
Cryptophractus pilosus Fitzinger, 1856
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3947.1.2 |
|
publication LSID |
lsid:zoobank.org:pub:A0C5C434-AFA5-4BA3-B1F2-0569EF07F483 |
|
DOI |
https://doi.org/10.5281/zenodo.5621912 |
|
persistent identifier |
https://treatment.plazi.org/id/414787E9-7934-FFA3-ABBD-FA16FEAAFCE7 |
|
treatment provided by |
Plazi |
|
scientific name |
Cryptophractus pilosus Fitzinger, 1856 |
| status |
|
Cryptophractus pilosus Fitzinger, 1856 [nomen revalidatum]
Cryptophractus pilosus Fitzinger, 1856:123 . Holotype NMW 222; type locality Peru. Praopus hirsutus Burmeister, 1862:147 . Possibly MUSM 27 (see above); type locality “Guayaquil”, Peru. [ Tatusia (Cryptophractus) ] pilosa: Trouessart, 1898:1140 View in CoL ; name combination.
[ Tatus (Cryptophractus) ] pilosus: Trouessart, 1905:814 ; name combination.
Tatu pilosa: Thomas, 1927:605 View in CoL ; name combination.
Dasypus pilosa: Yepes, 1928:468 View in CoL ; name combination with incorrect gender agreement. Dasypus pilosus: Frechkop & Yepes, 1949:27 View in CoL ; gender agreement correction.
Dasypus (Cryptophractus) pilosus: Talmage & Buchanan, 1954:84 View in CoL ; name combination.
Holotype. NMW 222, mounted skin ( Fitzinger 1856).
Type locality. Montane Peru (restricted by Wetzel & Mondolfi 1979).
Distribution. Montane cloud forests and subparamo of Peruvian Andes, in the departments of San Martín, La Libertad, Huánuco, Junín, Amazonas ( Grimwood 1969; Wetzel & Mondolfi 1979; Wetzel et al. 2007) and expanded herein to Pasco ( Fig. 3 View FIGURE 3 ). The specimens examined were generally collected close to water bodies, between 2,600 to 3,400 m above the sea level.
Diagnosis. Dense hair covering the whole carapace, obscuring it; cephalic shield narrow and elongate, loosely delimitated anteriorly, with acute posterior margin more distantly separated from the base of the ears, and V-shaped occipital sulcus; greater number of completely movable bands (9 to 11); osteoderms with no sulci, numerous foramina (more than 30), and frequently two larger foramina placed anteriorly in the movable osteoderms; rostrum and palate longer and narrower (rostral ratio 0.66 on average; see Wetzel & Mondolfi [1979] for description on how to take this measure); condylar process of mandible lower and sessile; and minute teeth.
Comparative description. Externally, Cryptophractus pilosus has the whole carapace covered by a thick coating of rigid, long, yellow to light-brown hairs that completely conceal the carapace, but for the antero-lateral margin of the scapular buckler ( Fig. 4 View FIGURE 4 ). The same type of hair is present in the cheeks and proximal portion of the limbs; shorter and less abundant hair covers the ventral surface of the body, where brownish transversal lines in the skin are in continuation to the movable bands. These areas also show some flat isolated osteoderms within the skin. Cryptophractus pilosus has five digits in the feet and four in the hands; a vestigial fifth digit is externally visible only in D. kappleri (but see Costa & Vizcaíno 2010). The ears are large and conical, covered with pebbly skin, whereas the eyes are a small slit.
The external measurements (Table 1) agree with previous observations on the species ( Fitzinger 1871; Flower 1886; Frechkop & Yepes 1949; Wetzel & Mondolfi 1979; Wetzel 1985). Only three adult specimens analyzed have weight data (MUSM 7501=2.30 kg; MUSM 7504=1.40 kg; MUSM 7505=1.85 kg), being on the upper range or slightly heavier than D. septemcinctus (1.45–1.80 kg), D. hybridus (2.04 kg) and D. sabanicola (1.00–2.00 kg) ( Hamlett 1939; Wetzel & Mondolfi 1979). The average total length also indicates that C. pilosus is larger than these species and D. yepesi , whereas is smaller than D. novemcinctus and D. kappleri ( Wetzel & Mondolfi 1979) . As observed in D. novemcinctus ( McBee & Baker 1982) , apparently the males (MUSM 2056, 7501) of C. pilosus are slightly larger than the females (MUSM 7504, 7505), as shown in Tables 1–3.
TABLE 1. External lengths of Cryptophractus pilosus (mm), taken from dried specimens. Averages calculated excluding the juvenile specimens.
With respect to the carapace, the cephalic shield is narrow and elongate, roughly rhomboid, and the osteoderms does not form rosettes; the posterior end is acute, more distantly separated from the base of the ears, and has a Vshaped occipital sulcus; anteriorly, the osteoderms become gradually irregular in shape, loosely articulated to each other, and arranged in lines parallel to the sagittal plan. The morphology of the cephalic shield differs from that of Dasypus spp., as was illustrated by Frechkop & Yepes (1949) and Kraft (1995). The pelvic buckler is variably longer than the scapular buckler ( Table 2 View TABLE 2 ). The latter has a round anterior border (angular only in D. kappleri ). The number of completely movable bands ranges from 9 to 11 (n=8). Besides of being more numerous than in Dasypus spp. ( Hamlett 1939; Wetzel & Mondolfi 1979; Vizcaíno 1995), each band is anteroposteriorly shorter. Also our observation on a dry carapace (MUSM-ING 633) suggests limited mobility among the four anterior bands, which seem to be fused with each other. The cylindrical tail corresponds to 65–76% of the head-body length, similarly to D. kappleri and D. novemcinctus and much longer than the other species of Dasypus ; its posterior third gradually tapers, whereas the proximal two-thirds is partitioned in 7–11 rings (n=8), each composed of two rows of osteoderms with scarce hairs on the posterior margin of each ring.
The morphology and ornamentation of the osteoderms are described here for the first time confirming that it is unique in relation to all extant armadillos ( Fig. 4 View FIGURE 4 and 5 View FIGURE 5 ). They lack principal or radial sulci. The foramina of the buckler osteoderms are arranged in a central circle bounding a nearly circular central figure, which have small neurovascular foramina inside it. A second row of similar foramina forms a posterior semicircle. The total number of foramina per osteoderm (including some confluent ones) is about 30 in the scapular and 35 in the pelvic osteoderms. The pelvic elements have bigger and more lacerated foramina. Scapular semimovable osteoderms have a semicircular arch of foramina in the mid part and at least two rows of foramina in the posterior border, together forming a nearly circular figure. Pelvic semimovable and movable osteoderms have one or two large foramina anteriorly. The foramina are distributed within an inverted “V” pattern in the formers, and their size decrease posteriorly. The foramina in the movable osteoderms are arranged in two posteriorly divergent lines, which are connected distally by a semicircular arc, forming a triangular principal figure; a circular figure is limited anteriorly by an additional, but inverted, semicircular arc of foramina at mid-length; the posterior margin is copious of foramina, arranged in two or three rows; the size of the foramina also decrease posteriorly. The observations on two specimens (MUSM 24214 and MUSM-ING 633) indicate that, despite the great size of the foramina in the osteoderms, a single hair comes out of each foramen.
The horn scales are thicker than in Dasypus spp., with irregular borders and numerous perforations for hair emersion. Several depositional layers are visible from inside to outside. Each buckler osteoderm is covered by a central scale and about six peripheral scales that also cover part of adjoining osteoderms. In movable osteoderms the principal figure is covered by a triangular and a circular scale, and inverted triangles cover the peripheral figures of two adjoining elements ( Fig. 5 View FIGURE 5 ).
The skull of Cryptophractus pilosus is unique in having the longest and narrowest rostrum of all extant armadillos. The lateral margins of the rostrum are sub-parallel in dorsal view but its anterior tip is slightly broader ( Fig. 6 View FIGURE 6 A). A delicate sagittal crest is present in most of the adult specimens (e.g., MUSM 7504), a rare feature in Dasypus spp. Anterior to the interorbital constriction the frontals are delicately vaulted by the development of frontal sinuses. The anterior processes of the frontals are more elongated than in Dasypus to contact the longer nasal bones.
In the ventral aspect of the skull ( Fig. 6 View FIGURE 6 B), the maxillae, and occasionally the premaxillae, are concave along the sagittal line. The palate corresponds to 68–74% (70% on average, n=7) of the maximum skull length, including juveniles, confirming the observations of Wetzel & Mondolfi (1979). In relation to D. novemcinctus , the increase of the palatal length in C. pilosus is achieved by the extension of the maxillae and premaxillae, whereas the palatines remain proportionally equal. The palatines are flat, with a heart-shaped anterior border and rounded lateral margins. The posterior border of the palate, comprising palatines and pterygoids, is indented medially with a contour varying from a long and narrow “V” to bell-shaped. This condition differs from D. kappleri , which has a straight and roughly square internal nasal aperture. The free border of the pterygoids has a sharp ventral projection, forming a triangular outline in lateral view. The glenoid articulation is oval and flat, with a ventrally projecting postglenoid process. The foramen ovale and the transverse canal foramen are separated in all available specimens, whereas these are confluent in some specimens of Dasypus spp.
In lateral view ( Fig. 6 View FIGURE 6 C), the dorsal contour of the rostrum is approximately straight, as in D. hybridus and D. septemcinctus , whereas it is markedly sigmoid in the other Dasypus species. The zygomatic arch is dorso-ventrally thinner than in D. novemcinctus and the crest for insertion of masseter muscle is weaker. The lacrimal is proportionally smaller, as in D. hybridus , and possesses one or exceptionally two lacrimal foramina. The morphology of the contact between the petrosal and the squamosal varies from a shallow occipital groove in the youngest specimen (MUSM 7503) to an occipital canal with different degrees of closure in the adults, as described for D. novemcinctus ( Wible 2010) . Five specimens (MUSM 7500, 7501, 7503, 7504, and 7505) preserve the ectotympanic and the ossicles of the middle ear. Adults present an ossified entotympanic that partially covers the ventral part of the petrosal, running medially to the ectotympanic from its the posterior part until the alisphenoidpterygoid suture ( Fig. 7 View FIGURE 7 ); only in MUSM 7503, a juvenile, the ossification of the entotympanic has a middle gap between two ossifications. In D. novemcinctus , the only extant Dasypus with detailed studies on the auditory region, the entotympanic is poorly developed, composed by small isolated ossifications ( Patterson et al. 1989; Wible 2010). The same pattern was observed in D. septemcinctus (MNRJ 10062; Fig. 7 View FIGURE 7 ).
The slender mandible ( Fig. 6 View FIGURE 6 D) shows a straighter horizontal ramus, much slender than in extant related species. The angular process is blunt and the condylar process is lower and sessile (whereas it has a neck with variable development in Dasypus ).
Table 3 View TABLE 3 shows some cranial measurements of Cryptophractus pilosus . They are consistent with previous observations on the species ( Wetzel & Mondolfi 1979; Wetzel 1985). The skull length and the zygomatic width are only smaller than in D. kappleri and than the largest specimens of D. novemcinctus . With respect to the rostral ratio (=length of the rostrum/maximum skull length), excluding juvenile specimens, it varies between 0.62 and 0.69, with an average of 0.66. According to data presented by Wetzel & Mondolfi (1979), this ratio is slightly higher than that of D. kappleri (0.64 on average) and sensibly exceeds those of Dasypus (Dasypus) group, which are mostly below 0.62 in D. novemcinctus and about 0.56 in D. septemcinctus , D. hybridus , and D. sabanicola ; the rostral ratio of D. yepesi (0.55, calculated for MLP 30-III-90 -8) agrees with the other small species.
Like in Dasypus ( Ciancio et al. 2012) , the peg-like molariforms of Cryptophractus pilosus have an anteriorposterior beveled occlusal relief ( Figs. 6 View FIGURE 6 C and 6D). However, the teeth are substantially smaller (with approximately 1 mm long and 0.7 mm wide) and subcylindrical (instead of prismatic with subrectangular crosssections). There are 7 to 8 upper and 7 to 9 lower molariforms, a tooth count higher than the 5/6 observed by Wetzel (1985). The dental series occupy only a short portion of the maxilla and mandible, leaving an extensive edentulous anterior section, longer than the dental series itself. Closed or aborted alveoli are highly frequent in the material analyzed. Previous studies in Dasypus spp. showed that a thin enamel layer is present in the apical portion of deciduous and permanent teeth, but is lost soon after the eruption; also the two real cusps (labial and lingual), are worn leaving secondary labial and lingual borders ( Martin 1916; Ciancio et al. 2010). Further studies are required to confirm the presence of dental enamel in C. pilosus .
A single specimen (MUSM 24214) has appendicular and axial elements preserved, mostly with unfused epiphyses, suggesting an immature condition of the skeleton. It includes three thoracic and five lumbar vertebrae, sinsacrum, right femur, tibia-fibula, astragalus, calcaneus, navicular, plantar sesamoid, metatarsals, and phalanges. Anatomical comparisons were made among these elements and those of Dasypus species (including a young juvenile specimen of D. septemcinctus ), except for D. sabanicola . Figure 8 View FIGURE 8 compares equivalent postcranial elements of C. pilosus and D. novemcinctus . In C. pilosus , the lumbar vertebrae have shorter neural spines and metapophyses and, in lateral view, the angle formed between these apophyses is wider. The morphology of the xenarthrous articulations is concordant with the description of Gaudin (1999). The synsacrum is proportionally longer, composed of seven vertebrae (varying from seven to nine in Dasypus spp.), although the irregular posterior margin of the transverse processes of the last one suggests the presence of an eighth posteriorly incorporated element. The iliac crest is more recurved and more laterally projected, as wide as the ischia. The femur is slenderer; oddly a hair pierces the posterior surface of the shaft. The tibia and fibula are not yet fused at their distal ends, as observed in adults of Dasypus . Compared to D. novemcinctus and D. hybridus , the astragalus of C. pilosus has a longitudinal separation in the tibial facet, the neck lacks ridges on the dorsal surface, and the sustentacular and navicular facets are larger. Both calcaneal heel and head are proportionally broader, the ectal facet extends further over the calcaneal heel, and laterally the peroneal process is incipient. Jasinski & Wallace (2014) hypothesized that a greater development of this process (as present in D. novemcinctus ) may be related to semifossorial habits. No differences were observed in the remaining elements.
Habitat and biology. The predicted distribution of C. pilosus (NatureServe 2014) includes a narrow montane strip in Peru from Amazonas to Junín Departments. Consequently, due to deforestation of montane habitats the conservation status of C. pilosus is vulnerable ( Pacheco 2002; Superina & Abba 2010). Its ecology and behavior are completely unknown ( Wetzel et al. 2007). In this regard we report the occurrence of multiple embryos in the species based on the observation of a female with four embryos (MUSM 7504). Also, a tick identified as Ixodes sp. was found attached to the skin of other specimen (MUSM 7499).
With respect to diet, it is predominantly based on insects in D. sabanicola and in D. novemcinctus ( Mondolfi 1967, McBee & Baker 1982, Smith & Redford 1990). Numerous morphological adaptations for myrmecophagy (=termitophagy) were recognized in D. novemcinctus ( Moeller 1968; Wetzel & Mondolfi 1979; Smith & Redford 1990; Charles et al. 2013). In C. pilosus , some of these characters are even more accentuated, such as a longer and narrower rostrum, the shorter dental series composed of minute teeth, frequent closed/aborted alveoli in adults, and lower and sessile mandibular condyle. These features suggest a predominantly ant and termite-eating diet, although no direct observations of feeding activity has been recorded. Also, the skull, mandible, and teeth of the mid-sized fossil armadillo Stegotherium (early–middle Miocene of Patagonia) shows similarities to C. pilosus . Although phylogenetic studies support a close relationship between this genus and Dasypodini ( Engelmann 1985; Gaudin & Wible 2006; and the present cladistic analysis), the resemblances between Stegotherium and C. pilosus might reflect a convergence related to their feeding habits, as a myrmecophagous diet was also inferred for the former ( Vizcaíno 1994).
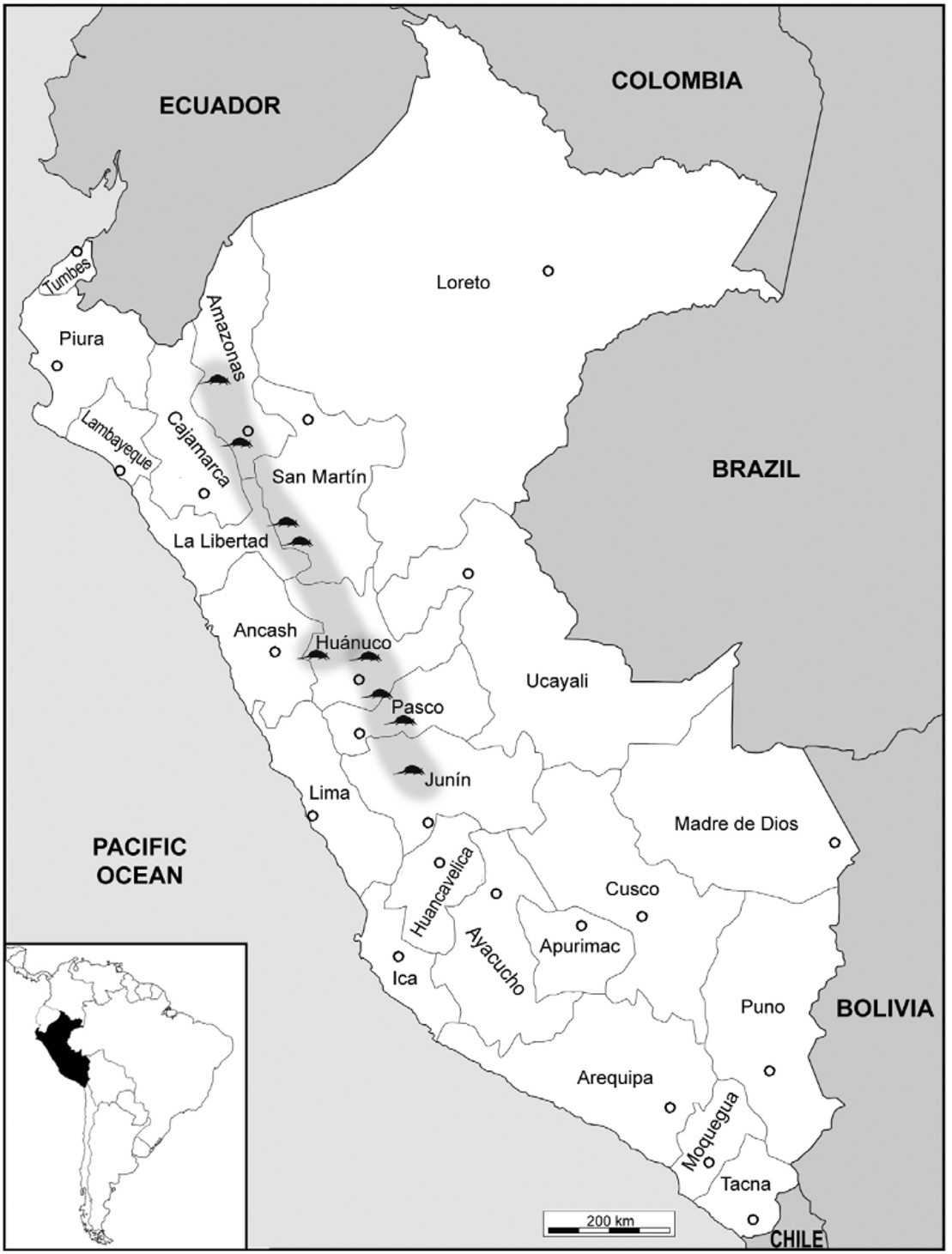


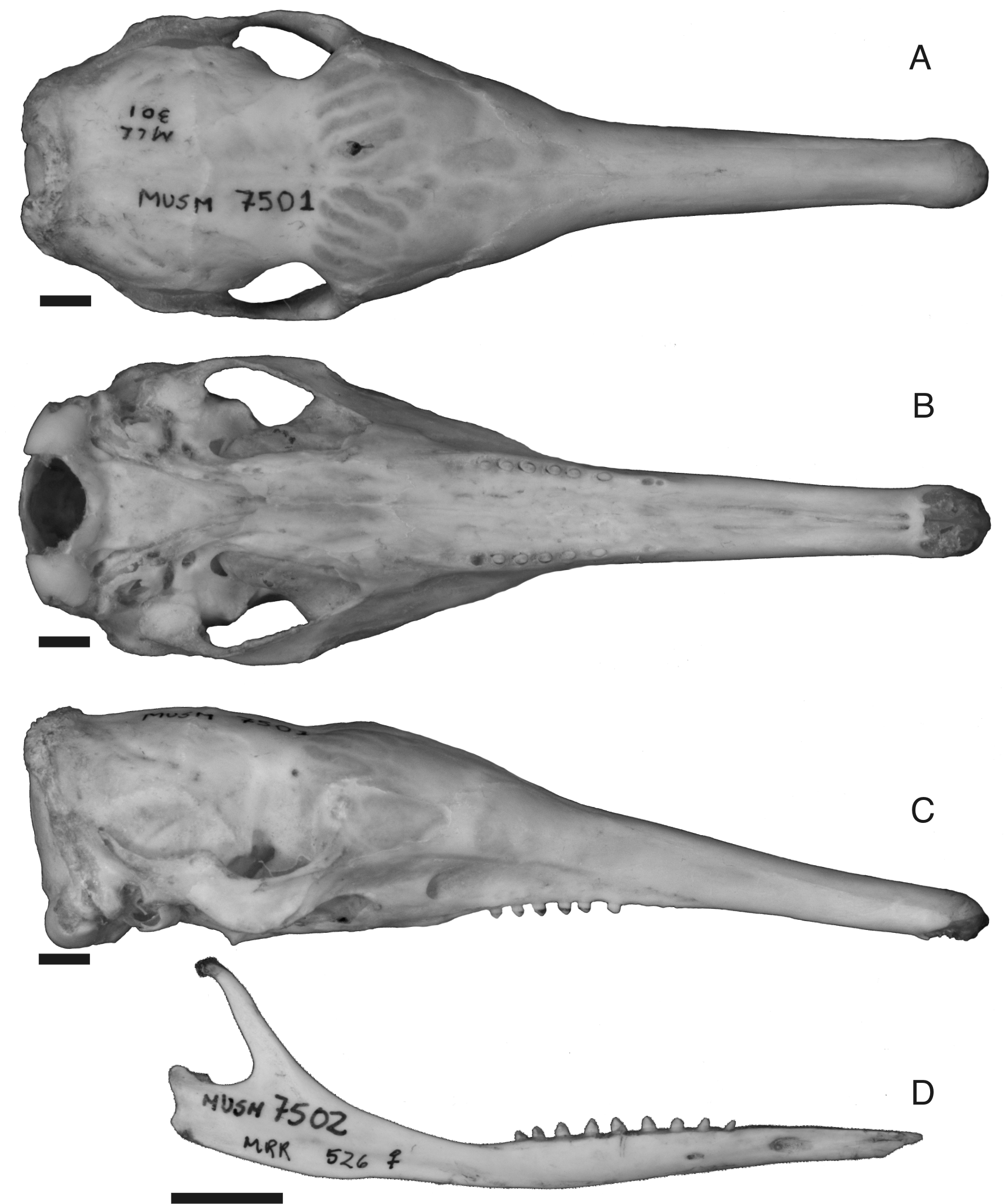


TABLE 2. Carapacial measurements of Cryptophractus pilosus (mm), taken along the sagittal line.
| Total | Head | Carapace | Tail | Hind foot Ear | |
|---|---|---|---|---|---|
| MUSM 2056 | 750 | 112 | 287 | 261 | 70 50 |
| MUSM 7499 | — | 117 | 237 | — | — 35 |
| MUSM 7501 | 648 | 125 | 266 | 268 | 80 43 |
| MUSM 7502* | — | — | 186 | 181 | 53 43 |
| MUSM 7503* | 413 | 80 | 159 | 162 | 56 33 |
| MUSM 7504 | 593 | 116 | 205 | 245 | 71 41 |
| MUSM 7505 | 650 | 122 | 233 | 260 | 75 42 |
| MUSM-ING 633 | — | — | 231 | 253 | 71 — |
| MUSM-CT 1312* | — | — | 167 | 146 | — — |
| Average | 660 | 118 | 236 | 243 | 74 44 |
| *juvenile specimens. |
TABLE 3. Cranial measurements of Cryptophractus pilosus. Abbreviations: MSL, maximum skull length; PL, palatal length at sagittal line; RL, rostral length; R ratio, rostral ratio (= RL / MSL); ZW, zygomatic width; ManL, maximum mandibular length. Averages calculated excluding the juvenile specimens.
| Specimen | MSL | PL | RL | R ratio | ZW | ManL |
|---|---|---|---|---|---|---|
| MUSM 2056 | 109.4 | 80.5 | 75.2 | 0.69 | 35.1 | 90.6 |
| MUSM 7500 | 110.7 | 75.5 | 71.8 | 0.65 | 36.2 | 90.4 |
| MUSM 7501 | 109.3 | 79.4 | 72.3 | 0.66 | 34.5 | 88.2 |
| MUSM 7502* | 89.0 | ~61 | 54.8 | 0.62 | 31.1 | 68.0 |
| MUSM 7503* | 69.8 | 51.1 | 43.4 | 0.62 | 24.1 | 61.0 |
| MUSM 7504 | 99.9 | 68.6 | 65.2 | 0.65 | 32.1 | 80.9 |
| MUSM 7505 | 105.9 | ~69 | 66.1 | 0.62 | 35.0 | 86.2 |
| Average | 102.9 | 68.6 | 65.7 | 0.64 | 33.6 | 83.6 |
| *juvenile specimens. |
| NMW |
Naturhistorisches Museum, Wien |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Cryptophractus pilosus Fitzinger, 1856
| Castro, Mariela C., Ciancio, Martín R., Pacheco, Víctor, Salas-Gismondi, Rodolfo M., Bostelmann, J. Enrique & Carlini, Alfredo A. 2015 |
Dasypus (Cryptophractus) pilosus:
| Talmage 1954: 84 |
Dasypus pilosa:
| Frechkop 1949: 27 |
| Yepes 1928: 468 |
Tatu pilosa:
| Thomas 1927: 605 |
Tatus (Cryptophractus)
| Trouessart 1905: 814 |
Cryptophractus pilosus
| Trouessart 1898: 1140 |
| Burmeister 1862: 147 |
| Fitzinger 1856: 123 |