
Eolestes ramosus, Greenwalt, Dale E. & Bechly, Günter, 2014
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3887.2.2 |
|
publication LSID |
lsid:zoobank.org:pub:D9275063-BD74-498E-B504-1421B50114A5 |
|
DOI |
https://doi.org/10.5281/zenodo.6141526 |
|
persistent identifier |
https://treatment.plazi.org/id/40446A46-FFBF-670A-FF4C-060A8D52FD81 |
|
treatment provided by |
Plazi |
|
scientific name |
Eolestes ramosus |
| status |
sp. nov. |
Eolestes ramosus n. sp.
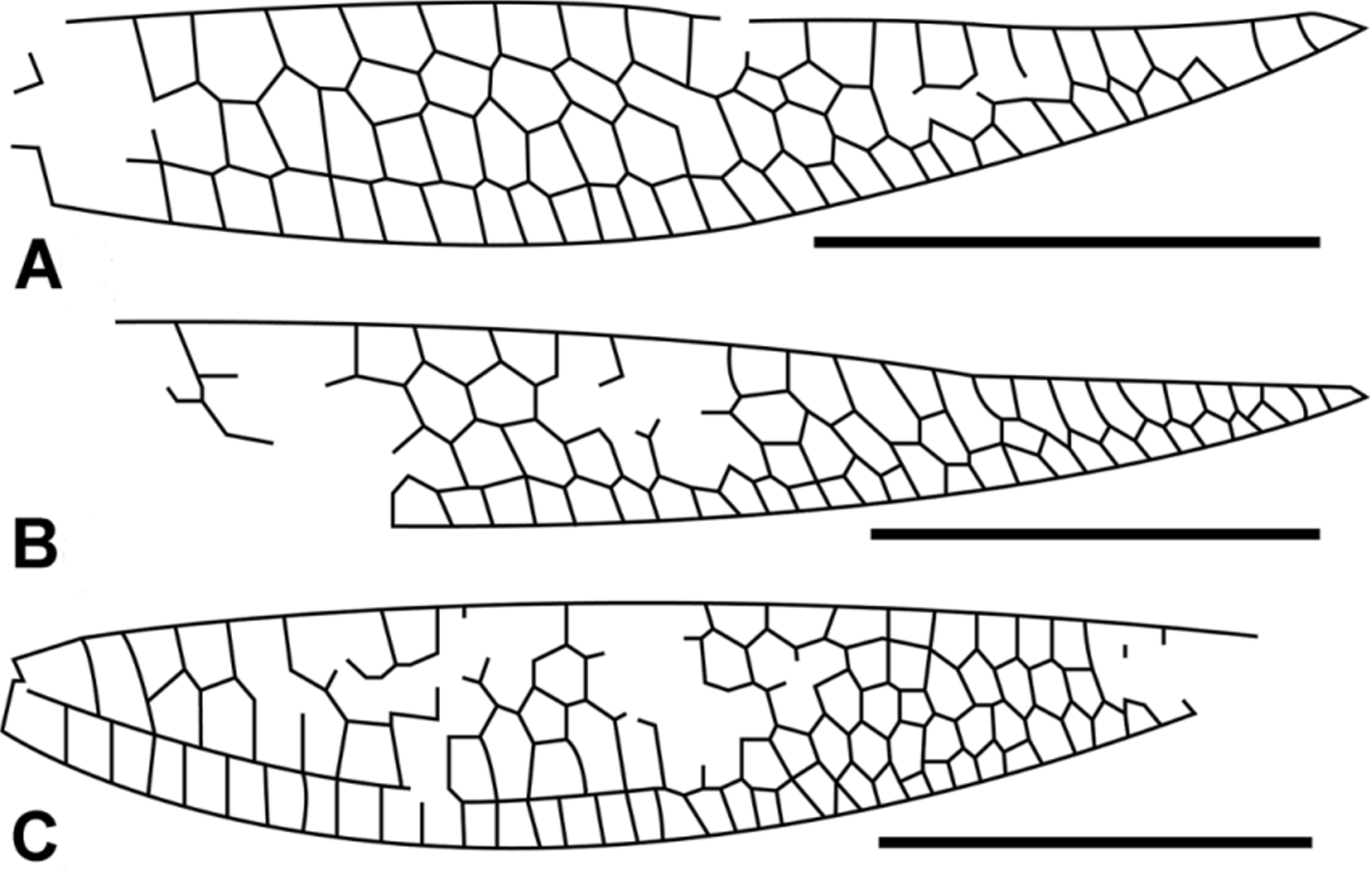
( Figures 5 View FIGURE 5 C, 6C, 7)
Holotype. USNM 559047, National Museum of Natural History, Washington, D.C.
Type locality and stratum. Disbrow Creek site, Middle Fork of the Flathead River, Pinnacle, Montana, USA. Coal Creek Member of the Kishenehn Formation, early Middle Eocene, 46.2±0.4 or 43.5±4.9 mya ( Constenius, 1996).
Etymology. Species name ramosus from the Latin word ramosus (branching), an indication of the branching of the supplementary longitudinal sectors in the MP-CuA field.
Diagnosis. Differs from E. syntheticus by the following characters: 1) CuP origin at instead of distal of the separation of AA’ and AA’’; 2) three supplementary longitudinal sectors in the cubital field vs. two; 3) 2–4 rows of cells between distal parts of IR2 and RP 3/4 vs. 1–2.
Description. An intact fossil damselfly wing, hyaline, 27.1 mm long and 6.06 mm and 7.13 mm wide at nodus and widest point respectively ( Fig. 5 View FIGURE 5 C); pterostigma four cells and 3.05 mm long, 0.61 mm wide; pterostigmal brace oblique; distance between wing apex and pterostigma, pterostigma and nodus, nodus and arculus and arculus and base 2.43, 12.13, 5.8 and 3.66 mm respectively; petiole length relative to distance from petiole to nodus = 0.44; post-pterostigmal cell area consists of two single cells proximally, three smaller cells distally and five double cells in between; two antenodal and two antesubnodal crossveins. Ax1 opposite separation of AA’ & AA’’ and separated from Ax2 by 1.30 mm; Ax2 opposite arcular crossvein just distal to RP origin and anterior arcular crossvein; RP separates from RP +MA just below RA+ RP so that RP +MA very short; supplementary antenodal crossveins absent; posterior arcular crossvein separating from discoidal cell 0.33 mm below RA; nodus and subnodal bracket not preserved, subnodal crossvein apparently oblique; 14 postnodal crossveins exactly aligned with postsubnodal crossveins with exception of last four; ‘‘lestine’’ oblique vein ‘‘O’’ present four cells distal of base of RP 2; base of IR1 four cells and 2.7 mm distal of RP 2 origin, IR1 zigzagged through first three cells, underlies a single row of cells that, proximal to pterostigma, are higher than they are wide and overlies a row of seven single cells that distally expand through six double cells and subsequently, an area of reticulated cells with two and three secondary longitudinal veins; base of RP 2 two cells and 1.46 mm distal of subnodus and six cells and 5.41 mm distal of origin of IR2; IR2 and PR3/4 originate much closer to arculus than nodus. IR2 arched abruptly toward RP at base with crossvein opposite, one cell and 0.86 mm from origin of RP 3/4; IR2 underlies a row of 18 single cells basally, followed by a gradual increase in number of secondary veins until there are 11 small cells at wing margin; RP 3/4 originates one cell and 1.30 mm from arculus and underlies a row of single cells that transitions from cells longer than high to higher than long before double cells appear just proximal to beginning of pterostigma; MA slightly zigzagged after a point about midway between arculus and nodus, and underlies a row of cells that transitions to double cells at level of IR1 origin and then gradually to 17 small cells at wing margin; MP underlies a single row of cells that gradually transitions from square-shaped to cells higher than wide; CuA a prominent vein that leaves subdiscoidal cell 0.29 mm below MP and transitions to a zigzagged pattern at about level of origin of RP 2; it underlies a broad cubital field 2.20 mm in height that starts with a single cell below base of MP and quickly transitions through one, two and then three secondary longitudinal veins and then back to two secondary veins at wing margin ( Fig. 6 View FIGURE 6 C); cubito-anal field consists of a single row of cells (12 distal of subnodus) that, except for the first, are higher than wide, and terminates at a point halfway between nodus and pterostigma; cubito-anal field 2.85 mm in height; CuP origin at separation of AA’ & AA’’ ( Fig. 6 View FIGURE 6 ), directly below Ax1; petiole well defined, 2.51 mm long (from base to separation of AA’ and AA’’) and 9.3% of wing length; discoidal cell closed basally, 1.50 mm long (end of posterior arcular vein to origin of MP) and 0.73 mm wide (origin of arcular vein to origin of MAb) with an acute posterior internal angle of 27 degrees (Note: given the curved nature of the posterior portions of MAb and MP & CuA, the angle is that between lines that align with the majority of these two veins.); distal side of discoidal cell (MAb) nearly perpendicular to RA (7 degrees from vertical relative to RA), and 1.11 mm in length; anterior, posterior and basal (posterior arculus) sides 0.52 mm, 1.45 mm and 0.39 mm long; Ratio of lengths of anterior and posterior sides of discoidal cell = 0.36; subdiscoidal cell elongate with no fusion of CuP & AA’ to posterior wing margin distal of CuP; MP only partially preserved but with apparently no or very little arch as it leaves discoidal cell at angles of 93 and 76 degrees from MAb and vertical respectively.

| USNM |
Smithsonian Institution, National Museum of Natural History |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.