
Cetinjella monasterii, Karaman & Horvatović, 2018
|
publication ID |
https://doi.org/10.11646/zootaxa.4459.2.3 |
|
publication LSID |
lsid:zoobank.org:pub:49F5AF2B-E795-404B-A946-2C00DB6011B2 |
|
DOI |
https://doi.org/10.5281/zenodo.5983530 |
|
persistent identifier |
https://treatment.plazi.org/id/40481448-6757-FF97-FF28-F9D62275FEDB |
|
treatment provided by |
Plazi |
|
scientific name |
Cetinjella monasterii |
| status |
|
Cetinjella monasterii View in CoL n. g., n. sp.
Figs 20–34 View FIGURES 20–23 View FIGURES 24–28 View FIGURES 29–31 View FIGURES 32–34
Material examined: HOLOTYPE: male 2.4 mm, Montenegro, Cetinje, Cetinjska pećina cave, 42° 23.314'N, 18° 55.327'E, 28.10.2007, leg. I. Karaman, ( ZZDBE-ISO1051 ) GoogleMaps . PARATYPES: ibid., 28.10.2 0 0 7, leg. I. Karaman, 3♀ GoogleMaps ; ibid., 25.11.2007, leg. I. Karaman & M. Horvatović 2♂, 14♀ GoogleMaps ; ibid., 23.9.2008, leg. I. Karaman, 2♂, 5♀ GoogleMaps .
Etymology. Named after the Cetinje Monastery within whose complex the Cetinjska pećina cave is located.
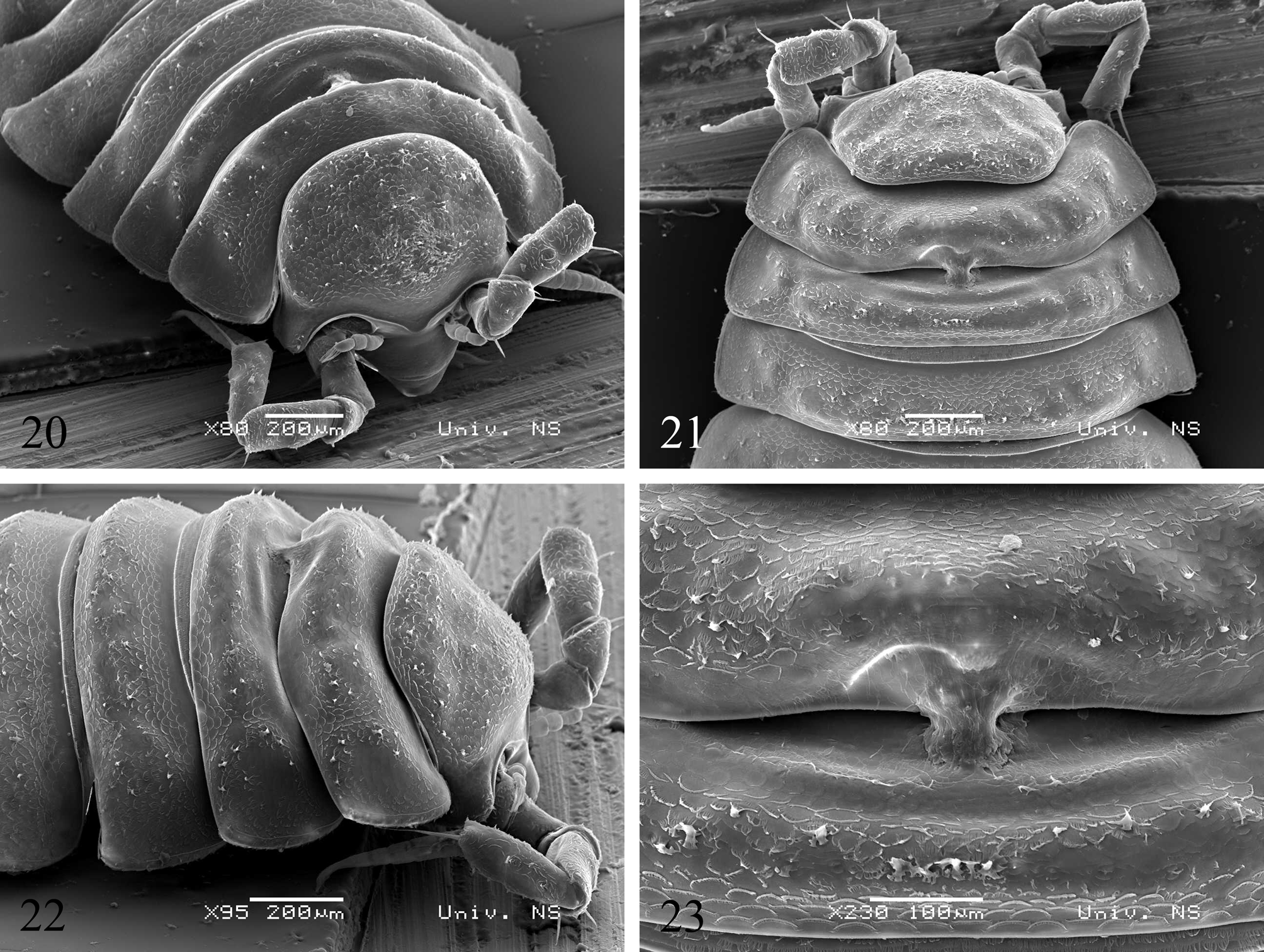
Description. Small troglobite blind species (male up to 2.45 mm, female 3.6 mm long). Body colourless. Cephalon and pereionites almost completely covered with scale rows of different orientations.
Lateral lobes quadrangular, as long as broad.
Pereonite 1 and 2 with distinctive transversal ridge; pereonites 3–4 with weakly expressed ridge, while pereonites 5–7 without ridge. Male pereonite 1 and 2 with gland-piliferous organ ( Figs 20–23 View FIGURES 20–23 ). Pleon short and wide. Pleonites 3–5 without developed epimera.
Telson with concave sides and broadly rounded apex.
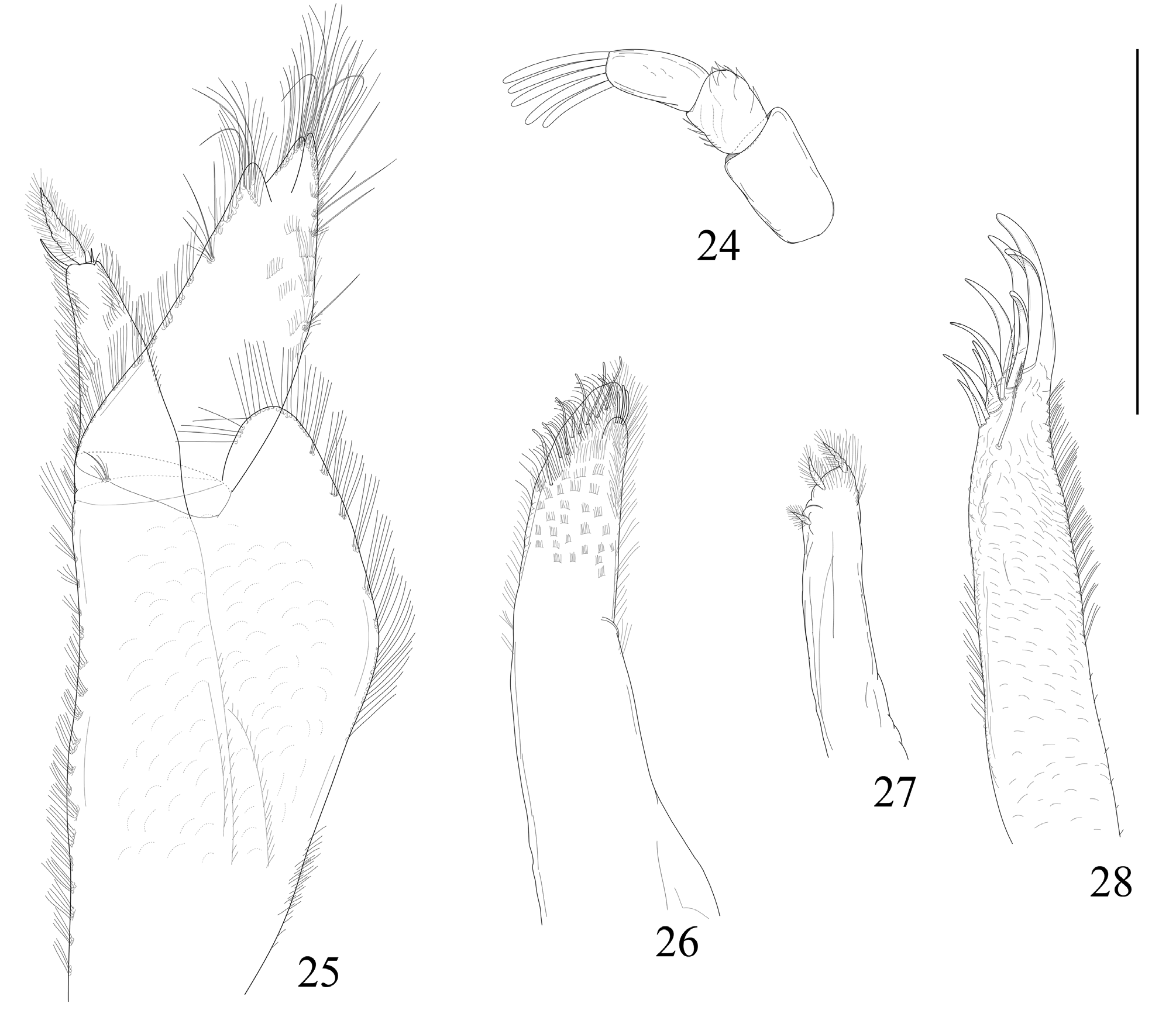
Antennula ( Fig. 24 View FIGURES 24–28 ) basal article 1.5 long as wide; second article as wide as long shorter than basal one; distal article as long as basal one, bearing five aesthetascs. Antenna ( Fig. 29 View FIGURES 29–31 ) short; article 3 slightly longer than wide; article 4 twice as long as wide; article 5 three time longer than wide, as long as flagellum.
Right mandible with two penicils; pars incisiva with three teeth; lacinia mobilis inverted cone shaped, thorny appicaly; pars molaris oval. Left mandible with three penicils; pars incisiva lateralis and pars incisiva medialis with three teeth each; pars molaris ovoid.
Maxilla 1 exopodite ( Fig. 28 View FIGURES 24–28 ) with 3 strong elongated outer teeth, 3 elongated medial teeth, 5 short and sharp inner teeth and two elongate pectinate seta distally. Endopodite ( Fig. 27 View FIGURES 24–28 ) with three very short and thin penicils, two apical ones longer than the proximal penicil. Maxilla 2 ( Fig. 26 View FIGURES 24–28 ) narrow, elongate with setose apical third, bilobate apex; inner lobe remarkable smaller, outer lobe with sparse elongated rod-like setae.
Maxilliped basis short, enlarged on distal portion; outer, inner, and distal margins bearing fringe of thin setae ( Fig. 25 View FIGURES 24–28 ); terminal palp and endite are elongated; palp with pronounced lobe with setae; endite bearing long and thin penicil.
Pereopods short with sharp and notably elongated fused dactylus and unguis; fused dactylus and unguis almost of the same length as propodus. Pereopod 1 ( Fig. 30 View FIGURES 29–31 ) carpus robust bearing 5 strong seate on sternal margin; ischium, merus, carpus and propodus of equal length. Pereopod 7 ( Fig. 31 View FIGURES 29–31 ) merus and carpus bearing 5 strong seate on sternal margin, propodus 3; carpus and propodus of equal length.
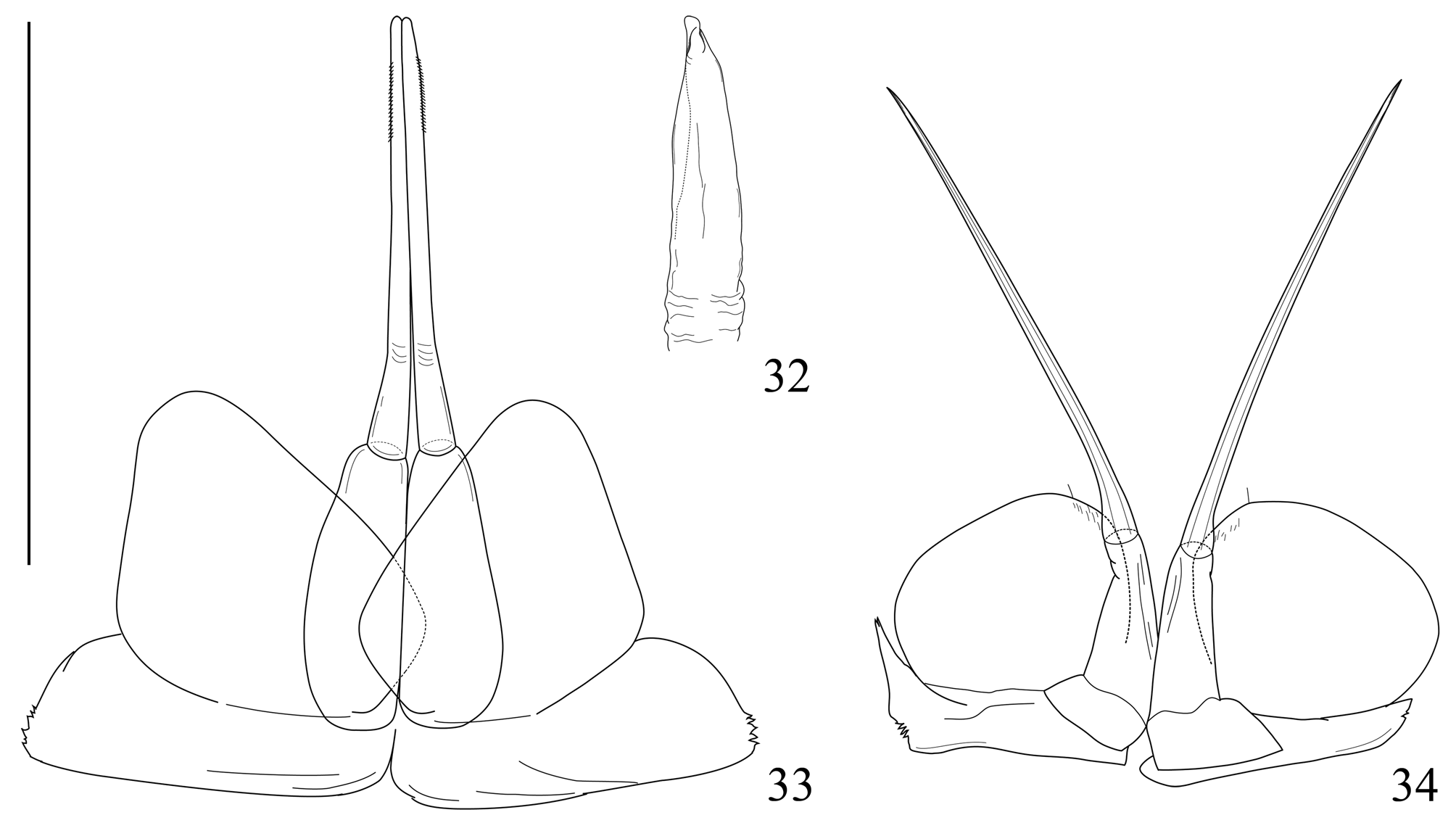
Genital papilla ( Fig. 32 View FIGURES 32–34 ) short and stout, apical part conical.
Pleopod 1 ( Fig. 33 View FIGURES 32–34 ) endopodite of two articles, distal article 1.5 time longer than basal one; basal article shorter than exopodite; endopodite distal article rod like with a crease in its basal part. Exopodite subrectangular, distal margin rounded. Pleopod 2 ( Fig. 34 View FIGURES 32–34 ) endopodite of two articles, distal one filiform, three time as long as basal one. Exopodite oval and short, as long as endopodite basal article.
Uropod endopodite shorter than exopodite, somewhat proximally inserted. Exopodite wide, significantly wider than endopodite.
Remarks. Scale like structures are of different orientations in frontal and caudal half of each pereionite, suggesting their water conducting system functions like that already known on the legs and antennae of oniscids. Scales in this formation seems to retain and direct the water toward the space between tergites, i.e. the dorsal part of water-conducting system. Such an arrangment of scales indicates that the most prominent dorsal parts of pereonites that are on the ridgeline of the frontal and caudal surfaces could have a role in taking and directing gravitational water (small hanging drops from rocks surfaces) toward the water-conducting system. The arrangement of these scales is a bit more complex on the cephalon, dirrecting water mainly toward the groove of the suprantennal line ( i.e. through its medial part toward the clypeus) and to a lesser extent toward the lateral lobi and occipital region ( Figs 20, 22 View FIGURES 20–23 ).
Cetinjella monasterii n. g., n. sp. is clearly distinguished from the Cyphonethes species complex, primarily by a number of mouth part characteristics, which reflect the different lifestyle of the species rather than a large phyletic distance in relation to the Cyphonethes species complex. Males of C. monasterii and Cyphonethes herzegowinensis possess a gland-piliferous organ on pereonite 1 and 2 that is similar in form, indicating their phyletic closeness rather than convergence. Male pleopod structures of Cetinjella monasterii n. g., n. sp. indicate its clear affinity to the genus Cyphonethes .
All specimens of C. monasterii n. g., n. sp. were found in a typical hygropetric habitat on cave wall flowstones with water seeps, suggesting the possible nature of this species. The pereopode structure with elongated fused dactylus and unguis seems adapted for movement through this water flow. Such elongated terminal parts in the walking legs are characteristic of aquatic isopods. At similar places in the same cave, another, still undescribed cave isopod species is present, with mouth part adaptations suggesting it is a hygropetric inhabitant. In the same cave Cyphonethes herzegowinensis is present as the dominant cave isopod species.

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
SuperFamily |
Oniscoidea |
|
Family |
|
|
Genus |