
Louisea edeaensis ( Bott, 1969 )
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4231.2.9 |
|
publication LSID |
lsid:zoobank.org:pub:C4C90F39-AB2A-497B-BE31-62AF69899D6D |
|
DOI |
https://doi.org/10.5281/zenodo.6015440 |
|
persistent identifier |
https://treatment.plazi.org/id/403AB05A-FFD2-9F2F-07A7-F9CCFA7C3468 |
|
treatment provided by |
Plazi |
|
scientific name |
Louisea edeaensis ( Bott, 1969 ) |
| status |
|
Louisea edeaensis ( Bott, 1969) View in CoL
( Figs. 1–3 View FIGURE 1 View FIGURE 2 View FIGURE 3 )
Globonautes macropus edeaensis Bott, 1969: 360 View in CoL ;1970: 24, pl. 1, fig. 3–5, pl. 26, fig. 8; Cumberlidge 1987: 2215, table 2. Louisea View in CoL edeaensis— Cumberlidge 1994: 124 –125, fig. 1, table 1; 1997: 577; 1999: 227, 5300, 54–57, figs 46F, 47E, 48E, 49F, 51F, 52F, 62F, 68F, table IX, plate 3; Ng et al. 2008: 168; Cumberlidge et al. 2009: 6.
Material examined. CAMEROON: 1 adult ♂ (CW 17.5, CL 13.3, CH 6.8, FW 6.8) ( ZMB Crust. 26930), Lake Ossa, Bedimet Island (03°48.959' N; 010°03.309' E), 90 m asl, 10 Jul. 2015, coll. P. A. Mvogo Ndongo GoogleMaps ; 6 subadult ♂s (CWs 15.0, 15.0, 14.1, 14.6, 12.5, 12.0) (LZUY 15-1), 4 adult ♀s (CWs 19.9, 17.4, 16.8, 16.7) (LZUY 15-2); 6 subadult ♀s (CWs 15.5, 14.9, 14.8, 13.8, 13.0, 12.5) (LZUY 15-3), same collection data as ZMB Crust. 26930; 3 juv. (CWs 11.0, 11.0, 5.6) ( LZUY 15-4 ), same locality, 15 Jan. 2016, coll. P. A. Mvogo Ndongo GoogleMaps ; 1 adult ♂ (CW 19.3), 2 adult ♀ s (CWs 19.4, 17.5) ( NHMW 1877 View Materials ), Yaounde, 1907, coll. Haberer. The differences between juveniles, subadults, and adults of the collected specimens of Louisea edeaensis observed from Bedimet Island, are discussed below.
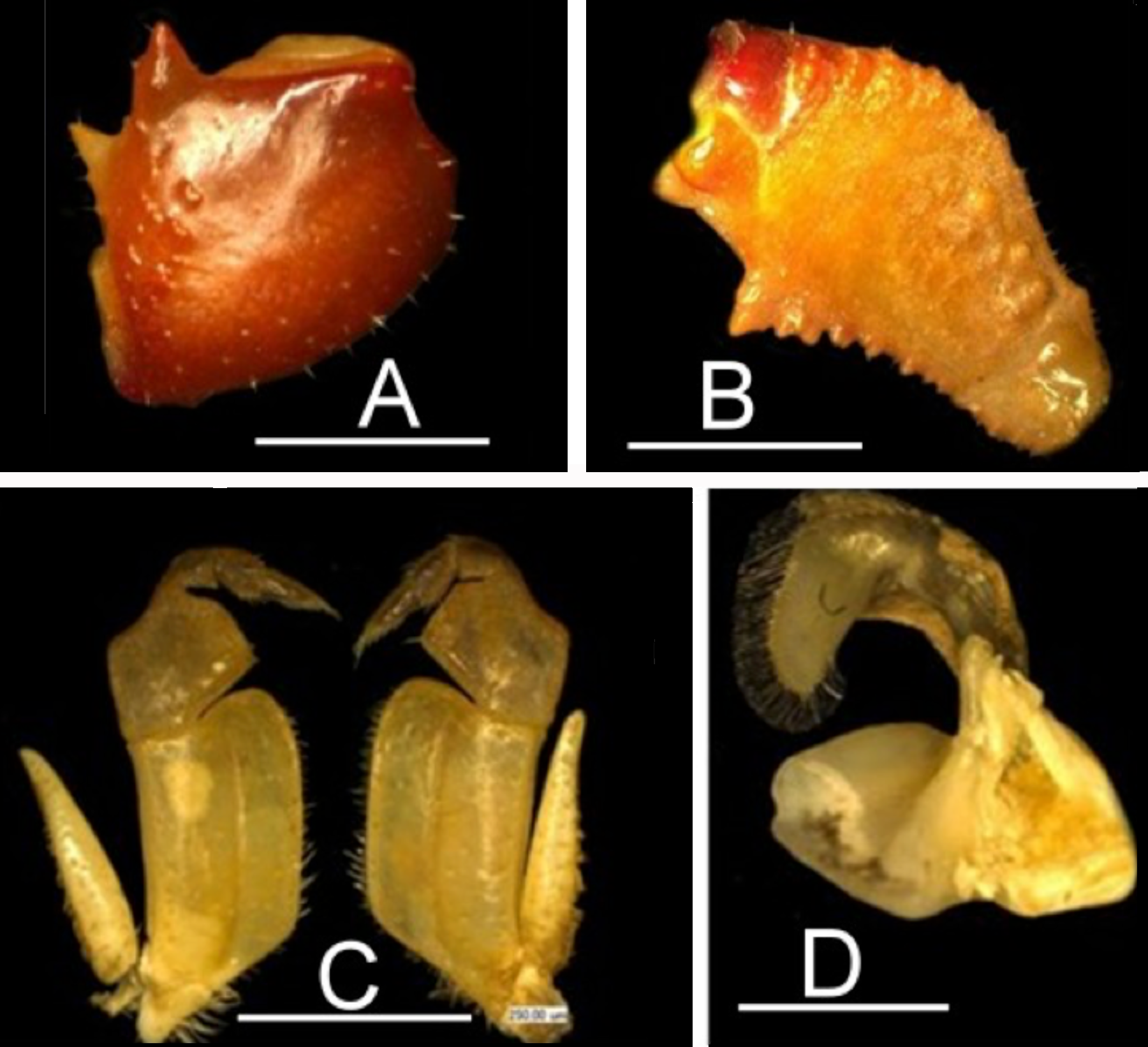
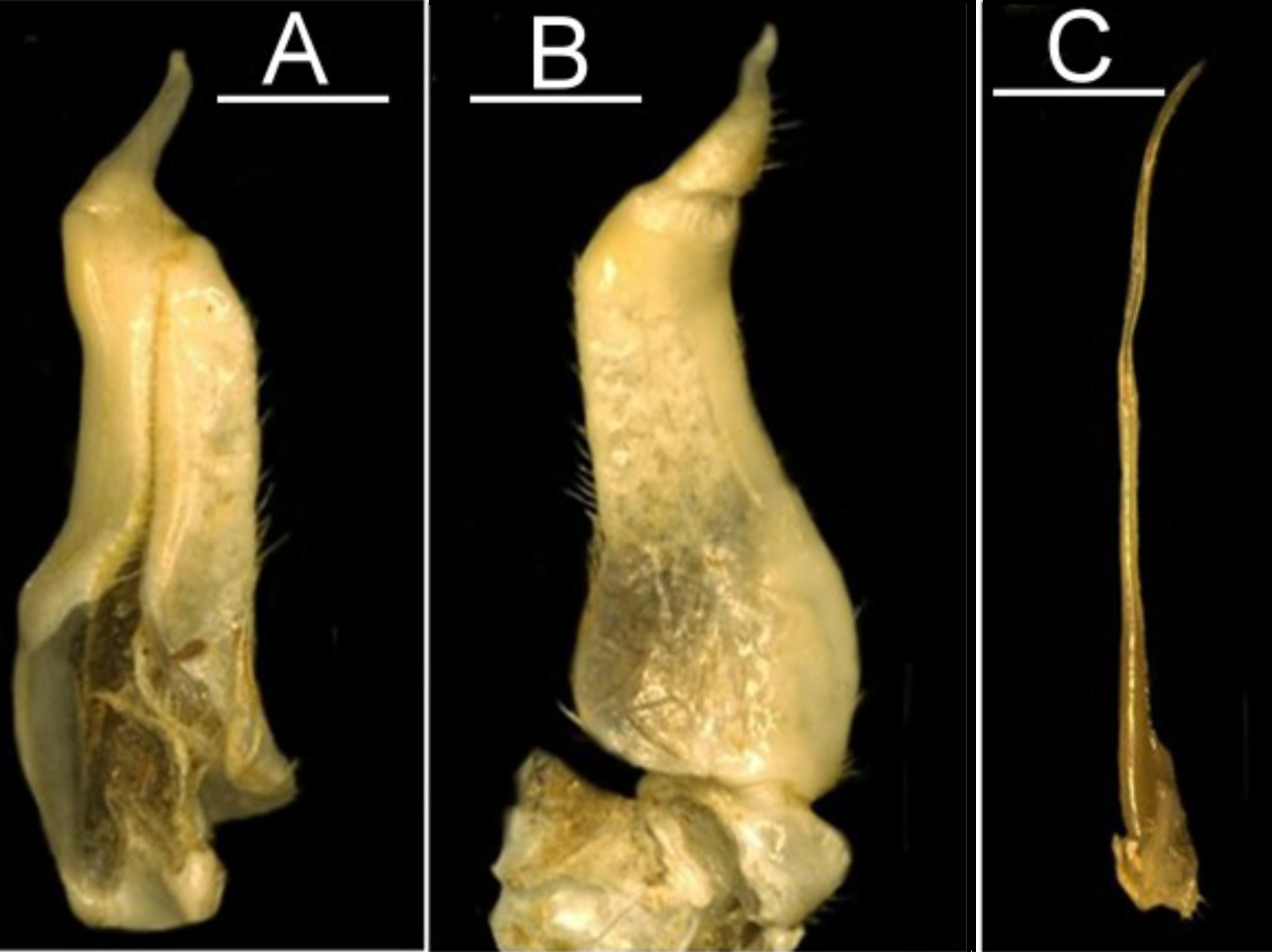
Diagnosis. Carapace moderately high ( CH /FW 1.0–1.1), smooth, postfrontal crest faint, complete, meeting epibranchial teeth; exorbital tooth small, low, epibranchial tooth small, intermediate tooth between exorbital, epibranchial teeth small but distinct ( Fig. 2A View FIGURE 2 ); third maxilliped exopod completely lacking flagellum, third maxilliped ischium with vertical groove ( Fig. 1 View FIGURE 1 C); mandibular palp with small lobe at junction between terminal, subterminal segments ( Fig. 1 View FIGURE 1 D); medial inferior margin of cheliped merus with large jagged distal tooth, followed by row of small pointed teeth along rest of meral margin ( Fig. 1 View FIGURE 1 B), first carpal tooth on cheliped carpus large, broad, bigger than second carpal tooth ( Fig. 1 View FIGURE 1 A); G1 terminal article outwardly-directed, base broad, tapering distally, ending in blunt hose-like tip ( Fig. 3 View FIGURE 3 A, B); G2 terminal article long, flagellum-like ( Fig. 3 View FIGURE 3 C); small species, mature at CW 16–17 mm.
Description. For a full description see Cumberlidge (1994, 1999). Character state descriptions that have been modified in the light of the new material are discussed below under Remarks.
Size. Small species, adult size range between CW 16.5–22.5 mm, pubertal moult occurring between CW 16– 17 mm, only known ovigerous female CW 19.9 mm.
Growth and size classes. Ontogenetic development in our population sample of L. edeaensis from Lake Ossa is discussed here in terms of three size categories (juveniles, subadults, and adults) following the criteria proposed by Cumberlidge (1999). The natural population of L. edeaensis includes male and female specimens in a range of body sizes that are grouped here into: (1) juveniles (small-bodied non-reproductive specimens), (2) sub-adults (medium-sized but non reproductive specimens), and (3) adults (large sexually mature adults). Juveniles of both sexes are characterized by their small size (CW up to 11 mm) and by the morphological similarity between male and female specimens; both sexes have a similar-shaped abdomen and equal-sized chelipeds, and the gonopods (males) and pleopods (females) are short and undeveloped. The abdomen of juveniles of both sexes is a slim elongated triangle that tapers to a rounded telson and covers only the subabdominal cavity. The G1 and G2 of juvenile males are both short, straight, and similar in size, whereas juvenile females possess four pairs of small, narrow pleopods. Subadult crabs (CW up to 15.5 mm) are characterized by their partial development towards the adult form as a result of allometric (accelerated) growth of the abdomen and pleopods (females) or of the major cheliped and gonopods (males). The abdomen of subadult female crabs is broad and wide but only partially covers the thoracic sternum and is distinctly broader than the slim triangular abdomen of juvenile females and all size classes of male crabs. Although the gonopods of subadult males are long enough to reach sternal segment s6 they are still undeveloped (mostly straight with only a slight outward curve). The abdomen of the only sexually mature adult male (CW 17.5 mm) from Lake Ossa is slim and triangular covering the narrow subabdominal cavity, whereas the abdomen of adult females is conspicuously wide and covers most of the thoracic sternum and overlaps the coxae of the walking legs (p2–p4). The subabdominal cavity of adult males is deep and narrow and contains the G1 and G2, whereas that of adult females is broad, shallow, and includes four sets of well-developed plumose pleopods for attachment of egg masses. The major cheliped of adult males is distinctly larger than the minor cheliped, and the dactylus is arched so that a distinct space remains between the closed fingers. The chelipeds of females grow isometrically and are not dramatically different from each other in any of the three size classes and one cheliped is not conspicuously enlarged with the respect to the other (as is the case for adult males). The G1 of the adult male is long enough to reach sternal segment s4 (about one third the length of the G1 subterminal segment) ( Figs. 3 View FIGURE 3 A, B), and the terminal article is broad at the base and tapers sharply before curving outward to end in a broad tip, whereas the G2 subterminal segment is long and thin, and the terminal article is long and flagellum-like ( Fig. 3 View FIGURE 3 C).
The morphological differences shown by the three size classes in L. edeaensis can be understood in terms of the isometric (proportional) growth and allometric (accelerated) growth shown by some of the important taxonomic characters. All characters in juveniles show only isometric growth, whereas subadult and adult males show different degrees of allometric growth (accelerated development) in the major cheliped and gonopods but not in their abdomen, while subadult females show allometric growth in their pleopods and abdomens (but not in their chelipeds). The boundary between juveniles (with all characters proportional) and subadults is detected when specimens show accelerated growth of their abdomen and pleopods (females) or of their chelipeds and gonopods (males). The boundary between subadults and adults is the pubertal moult from non-reproductive to reproductive animals. This moult is most clearly recognized in females who are considered to be adults when their allometrically growing abdomen has increased to the point where its edges cover the coxae of the walking legs (and their allometrically growing pleopods are long, broad, and feathery, and capable of supporting eggs and or hatchlings). Male and female freshwater crabs go through the pubertal moult at the same size, so the carapace width that signals that a female specimen is an adult can be used to establish whether a male specimen is an adult (but in the absence of an adult female, a male with a conspicuously enlarged major cheliped and well-developed gonopods can be judged to be adult with confidence). The differences between subadult (non-reproductive) and adult (reproductive) freshwater crabs is that although subadult males have enlarged gonopods and a major cheliped in comparison with juveniles, these characters are still only partially developed and have not yet reached the final form seen in adults.
Color. Dorsal carapace ( Fig. 2A View FIGURE 2 ), postfrontal crest, third maxillipeds, chelipeds, walking legs of living adult specimens of L. edeaensis are all red brown, whereas the lateral carapace and sternum are pale brown to creamyellow.
Distribution. Louisea edeaensis is endemic to the rainforest zone of southern Cameroon, where it is now known from an expanded range that includes four localities in the Sanaga (Edea and Lake Ossa), Mfoundi (Yaounde), and Wouri (Yabassi) river basins. Louisea edeaensis might be expected to occur in other suitable wetland habitats in these river drainages, but this part of Africa is still poorly surveyed for freshwater crabs.
Remarks. The new series of specimens of L. edeaensis from Bedimet Island in Lake Ossa agree well with the diagnostic character states seen in the holotype from Edea, and with the specimens from Yaounde and Yabassi ( Cumberlidge 1994, 1999). All have a moderately high carapace ( CH /FW 1.0–1.1), a smooth carapace surface; a small and short exorbital tooth; a small epibranchial tooth; one distinct intermediate tooth between the exorbital and epibranchial teeth; a large jagged distal tooth on the medial inferior margin of the cheliped merus; a row of small pointed teeth along the rest of the meral margin; an outwardly-directed G1 terminal article that is broad at the base, tapers sharply and ends in a broad tip; long and flagellum-like G2 terminal article. All specimens are relatively small in size, not exceeding 20 mm CW.
The new specimens of L. edeaensis allow a more detailed examination of several other important taxonomic characters that have been described differently by Cumberlidge (1994, 1999). The mandibular palp of the specimens from Lake Ossa and the specimen from Yabassi (“ Cumberlidge 1994: fig. 3d”) have a small but distinct lobe at the junction between the terminal and subterminal segments (“ Cumberlidge 1994: fig. 2F”) which is similar to the holotype, although it was described by Cumberlidge (1994) as ‘a rudimentary flange at junction between segments’. The third maxilliped ischium of the specimens from Lake Ossa has a distinct vertical groove ( Fig. 1 View FIGURE 1 C), which is clearer than that of the holotype and the specimens from Yabassi which have a faint or absent vertical groove (“ Cumberlidge, 1994: figs. 1c, 3c”). The holotype of L. edeaensis (“ Cumberlidge, 1994: fig. 1a, b”) from Edea and the specimens from Lake Ossa both have a faint postfrontal crest ( Fig. 2A View FIGURE 2 ) that on close examination extends as far as the epibranchial teeth, which is a character state that agrees with that of the Yabassi specimens, despite the fact that those specimens were illustrated with a complete and distinct postfrontal crest by “ Cumberlidge (1994: fig. 2a, b)”.
The holotype of L. edeaensis from Edea (“ Cumberlidge 1994: fig. 1a, b”) and the specimens from Yabassi (“ Cumberlidge 1994: fig. 2a, b”) and Lake Ossa ( Fig. 2A View FIGURE 2 ) differ from each other in a few respects. The first carpal tooth of the cheliped of the holotype is smaller than the second carpal tooth (“ Cumberlidge 1994: fig. 1a”), whereas in the specimens from Yabassi (“ Cumberlidge 1994: fig. 3f”) and in the specimens from Lake Ossa, the tooth is large, triangular, and larger than the second carpal tooth ( Fig. 1 View FIGURE 1 A). It is possible that the holotype is atypical in this respect and that its first carpal tooth has been damaged making it appear smaller than it actually is. Furthermore, the G1 of both the holotype of L. edeaensis from Edea (“ Cumberlidge 1994: fig. 1h”) and the specimens from Lake Ossa have a broad rectangular dorsal membrane at the junction between the subterminal segment and the terminal article ( Fig. 3 View FIGURE 3 A, B), but this membrane is not shown at all in the G1 of the specimen from Yabassi illustrated by “ Cumberlidge (1994: fig. 3i) ”, which may well be an artwork error.
The present work brings into question the diagnostic characters used by Cumberlidge (1999) to distinguish between L. edeaensis and L. balssi (such as differences in the relative size of the first carpal tooth, and a distinct and complete vs. faint and incomplete postfrontal crest) and creates the need to update the key to the species of Louisea provided by Cumberlidge (1999). These latter two morphological differences led Cumberlidge (1999) to tentatively include the specimens from Yabassi under L. balssi , but it now appears that in the light of new evidence the specimens from Yabassi properly belong to L. edeaensis , an opinion which is in agreement with Cumberlidge (1994). In summary, the postfrontal crest of L. edeaensis is faint and completely crosses the carapace, and the first carpal tooth of the cheliped is larger than the second carpal tooth. Despite this, we are unable to discuss in detail the taxonomic status of L. balssi , due to a lack of new material and the fact that the only known specimens are juveniles collected in 1907, which are all in poor condition and no information on the gonopod is available. Comparisons of L. edeaensis and L. balssi must therefore await future collection of more specimens of L. balssi that will hopefully include an adult male specimen suitable for morphological studies, and that will also provide fresh tissue for the inclusion of this rare and enigmatic taxon in molecular phylogenetic studies.
Habitat. The Lake Ossa complex comprises nine lakes and over twenty islands (including Bedimet Island (550 ha), where L. edeaensis was collected) and is part of the Lake Ossa Wildlife Reserve, a protected area managed for mammals and birds, but not for freshwater invertebrates. Lake Ossa lies in the tropical rain forest zone in the wettest region of Southwest Cameroon and receives more than 3,000 mm of rainfall annually ( Wirrmann 1992; Reynaud-Farrera et al. 1996). The extensive aquatic habitats in the Lake Ossa complex are drained by numerous streams that flow via a series of lakes into the Sanaga River ( Reynaud-Farrera et al. 1996). Although no month is consistently devoid of rain, a dry season (December to March) and a rainy season (April to September) are clearly distinguishable ( Olivry 1986). Air temperatures show no great seasonal variation with average monthly maxima between 27°C and 35°C, and average monthly minima between 21°C and 25°C ( Olivry1986).
Louisea edeaensis appears to prefer temporary water bodies such as puddles near small permanent streams as well as the damp conditions under fallen leaves on land adjacent to streams, and it is noteworthy that no specimens were found in nearby streams with a year-round flow. A number of threats to the continued survival of L. edeaensis were identified during this study including those that impact its habitat. Many parts of Bedimet Island (including the locality where this species was collected) have been cleared of natural forest cover to make way for agriculture with the result that the vegetation is now dominated by shade-tolerant tropical plants of the family Amaranthaceae that grow well in moist soils, cultivated fields, and wastelands. Apart from the substantial disturbance of the original vegetation on Bedimet Island, pesticides applied to the crops may drain into the aquatic habitats inhabited by L. edeaensis , which could well have a serious impact on this species. Furthermore, the clearing of original forest cover has exposed the aquatic systems on Bedimet Island (and the freshwater invertebrates that live there) to direct sunlight, and this is reflected in the high average daytime water temperatures of up to 30°C recorded during this study (compared to average water temperatures in streams shaded by forest cover of between 22–26°C). In addition, our water chemistry data indicate that the marginal aquatic habitats on Bedimet Island have a very low average pH of 4.5, indicating seriously acidic conditions at these localities well outside of the preferred pH range (6.5–9.0) for most freshwater organisms ( Robertson-Brayan 2004). This is of concern because acidic conditions may influence hatching and survival of L. edeaensis and other aquatic invertebrates whose larval and/or juvenile stages are often more sensitive to low pH than are the adults. The immediate threats from habitat disturbance, altered water chemistry, and pollution all clearly raise questions about the long-term existence of L. edeaensis at this specific locality.
Conservation status. The extinction risk status of L. edeaensis was assessed in 2008 ( Cumberlidge, 2008a) as EN B1ab (i,ii,iii,iv,v) at a time when this species was known only from just two localities and had not been collected for over 100 years ( IUCN 2003, Cumberlidge 2011; Cumberlidge et al. 2009). Specifically, this extinction risk assessment meant that L. edeaensis was known from less than five locations, had an extent of occurrence (EOO) of less than 5000 km 2, and that its EOO, area of occupancy (AOO), and the number of locations were in continuing decline, as were the quality of its habitat and the number of mature individuals ( Cumberlidge 2008a). The present study adds two new locations for L. edeaensis (one from 1908/1909 and one from 2015) and these four locations allow the calculation of a revised geographic range estimated by using the EOO (the area contained within the minimum convex polygon around all sites of present occurrence) and the AOO (the area within the EOO that is actually occupied by the taxon). The EOO and AOO measurements from point locality data using GeoCAT (http://geocat.kew.org; Bachman et al. 2011) give a revised EOO of 6, 780 km 2 and an AOO of 16 km 2 (estimated by overlaying a 2 × 2 km grid and summing the area of occupied cells). If considered alone, this EOO would assign this species to either the vulnerable (VU) or EN categories. Given the new knowledge of its specific habitat requirements and the existence of immediate threats to its habitat, however, this species is more likely to be upgraded to critical (CR) once a formal IUCN Red List extinction risk assessment has been carried out. The newly-reported threats to an already endangered species area cause for concern and they underline the need for further studies of this species. This concern is especially important, because recent systematic attempts to collect specimens of Louisea from the type locality at Edea (last collected in 1910) and from Yabassi (last collected in 1907) during this campaign were unsuccessful. There is clearly a need for further field research aimed at gathering data needed to develop a conservation action plan in order to save this rare and endangered species from extinction.

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Louisea edeaensis ( Bott, 1969 )
| Mvogo Ndongo, Pierre A., Rintelen, Thomas Von, Schubart, Christoph D., Albrecht, Christian, Tamesse, Joseph L. & Cumberlidge, Neil 2017 |
Globonautes macropus edeaensis
| Cumberlidge 2009: 6 |
| Cumberlidge 1994: 124 |
| Cumberlidge 1987: 2215 |
| Bott 1969: 360 |