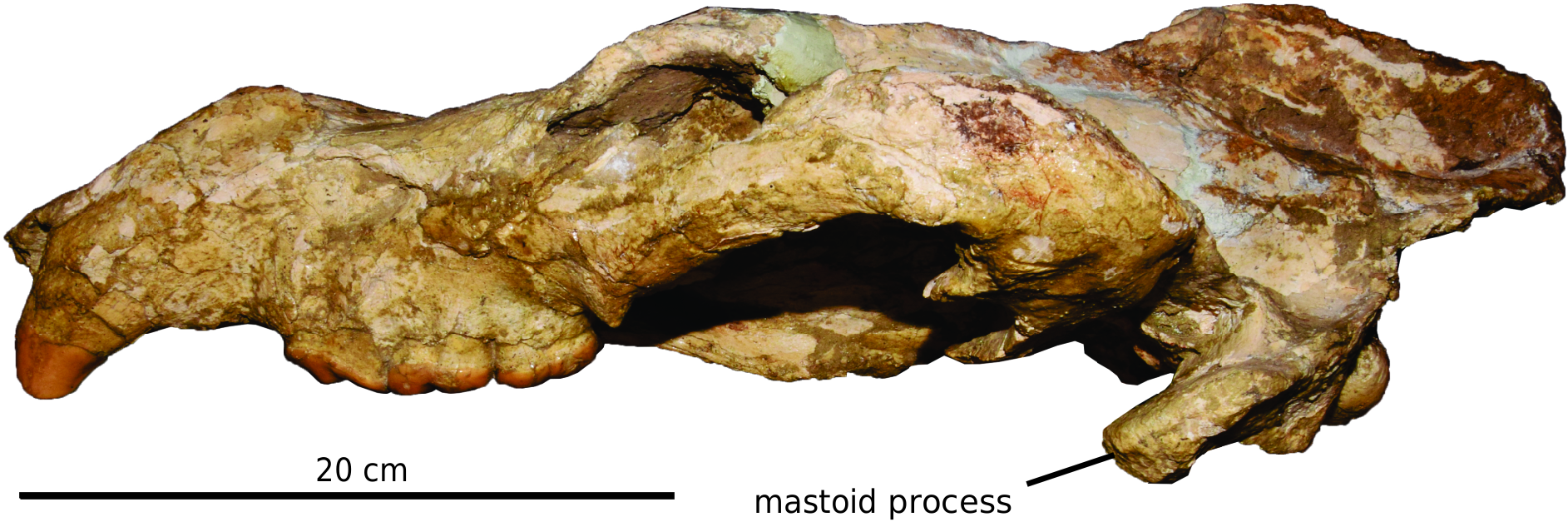
DORSAL VIEW: The rostrum is robust, with a distinct postcanine constriction. The forehead is rather wide and the postorbital process of the frontal is moderately developed. The temporal ridges unite into the sagittal crest in the middle (anteroposteriorly) part of the braincase. The zygomatic arch reaches its widest point at the level of the glenoid fossa. The mastoid process is well developed and extends far laterally, surpassing the lateral extent of the squamosal shelf.
LATERAL VIEW: The cranium in AMNH F:AM 49372 is dorsoventrally compressed postmortem. What can still be seen is that the anterior border of the orbit lies at the level of the M2 metacone. The mastoid process extends farther ventrally than does the postglenoid process. The paroccipital process is weak and not posteriorly extended. The squamosal shelf is not dorsally convex. The sagittal crest is strongly developed at the posterior part of the braincase.
VENTRAL VIEW: The anterior border of the rostrum is arched. The palate is not flat, as it is distinctly convex at the level of canine and concave at the level of M2. This pattern is very similar to that of the extant sloth bear Ursus ursinus . The palatine fissure is small, located at a level between I3 and the canine. The anterior palatine foramen is located at a level between M1 and M2; the rostrum also reaches its widest point at this level. The anterior end of the zygomatic arch is located at the M2. The ventral border of the bony choana is located at the level of the M2 posterior border. The postglenoid process is moderately developed, whereas the preglenoid process is weak. The mastoid process is greatly expanded laterally. The paroccipital process is located at the base of the mastoid process, lateral to the auditory bulla. The bulla is poorly preserved, but the external meatus is well developed and the bulla is slightly inflated in the middle part. The postglenoid foramen is located at the level of the postglenoid process, distinctly medial to the level of the paroccipital process.
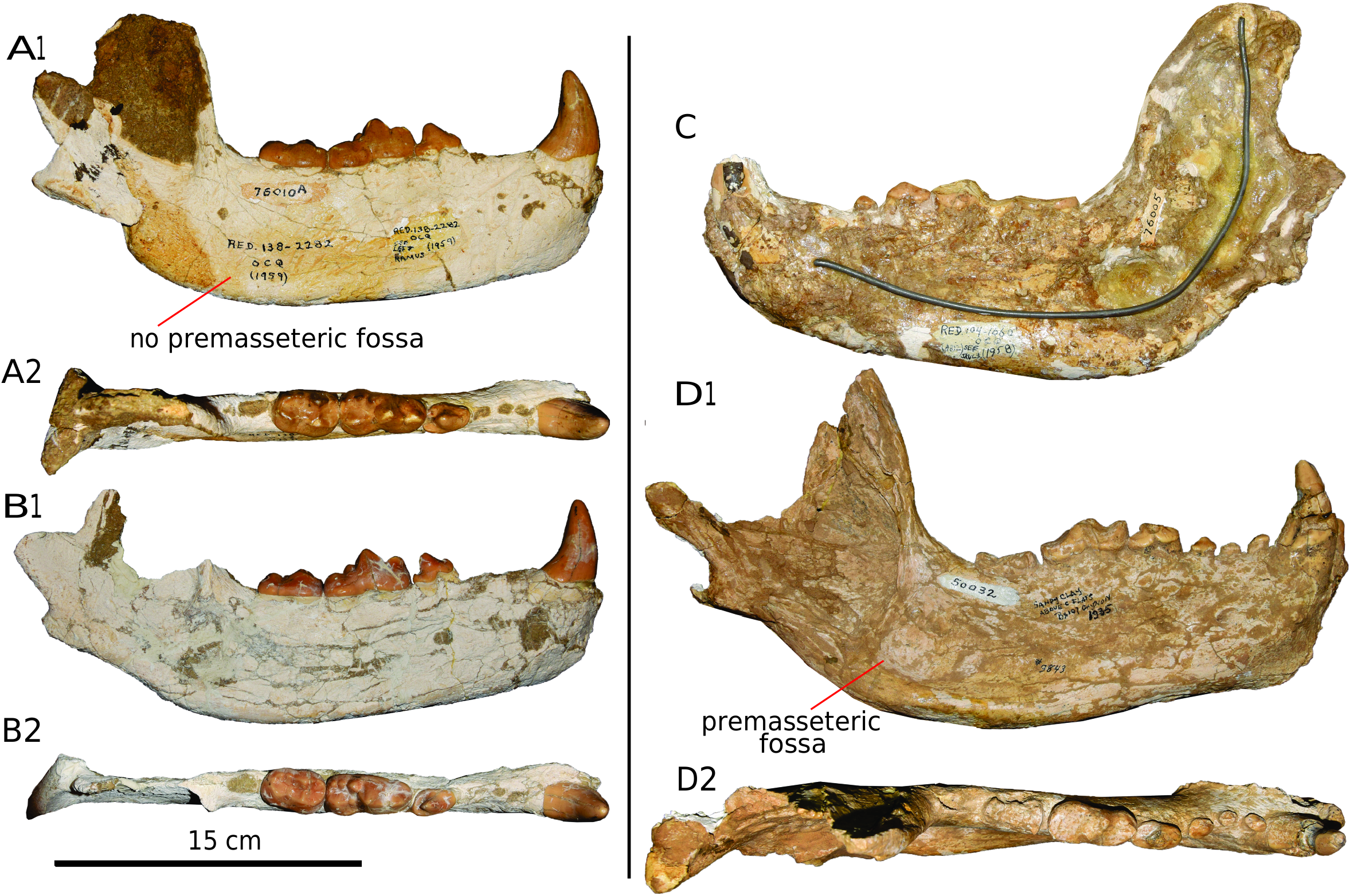
MANDIBLE: No complete and well-preserved mandibles are known. Our mandible description is mainly based on three of the most complete specimens (AMNH F:AM 76005, AMNH F:AM 76010, and AMNH F:AM 144551) from the Quiburis Formation ( fig. 10 View FIG ).
AMNH F:AM 76005 is a laterally crushed skull with associated mandible. The mandible preserves a nearly complete contour, though details cannot be clearly seen. The two most distinctive characters are the presence of a well-demarcated premasseteric fossa and the high positions of the mandibular condyle and angular process. The premasseteric fossa is rounded and confined to the posterior part of the horizontal ramus. The mandibular condyle is located significantly higher than the toothrow, and the angular process is located slightly lower than the toothrow. The coronoid process is well developed and posteriorly inflected.
The horizontal ramus of AMNH F: AM 76010 is in good condition, but the ascending ramus is broken. The horizontal ramus of AMNH F: AM 76010 has a uniform height along its A-P length, whereas in AMNH F: AM 76005 the anterior part of the horizontal ramus is lower. The most noteworthy difference between AMNH F: AM 76010 and AMNH F: AM 76005 is the total absence of a premasseteric fossa in AMNH F: AM 76010 . This character is variable within the species, as the premasseteric fossa is present in 16 in 18 specimens examined, and in seven it is only weakly developed (it is absent in the other two). There are three mental foramina. The anterior is the largest and is located at the level of the p1; the two posterior ones are nearly the same size and located close to each other below and between p2 and p3. The mandibular condyle and angular process are elevated in position relative to the mandibular corpus in AMNH F: AM 76010 and AMNH F: AM 76005 . AMNH F: AM 144551 is generally similar to AMNH F: AM 76010 , but there is a weak premasseteric fossa present in AMNH F: AM 144551 .
DENTITION: Our description is based on a summary of all the specimens we examined, with intraspecific variation noted where applicable. The teeth of AMNH F:AM 49372 are lost or worn.
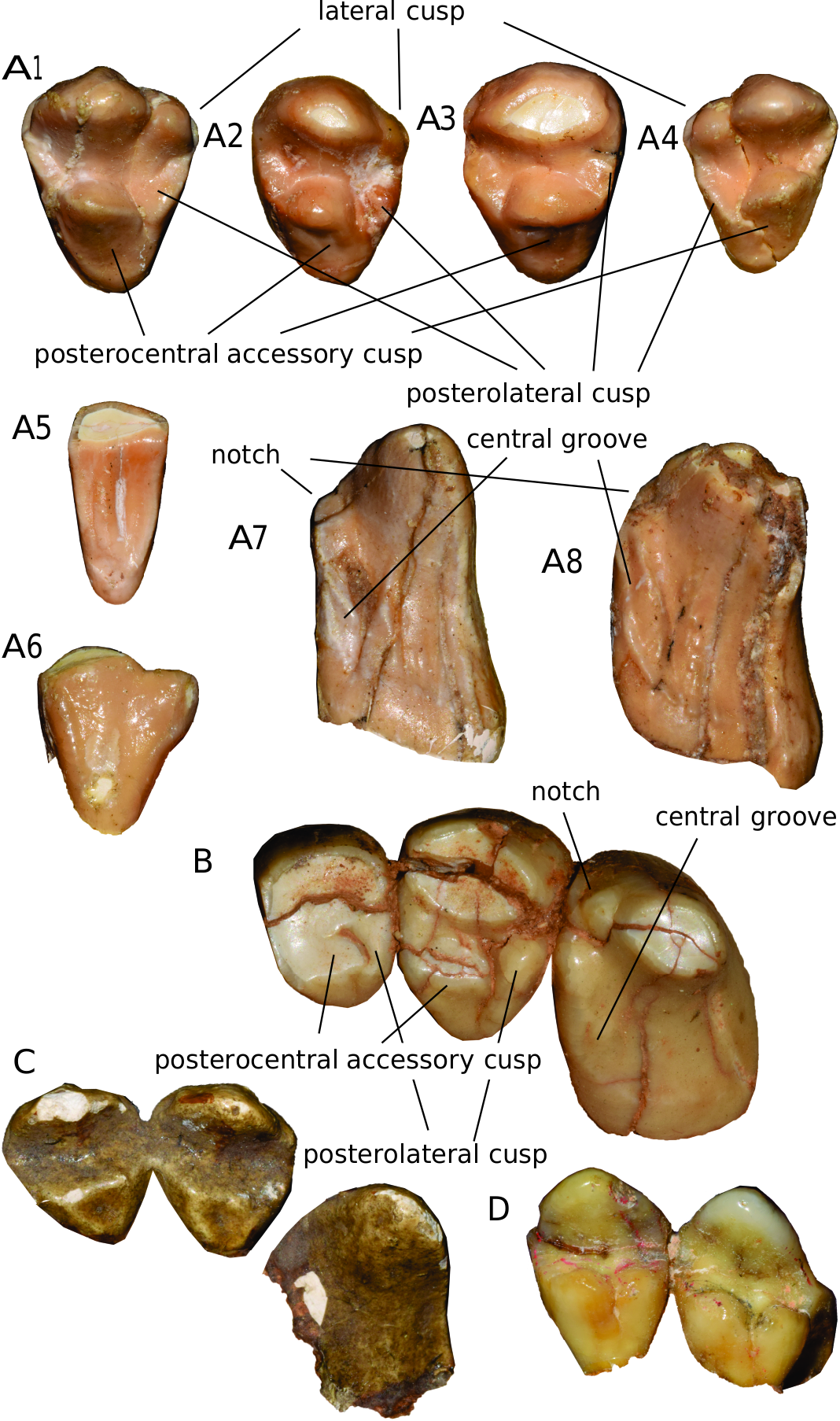
I1 and I2 are represented mainly by isolated teeth ( fig. 11 View FIG ). I1 and I2 are not distinguishable from each other due to their generally similar morphology and the lack of directly associated specimens within a single skull for intraindividual comparison. In I1/2 the posteromedial accessory cusp is strongly developed, and a very large accessory cusp (here termed the posterocentral accessory cusp) is separated from the posteromedial accessory cusp. The postero-
A B
C
10 cm
lateral accessory cusp is small in size, is much weaker than the other accessory cusps, and is ridgelike. The main cusp is often subdivided on the lateral side, but this character seems to be variable intraspecifically. I3 is distinctly larger than I1 and I2. Both medial and lateral ridges are present. The medial cingulum is strongly developed, and there is a small notch between the medial ridge and medial cingulum, and a distinct groove along the central line of the medial cingulum, as in Indarctos .
P1–P3 are small, buttonlike, and always present. P1 is single-rooted. P2 is mostly single rooted, but occasionally with fused two roots (in one of five specimens). P3 is more variable, with a single root most frequently observed (two of four cases), but with some specimens having two partially to fully separated roots (one separated, and one partially fused).
The P4 parastyle is large ( fig. 6 View FIG ). The P4 buccal contour is generally straight or shows little concavity. The lingual lobe is moderate in size, and the hypocone is variable in its morphology, typically subdivided into two cusps, with the anterior one often smaller. M1 varies in shape from square to slightly trapezoidal. The width across the metacone is always wider than that across the paracone. The anterior border of M1 is concave in most specimens. The parastyle and metastyle are often absent, but can be present and distinct from each other in some specimens. The protocone is occasionally weakly subdivided into two cusps. The direction of the postprotocrista most often diverges from the paracone-metacone axis, but sometimes parallels this axis. A small transverse ridge between the paracone and protocone is often present. A medial ridge of the metaconule is mostly present. M2 always has a talon, but it varies in size. The talon can be moderately elongated (e.g., AMNH F:AM 76006, fig. 6 View FIG : B1) or very short (e.g., AMNH F:AM 49372, fig. 6 View FIG : B3); the latter condition is not much different from that of Agriotherium . The buccal concavity between the paracone and metacone is always present. A small medial branch of the anterior ridge of the paracone (RPa1.2 of Jiangzuo et al., 2019), a small transverse ridge between the paracone and protocone, and a medial ridge of the metaconule are all only occasionally present.
The i1 and i2 are simple, dominated by a single main cusp. The i3 bears a large lateral accessory cusp.
Like the upper premolars, lower p1–p3 are generally single-rooted, and always rounded and buttonlike in shape. The p1 and p3 are always present, whereas p2 is occasionally absent (in two of 16 specimens). The p3 is occasionally double rooted (two of 10 specimens), but otherwise is typically single rooted.
The p4 has a triangular occlusal outline, with a buccal concavity in most specimens ( fig. 7 View FIG : B1, 5, 8, 12). A distinct anterior accessory cusp is nearly always present (20 of 22 specimens) and can be moderate to strong. The hypoconid (posterior cingulid cusp) also is generally present. A medial ridge of the main cusp is generally absent, but indistinctly occurs in some specimens (3 of 21 specimens). The m1 is elongate, especially so with its anteriorly directed paraconid; a distinct lingual concavity often occurs between the paraconid and protoconid. There are always three successively smaller cusps, from anterior to posterior, on the posterolingual side of m1, representing the metaconid and subdivided entoconid, a typical Indarctos - like pattern ( Qiu and Tedford, 2003; Jiangzuo and Flynn, 2019). A well-developed mesoconid is situated between the protoconid and hypoconid, again similar to the condition in Indarctos . The m2 has a massive trigonid that is wider than the talonid. The m2 metaconid is always well developed, and often is similar in height to that of the protoconid. An m2 premetaconid is sometimes present; the two m2 entoconids (subdivided entoconid) are always well separated. The m3 is oval to triangular in shape, sometimes with a metaconid present.
POSTCRANIAL SKELETON: A large number of postcranial bones of this species are known from the Quiburis Formation. However, nearly all these bones are crushed, limiting the anatomical information that can be extracted from them.
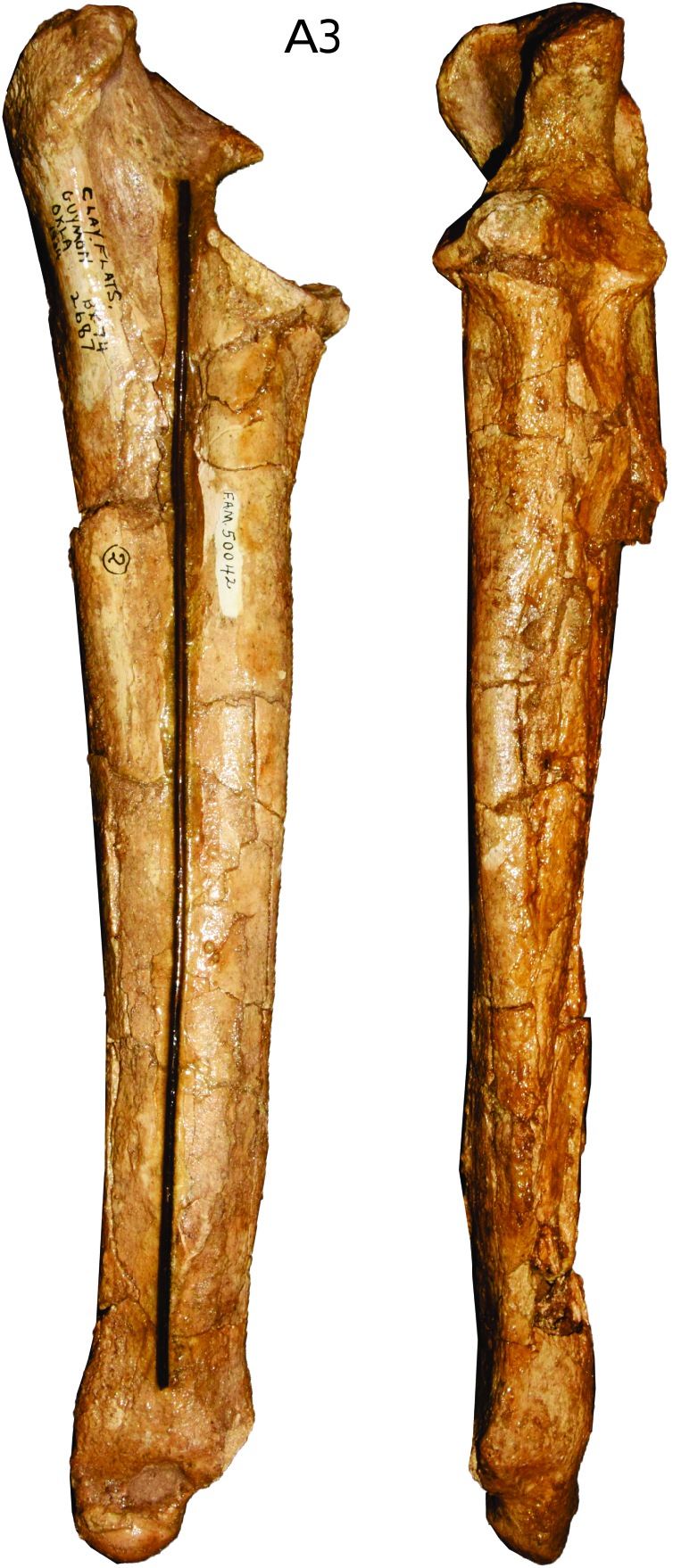
SCAPULA: The scapula is extremely rarely preserved in this species—only a broken distal scapula fragment association with a radius and ulna, AMNH F:AM 50042, is known from the Guymon Formation, Texas. The distal articular facet for the humerus is elongated anteroposteriorly.
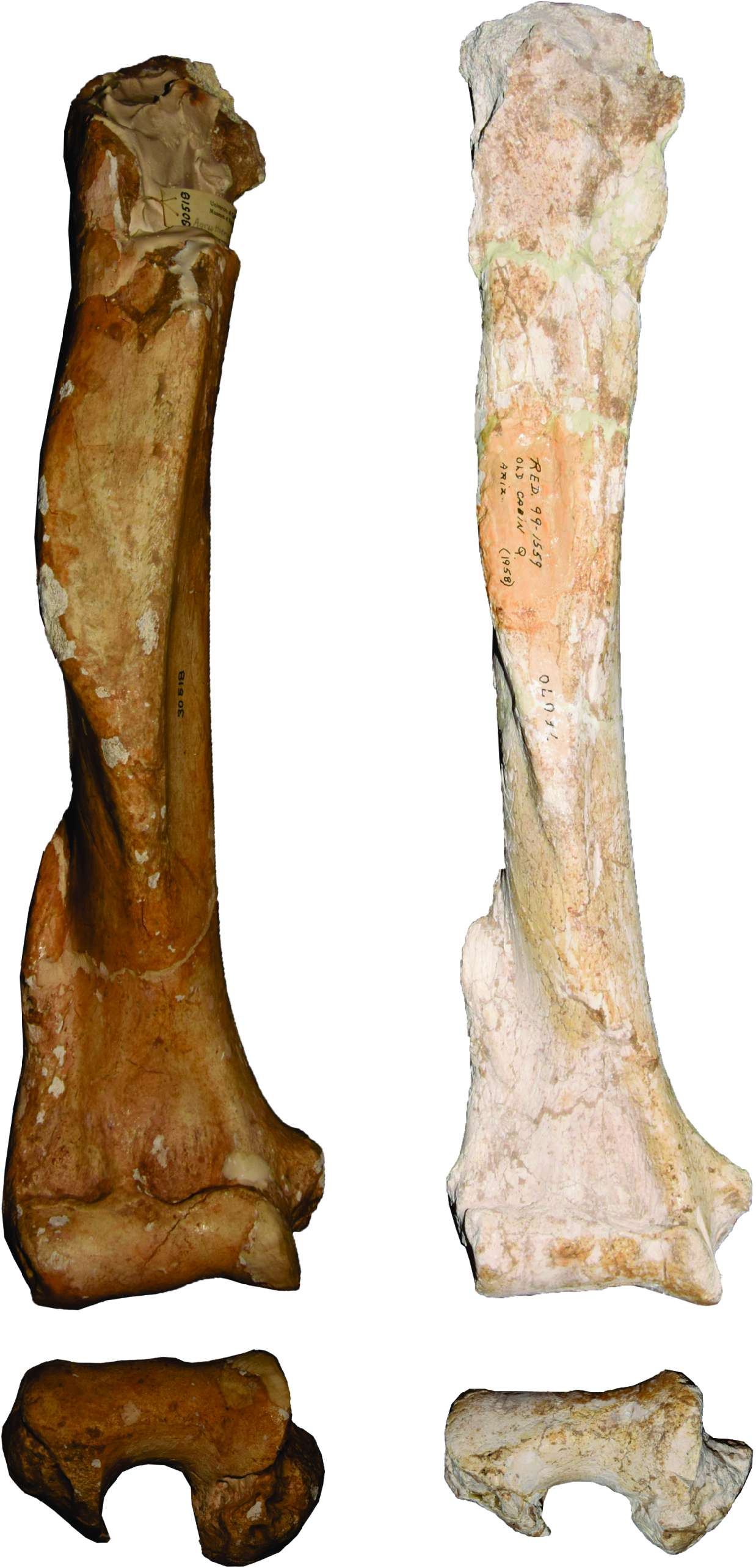
HUMERUS: Five nearly complete humeri are present. The best preserved is UCMP 31818 from Coffee Ranch, Texas, with a nearly undeformed distal part but crushed proximal portion ( fig. 12 View FIG ). The deltoid crest is well developed and extends distally along 2/3 the length of the humerus. The distal part of the humerus is weakly expanded. The trochlea is weakly undulating on its distal border, whereas there are two concavities on the proximal border. The epicondyles are very weak for an ursoid. The lateral border of the ectepicondyle ridge is slightly folded medially. The entepicondylar foramen is absent. In distal view, both the entepicondyle and ectepyle are well developed and oriented perpendicularly to the trochlea, forming the lateral wall of the olecranon fossa. AMNH F:AM 76070 also is well preserved, with moderate dorsoventral compression. Its morphology is generally similar to that of UCMP 31818, although the lateral border of its ectepicondyle ridge is distinctly medially folded. Other humeri in this species have similar morphologies. None of the humeri preserve an undeformed proximal end, but it is probably anteroposteriorly elongated, judging from the corresponding facet of the scapula.
RADIUS: The only undeformed radius of this species, UNSM 76013, comes from the SM-101 locality, Sherman County, Nebraska ( fig. 13 View FIG ). This bone is slightly curved. The proximal facet for the humerus is oval in shape, with a slightly concave anterior border. A proximal tuberosity is present near the proximal facet, and the distal facet for articulation with the scapholunar is robust. There are three muscle tendon grooves on the distal end of the radius ( Evans and de Lahunta, 2013). The groove for the extensor digitalis communis is the deepest, as in Indarctos , but distinct from Ursus and Ailuropoda in which this groove is wide and shallow. All other radii are either broken or compressed, but their general morphology does not differ from that of UNSM 76013.
ULNA: The best-preserved ulna, AMNH F:AM 50042, comes from the Guymon Formation ( fig. 14 View FIG ). The shaft is slightly crushed, but the proximal and distal ends are largely undeformed. The olecranon process is moderately developed, with a strongly medially expanded process; its anterior boundary with the anconeal process is unclear. The coronoid process of the ulna is well developed and wide. A fossa distal to the coronoid process is present but shallow. Other ulnae, though crushed or incomplete, show similar morphologies.
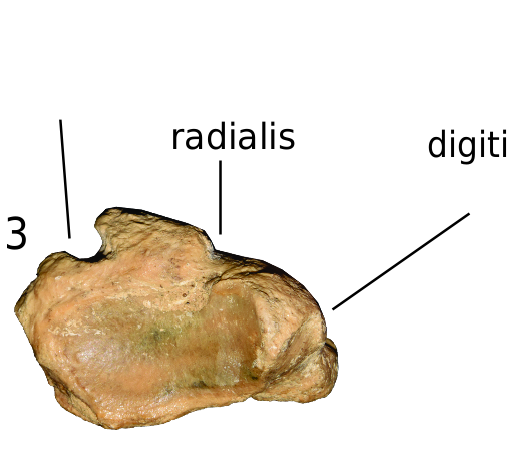
CARPAL BONES: Carpal bones of this species are rare. Only the scapholunar, cuneiform, and pisiform are represented. AMNH F:AM 146450 is the best-preserved scapholunar, though communis carpi abductor I longus A A 410 cm
A2 B C
10 cm
its lateral border is broken. The bone is massive, with a strongly developed ventromedial process; the distal end of this process bends laterally. In distal view, the articular facet for the trapezium and trapezoid is not ventrally extended. The only complete cuneiform is AMNH F:AM 146452 from Coffee Ranch. This bone has a rhomboid shape. The medial facet for the scapholunar is small, whereas the lateral facet for the pisiform is well developed. Two nearly undeformed pisiforms are preserved. AMNH F:AM 112588 from Coffee Ranch is massive. The proximal facets for articulation with the cuneiform and ulna are asymmetric in size. AMNH Red. (Redington) 117-1831 from the Quiburis Formation is smaller than the Coffee Ranch specimens, but similar in morphology.
FEMUR: All femurs are heavily crushed. Nevertheless, some characters can be observed ( fig. 15 View FIG ). The femur is generally short and the greater trochanter is well developed. The femoral head is slightly higher than the greater trochanter, as also can be seen in the undeformed femur figured by Harrison (1983). The greater trochanter of AMNH F:AM 146503 is similar in height to the head of the femur, reminiscent of the condition in felids, although deformation may
A B C
cm
10
account for its unusual morphology relative to other specimens. The only large felid from the Quiburis Formation is Amphimachairodus coloradensis , but it does not seem possible for this species to have a femur that reached such an extremely large size ( 475 mm), so it is unlikely that this bone represents a felid. Its shaft is elongated. The distal articular facets are heavily crushed, but the medial border is clearly substantially higher than the lateral border. The undeformed femur (L= 412.4 mm) from the Edson Local Fauna ( Harrison, 1983) is distinctly smaller than those from the Quiburis Formation (L= 454.12–516.28 mm), suggesting possible intraspecific geographic variation.
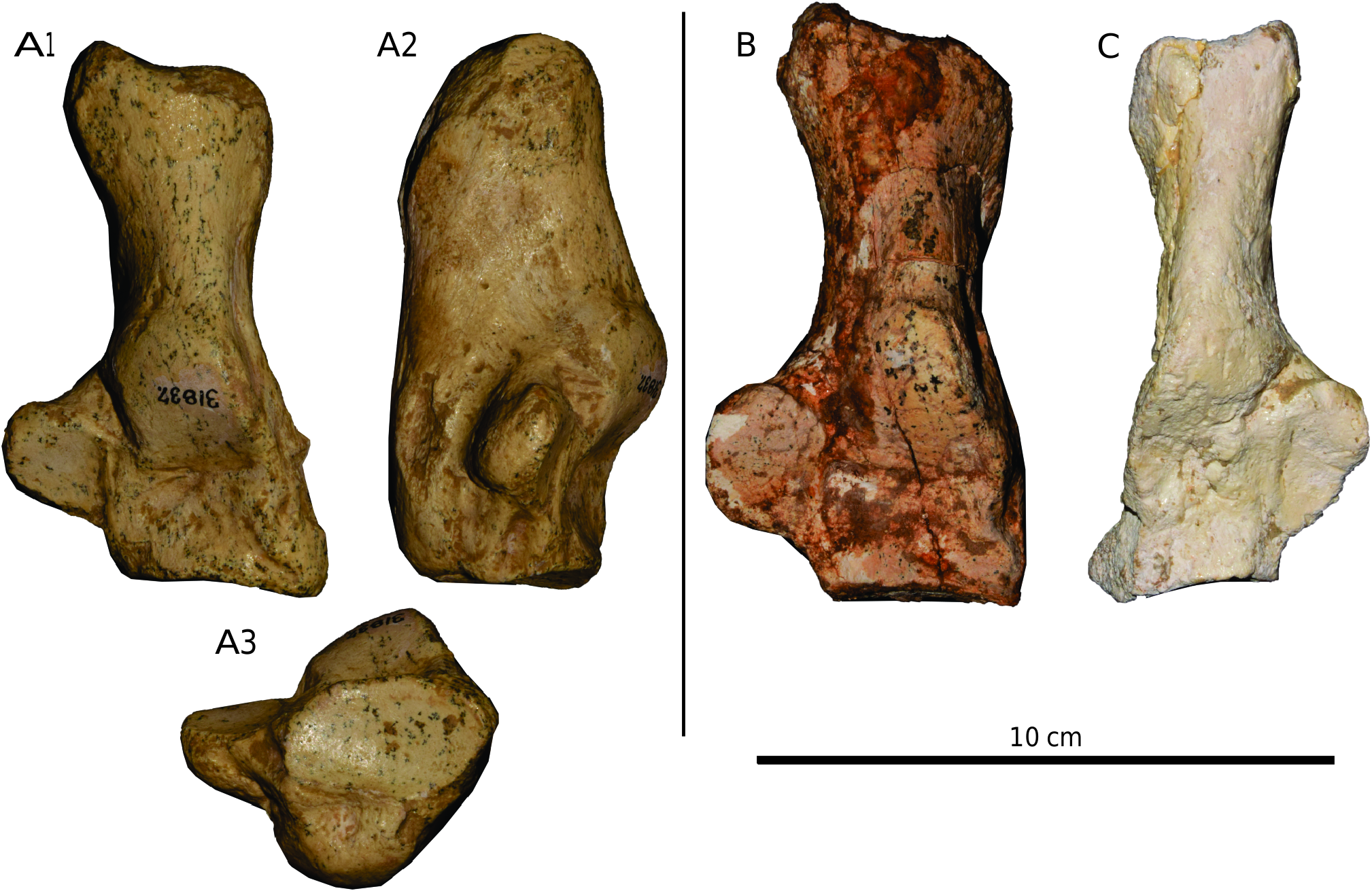
TIBIA AND FIBULA: All tibias are heavily crushed and few distinct characters can be seen. The tibial tuberosity seems to be moderately developed. Only a broken fibula attached to a tibia (AMNH F:AM 146513) is known; on it, the distal fibular facet for the astragalus is well developed.
ASTRAGALUS: Several astragali are preserved, but all are crushed or broken. The neck is moderately long. The facet for the cuboid is present, but seems to be small.
CALCANEUM: The best-preserved calcaneum UCMP 31837, which is undeformed, is from Coffee Ranch ( fig. 16 View FIG ). This calcaneum is massive and deep. Its sustentacular process is relatively small, and its distal border is widely separated from the distal articular facet. The distal facet for the cuboid is anteroposteriorly narrow. Other calcanea are consistent in these characters.
MESOCUNEIFORM: Only one mesocuneiform, from Coffee Ranch (AMNH FM 146707), is preserved. Both the proximal and distal articular facets are triangular in shape. Its medial border is thicker than the lateral border, and the facets for the entocuneiform and ectocuneiform both are well developed.
CUBOID: The only undeformed cuboid, UCMP 30519, is from Coffee Ranch. The bone is relatively elongate. The proximal and distal facets form a small acute angle. The proximal facet for the calcaneum is transversely elongated. Two smaller facets lie medial to the calcaneal facet. The dorsal one is larger, representing the facet for the astragalus, and the ventral one is smaller, representing the facet for the navicular. The medial facet for the ectocuneiform is small and less distinct than the other cuboid facets. The distal facet for MT4 and MT5 is triangular in shape, and the part articulating with MT5 is rather small. The ventral process of the cuboid is massive, as in all bears.
METAPODIALS (METACARPALS/METATARSALS): The metapodial bones are slender, and are similar in general morphology to other bears. The medial process of MC I for the M. abductor pollicis longus is relatively weak; the medial scar for the M. extensor carpi radialis longus does not form a process; the rugous scar for the carpal ligament of MC IV is well developed, extending more than halfway along the bone shaft on its ventral surface; and MC/MT IV are longer than MC/MT V.



No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Huracan coffeyi ( Dalquest, 1986 )
| Jiangzuo, Qigao, Flynn, John J., Wang, Shiqi, Hou, Sukuan & Deng, Tao 2023 |
Agriotherium schneideri
| Hunt, R. M. J. 1998: 184 |
Agriotherium sp.
| Harrison, J. A. 1983: 22 |
Agriotherium schneideri : Savage, 1941: 696
| Savage, D. E. 1941: 696 |