
Titanethes albus matovici, Karaman & Horvatović, 2018
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4459.2.3 |
|
publication LSID |
lsid:zoobank.org:pub:49F5AF2B-E795-404B-A946-2C00DB6011B2 |
|
DOI |
https://doi.org/10.5281/zenodo.5983534 |
|
persistent identifier |
https://treatment.plazi.org/id/40481448-6752-FF8B-FF28-FBEA25FAF915 |
|
treatment provided by |
Plazi |
|
scientific name |
Titanethes albus matovici |
| status |
subsp. nov. |
Titanethes albus matovici View in CoL n. ssp.
Figs 38–56 View FIGURES 38–41 View FIGURES 42–45 View FIGURES 46–50 View FIGURES 51–53 View FIGURES 54–56 , 72–75 View FIGURES 72–75
Titanethes biseriatus: Strouhal, 1939a View in CoL
Material examined: HOLOTYPE: male 8.4 mm, Bosnia and Herzegovina, Republika Srpska, Foča, Miljevina , Izbišno, Peruc cave, 43° 31.946'N, 18° 36.356'E, 18.1.2014, leg. I. Karaman & M. Horvatović, (ZZDBE- IZO1123) GoogleMaps . PARATYPES: ibid., 18.1.2014, leg. I. Karaman & M. Horvatović, 5♂, 12♀ GoogleMaps ; Foča, Miljevina, Izbišno, Peruc cave, 10.2013, leg. O. Matović, 2♀; Foča, Miljevina, Mala kutlača cave, 4.8.2014, leg. I. Karaman, 1♀; Foča, Miljevina Kuk cave, 7.8.2014, leg. I. Karaman, 1♂; Foča, Miljevina, Poljice, Pećina u Kanjonu Bistrice cave, 5.10.2016, leg. I. Karaman, 1♂ ; Banja Luka, Ljubačevo, quarry, Pećina Ljubačevo cave, 6.2.2016, Leg. I. Napotnik, 1♂, 3♀ ; Bosnia and Herzegovina, Sanski Most , Hrustovo, Hrustovača cave, 27.3.2012, leg. I. Njunjić, 1♂, 1♀, 3juv.
Etymology. The subspecies is named after Ognjen Matović, speleologist from Foča who first provided us with specimens and organized the first biospeleological investigations in the Foča region.
Diagnosis. The subspecies is characterized by the form of exopodites of male pleopods 1, with conspicuous mediobasal protrusion.
Description. Large troglobite animals (male up to 11 mm, female 12 mm long).
Cephalon and pereionites covered with sparsely distributed low tubercles ( Figs 38–40 View FIGURES 38–41 ).
Lateral lobes on cephalon are narrow, of conical shape ( Figs 38–39 View FIGURES 38–41 ).
Pleon wide and smooth; Pleonites 1–3 with short epimera and with traces of posterior row of tubercles; Male pleonite 4 with gland-piliferous organ in the form of small slightly svolen area with sparce scattered pores and lamina ( Figs 40–41 View FIGURES 38–41 ) or without it ( Fig. 42 View FIGURES 42–45 ). Male pleonite 5 medially with gland-piliferous surface ( Figs 40 View FIGURES 38–41 , 43– 45 View FIGURES 42–45 ). Porous surfaces are present dorsolaterally on pleonites and laterally on the basis of the uropods.
Telson with concave sides and truncated apex.
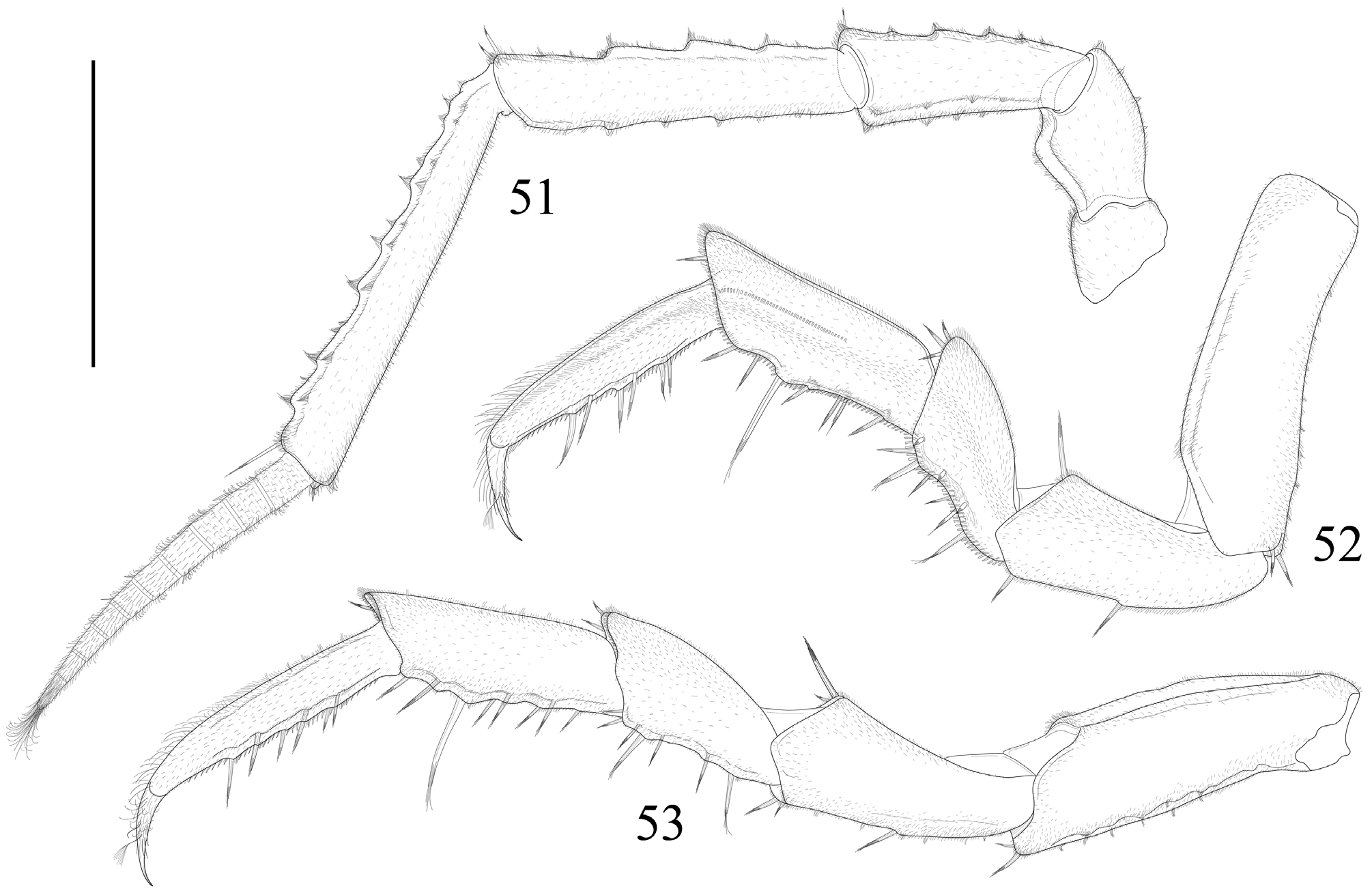
Antennula ( Fig. 46 View FIGURES 46–50 ) basal article 1.5 long as wide; second article as wide as long shorter than basal one; distal article as long as basal one, bearing eight aesthetascs. Antenna ( Fig. 51 View FIGURES 51–53 ) long; article 3 is three time as long as broad; article 4 as long as flagellum; article 5 longer than flagellum, twice as long article 3.
Right mandible incisiva with two penicils; pars incisiva with three teeth; lacinia mobilis massive, rim of terminal part with row of thorns; pars molaris oval with one penicil on proximal rim. Left mandible incisiva with thre penicils; pars incisiva lateralis with four teeth; pars incisiva medialis, with three teeth; pars molaris ovoid, with one penicil on proximal rim.
Maxilla 1 exopodite ( Fig. 50 View FIGURES 46–50 ) with 4 strong outer teeth, 5 medial teeth, 2 short and sharp inner teeth and two elongate pectinate seta distally. Endopodite ( Fig. 49 View FIGURES 46–50 ) with three penicils, longer than diameter of the endopodite proximal part. Maxilla 2 ( Fig. 48 View FIGURES 46–50 ) narrow, elongate with setose apical part, bilobate apex; inner lobe with 3 rod-like setae on its tip and a surface of very tiny elongated setae on its inner margin; outer lobe on its ventral side with a surface of dense and very tiny setae and row of rod-like setae terminally; outer lobe terminally on its dorsal side with row of elongated strong setae.
Maxilliped ( Fig. 47 View FIGURES 46–50 ) endopodite distolaterally with rounded tip; outer margin with rows of elongated tiny setae; palp on medial edge with a pronounced lobe with setae; endite terminally bearing stout penicil and 3 spines subterminally (one on medial edge, and two on outer edge).
Pereopods remarquable elongated, with elongated fused dactylus and unguis; fused dactylus and unguis less than a half the length of propodus. Pereopod 1 ( Fig. 52 View FIGURES 51–53 ) propodus, ischium and carpus of the same length; merus remarquable shorter than carpus. Pereopod 7 ( Fig. 53 View FIGURES 51–53 ) propodus longer than carpus; ischium as long as carpus; merus twice as long as wide.
Genital papilla ( Fig. 54 View FIGURES 54–56 ) gradually narrows along its whole length.
Pleopod 1 ( Fig. 55 View FIGURES 54–56 ) endopodite of two articles, distal shorter than basal one; basal article as long as exopodite; distal article rod like with bent tip. Exopodite with concave distal margin and conspicous triangular mediodistal protrusion bent outwards; mediobasal part strongly protruded too. Pleopod 2 ( Fig. 56 View FIGURES 54–56 ) endopodite of two articles, distal one three time as long as basal one. Exopodite trapezoidal, with straight distal margin.
Uropod endopodite distinctly shorter than exopodite, more proximally inserted.
Remarks. This subspecies was already known, described and illustrated in some detail by Strouhal (1939a) and later by Schmölzer (1965). It was actually mistakenly identified as Cyphonethes biseriatus (then Titanethes biseriatus ). Based on the details of the description and illustrated structure of male pleopod 1 exopodite ( Figs 57– 65 View FIGURES 57–65 ), we concluded that the specimens we collected and the samples that Strouhal (1939a) dealt with, belong to the same subspecies of the genus Titanethes .
Titanethes albus matovici n. ssp. is the second taxon in the genus Titanethes . Unlike the large nominal subspecies ( T. albus albus ), which is spread over a wide area on the slopes of the southeastern Alps and the westernmost part of the Dinarides (as expected as a subspecies complex), this taxon is somewhat smaller and disjunctly distributed in the central part of the Inner Dinarides ( Fig. 3 View FIGURE 3 ). Currently it is known from an area of the far north of Herzegovina, between the towns of Foča and Konjic and in Bosanska Krajina (Western Bosnia) south of the towns of Banja Luka and Sanski Most. In the wider area near Foča that we investigated, we also found it in four caves located in a canyon of the river Bistrica. In the cave Peruc, it sympartically appears with two other cave isopod species: Mesoniscus graniger and an undescribed Trichoniscus species. After Strouhal (1939a), this taxon (presented there as T. biseriatus ) is present together with Alpioniscus (I.) absoloni assimilis (Strouhal, 1939) in a cave on Visočica Mt. (east of Konjic).
In Ljubačevo cave (Western Bosnia), specimens are unusually small (male 6.2 mm, females 6–7.5 mm), which we can treat as a nanism. Adaptive plasticity has played an important role in the body size shift of this population, whose range is limited to a small environment. The size of the cave was 700 m long, where an isolated cavern in a slightly carstified area most likely provides selective pressure. As a consequence of nanism, there are some unusual morphological characteristics present in males, as described below. In Hrustovača cave (40 km west of Ljubačevo) specimens are of "normal" dimensions (male 9 mm, female 8 mm).
In T. albus matovici n. ssp. tergal glands and subsequent structures (gland-piliferous organs) show different degrees of reduction. Dorsolateral paired tubercles ( Fig. 35 View FIGURE 35 ) of the fourth pleonite are totally reduced and present in the form of a slightly swollen area with sparsely scattered pores and lamina (in a male from the cave Hrustovača in Western Bosnia these structures are completely missing; Fig. 42 View FIGURES 42–45 ). After the description and illustration of the male of T. albus matovici n. ssp. from a cave on Visočica Mt. (central Bosnia) given by Strouhal (1939a) (as T. biseriatus ) it seems that this population has more pronounced tergal gland structures on pleon tergite 4 compared to populations from other localities. These differences in the presence and expression of the above mentioned characters could be the result of regressive evolutionary changes in the process of adaptation to the conditions of an underground habitat (for more details see the discussion section).
In the male from Ljubačevo cave (a population with reduced body dimensions) there is a medio-dorsal swollen structure on the fifth pleonite and traces of it on the pleotelson ( Fig. 45 View FIGURES 42–45 ), so far not recorded in Titanethes . According to their position, these structures correspond to the position of glandular tissue within these segments (tergal glands type 3), presented by Vittori et al. (2012). These structures provide space for the glandular tissue of individuals whose body dimensions are reduced, due to nanism. Also, the glandular pores were not noticeable among the present lamina, consistent with other investigated populations of T. albus matovici n. ssp. ( Figs 40–41 View FIGURES 38–41 ). In addition, because they are not hidden among lamina we can assume that glandular tissue is reduced. Alternatively, their glandular secretion may be different in nature than that of other male tergal glands, and do not require visibly open pores. It is possible that they could produce some volatile compounds, as was partly indicated by Vittori et al. (2012). In the T. albus matovici n. ssp. from Hrustovača cave and T. dahli (i.e. " dahli ", the form of T. albus albus without gland-piliferous organ) that we investigated, we did not observe traces of pores and lamina on the fifth pleonite and the pleotelson as well as in Titanethes females.








No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Titanethes albus matovici
| Karaman, Ivo M. & Horvatović, Mladen 2018 |
Titanethes biseriatus
| : Strouhal 1939 |