
Synhalcurias elegans ( Wassilieff, 1908 )
|
publication ID |
https://doi.org/ 10.11646/zootaxa.5048.4.5 |
|
publication LSID |
lsid:zoobank.org:pub:4EB228BE-313B-4092-B776-B145A9335B2A |
|
DOI |
https://doi.org/10.5281/zenodo.5570426 |
|
persistent identifier |
https://treatment.plazi.org/id/4103C076-FFBB-FF9C-FF29-341BFB88FC61 |
|
treatment provided by |
Plazi |
|
scientific name |
Synhalcurias elegans ( Wassilieff, 1908 ) |
| status |
|
Synhalcurias elegans ( Wassilieff, 1908) View in CoL
(Japanese name: seitaka-kawari-ginchaku)
Figs. 2–4 View FIGURE 2 View FIGURE 3 View FIGURE 4 ; Table 1 View TABLE 1
Ilyanthopsis elegans Wassilieff, 1908: 8 View in CoL , textfigs 2–5, pl. 1 fig. 2, pl. 3 fig. 38, pl. 4 figs. 39, 40a, 40b.
Synhalcurias elegans: Carlgren, 1914: 50–53 View in CoL , pl. figs. 1–4; Carlgren, 1918: 6, 27, textfigs 2-4; Stephenson, 1922: 260; Carlgren, 1940: 22; Carlgren, 1949: 20; Uchida, 1992: 129, pl. 29 fig. 5; Uchida and Soyama, 2001: 21; Uchida, 2007: 23–27, fig. 2,3, pl. 2. xxx
Material examined. Types ( Fig. 2 View FIGURE 2 ). ZSM2004147 View Materials (syntype): specimen collected from Sagami Bay , Japan by Franz Doflein on May 1904 . ZSM2004148 View Materials (syntype): specimen collected from Sagami Bay , Japan, at 110 m, by Allan Owston using his yacht “Golden Hind” on July 24, 1902 . Other specimens ( Fig. 3 View FIGURE 3 ). NSMT-Co 1693: from Mikawa Bay , specimen dissected and fixed by Takato Izumi on June 1, 2016 . NSMT-Co 1694: collected January 2018, at Uragami , Wakayama Prefecture, at 100–130 m, by Isao Hirabayashi, and dissected and fixed by Takuma Fujii and Kensuke Yanagi on May 22, 2018 .
Description. External anatomy. Body cylindrical ( Fig. 3A View FIGURE 3 ), 7–15 cm in length and 50–100 mm in diameter in living specimens, 50–100 mm in length and 35–80 mm in diameter in preserved specimens. Column surface relatively smooth, wrinkled but without any structures. Ectoderm of column pale orange or yellow when alive ( Fig. 3A View FIGURE 3 ), with sparse, small nematocyst batteries. The naked column opaque milky white. Basal disc in aboral end, thick, robust and adhesive ( Fig. 3A View FIGURE 3 ). Distal column not developed into lobes ( Fig. 3B View FIGURE 3 ). Tentacles 76–100 in number on oral disc, all marginal, in two rows. All tentacles 5–10 mm in length, pale orange in color, long and slender without any structures and no thickenings. Tentacles less contractile and less adhesive. Oral disc pale orange, with radial stripes corresponding to every tentacle. Mouth at center of oral disc, lip-like, pale orange and yellow in color ( Fig. 3A, B View FIGURE 3 ).
Internal anatomy. Perfect mesenteries 76-100, extending from distal to proximal ends; 12 in first cycle, including four directives; eight in second cycle; 16 in third cycle; and 32 in fourth cycle; fifth cycle incomplete. Mesenteries of second cycle in endocoels of mesenteries of first cycle, same as general arrangement of Actinernoidea . Microcnemes absent ( Fig. 3C View FIGURE 3 ). Tentacular longitudinal and circular muscles both too weak to observe in histological sections ( Fig. 3F, G View FIGURE 3 ). Mesoglea on aboral ends of tentacles not thickened like Actinernus . Retractor muscles extremely weak and diffuse ( Fig. 3C–E View FIGURE 3 ); muscle processes too weak to count ( Fig. 3D, E View FIGURE 3 ). Parietal muscles of mesenteries very weak and indistinct, with few muscle fibers ( Fig. 3D, E View FIGURE 3 ). Mesoglea thickest in body wall and actinopharynx ( Fig. 3C View FIGURE 3 ), 2–5 mm, far thicker than ectoderm and endoderm; mesoglea thinner in mesenteries ( Fig. 3D, E View FIGURE 3 ) and basal disc, thinnest in tentacles. Two siphonoglyphs connected to actinopharynx on dorsal and ventral sides, with 18–22 longitudinal grooves ( Fig. 3C View FIGURE 3 ). Marginal sphincter muscle and basilar muscles both absent. Dioecious: almost all mesenteries fertile, immature spermatic cysts in the specimens examined.
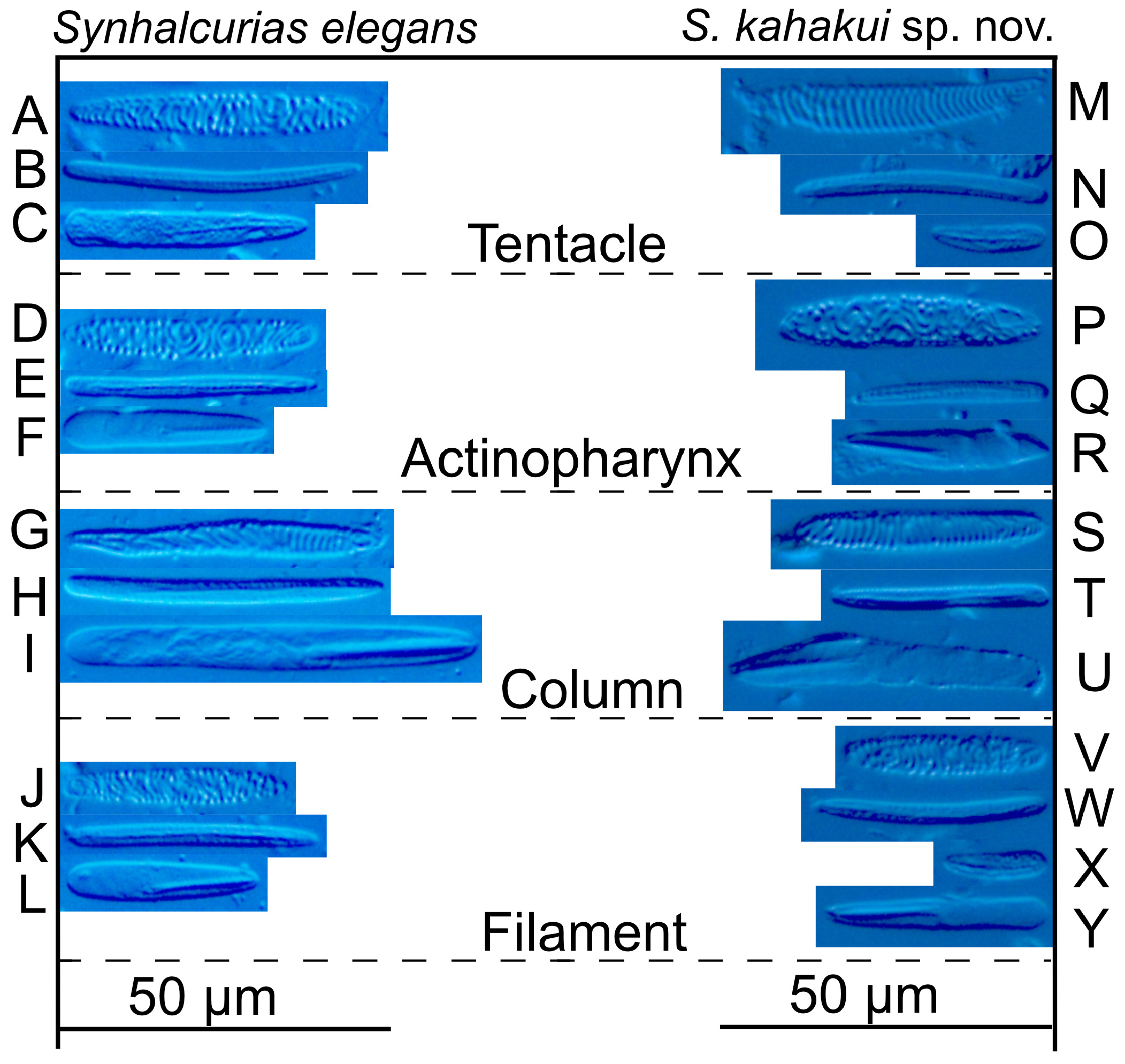
Cnidom. Basitrichs, spirocysts, microbasic b -mastigophores and microbasic p -mastigophores. See Table 1 View TABLE 1 and Fig. 4A–L View FIGURE 4 for size and distribution. Spirocysts in the column of the syntype ( ZSM2004147 View Materials ) are slightly distinguished into three types: small, gracile, and robust ones .
Distribution. Sagami Bay, Kanagawa Pref. and Suruga Bay, Shizuoka Pref. ( Wassilieff, 1908); Ogasawara Islands, Tokyo Met. ( Carlgren, 1918); Suruga Bay, Shizuoka Pref., Taiji and Kushimoto, Wakayama Pref., Enshu- Nada Sea, Aichi Pref. ( Uchida, 2007); Mikawa Bay, Aichi Pref., and Uragami, Wakayama Pref. (present study), all in Japan. All localities situated off the Pacific coast of Japan except for the Ogasawara Islands (see remarks of Synhalcurias kahakui sp. nov.).
Remarks. This species was originally described as Ilyanthopsis elegans by Wassilieff (1908). However, this genus was abolished by Carlgren (1914) because its type species, I. longifilis Hertwig (1888) , had been already synonymized with Condylactis passiflora Duchassaing and Michelotti, 1864 (currently considered a junior synonym of Condylactis gigantea [ Fautin, 2016]) by Pax (1910). Thus, Carlgren established a new genus, Synhalcurias Carlgren, 1914 , for I. elegans . This new genus was monotypic.
Since its original description and additional specimens in Carlgren (1918), Synhalcurias elegans had not subsequently been documented until Uchida (2007); he redescribed S. elegans using specimens collected from several localities of Japan (see distribution). The characters of our specimens almost perfectly correspond to the redescription of Uchida (2007) and the two syntypes. There is a slight difference in the cnidae between the type material, description of Uchida (2007) and our specimens: the spirocysts of the column can be distinguished into the three types in the type specimen (ZSM2004147), into two types according to Uchida (2007), but are of only one type in our specimen (NSMT-Co 1693). However, there were several cnidae with intermediate form between the two types, thus this difference should not be a conclusive factor to differentiate the species. Thus, we identified these specimens as S. elegans , and redescribed this species by emending mesenterial arrangements and histology of the description of Uchida (2007) and added images of the cnidae. Note that Carlgren (1918) mentioned hermaphrodite specimens; we observed only gonochoric individuals and only immature spermatic cysts.


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Synhalcurias elegans ( Wassilieff, 1908 )
| Izumi, Takato & Yanagi, Kensuke 2021 |
Synhalcurias elegans: Carlgren, 1914: 50–53
| Uchida, H. 2007: 23 |
| Uchida, H. & Soyama, I. 2001: 21 |
| Uchida, H. 1992: 129 |
| Carlgren, O. 1949: 20 |
| Stephenson, T. A. 1922: 260 |
| Carlgren, O. 1918: 6 |
| Carlgren, O. 1914: 53 |
Ilyanthopsis elegans
| Wassilieff, A. 1908: 8 |