
Hyperxiphia hirashimai ( Okutani, 1965 ), 2020
|
publication ID |
https://doi.org/10.11646/zootaxa.4822.3.5 |
|
publication LSID |
lsid:zoobank.org:pub:C199B68E-E676-42CA-89BA-1415236ACFF4 |
|
DOI |
https://doi.org/10.5281/zenodo.4450912 |
|
persistent identifier |
https://treatment.plazi.org/id/421AB335-FFA5-FFEA-FF17-FC61BC83FDCD |
|
treatment provided by |
Plazi |
|
scientific name |
Hyperxiphia hirashimai ( Okutani, 1965 ) |
| status |
comb. nov. |
Hyperxiphia hirashimai ( Okutani, 1965) , n. comb.
( Figs 2–5 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 )
Genaxiphia hirashimai Okutani, 1965: 74 ; Abe & Togashi, 1989: 559; Smith, 1978: 105; Taeger et al., 2010: 113; Shinohara, 2019d: 21; Shinohara, 2020: 470.
Female (hitherto undescribed, based on a specimen from Narukawa-keikoku, Fig. 2 View FIGURE 2 ). Length without ovipositor about 12 mm. Head black with extensive creamy white marks as in Figure 2 View FIGURE 2 A–E; mouth parts creamy white, partly obscurely blackish, with apex of mandible black. Antenna entirely black. Thorax black, with following creamy white: large spot in lower part and anterior and dorsal margins of pronotum, spot on anterior lateroventral part of propleuron, most of tegula, outer part of axilla, three spots on mesepisternum (one at ventral anterior margin and two along the dorsal ridge), metascutellum and anterolateral swollen part of metanotum (base of cenchrus); mesoscutal lateral lobe marked with dark brown. Legs black, with part of fore and mid coxae, most of hind coxa, basal half of tibia and tarsomere 1 (except for extreme apex) of all legs creamy white. Wings slightly infuscated; veins and stigma blackish brown. Abdomen black, with following creamy white: tergum 2 (except for anterior and posterior margins), lateral spot on terga 3–9 (very small and obscure on 3, small on 4, large on 5–9, large and along posterior margin on 9), large transverse spot on dorsum of tergum 8, and dorsal apical part of terminal tergum; ovipositor sheath marked with dark brown.
Malar space about 0.4× length of distance between toruli, with ventral pit (antennal groove) large and deep, clearly delimited by shape carina dorsally; space between eye and dorsal margin of ventral pit almost linear; occipital carina (crassa) distinct, ventral end obsolete; genal carina developed nearly to vertex but their dorsal ends widely separated from each other; inner orbits slightly divergent below, interocular distance at level of toruli about equal in length to eye height; wide longitudinal depression from median ocellus to median fovea. Labial palpus with 4 palpomeres; maxillary palpus with 5 palpomeres, terminal palpomere pointed at apex. Both antennae with 17 antennomeres; scape (incl. radicula): pedicel:flagellomere 1:flagellomere 2 as 2.1:1.0:1.6:0.8. Hind tarsomere 1 about 1.2× length of remaining tarsomeres combined; tarsal claws with large inner tooth. Fore wing with cell 3R1 closed at apex (vein R1 connecting with vein Rs); cell 2Rs slightly wider than cell 1M; crossvein 2r-m basal to crossvein 2m-cu on vein M; hind wing with cell R1 closed at apex. Ovipositor sheath with apical sheath subequal in length to basal sheath.
Frons and interantennal area reticulate, with rather conspicuous striae radiating down from ocelli ( Fig. 2D View FIGURE 2 ); gena covered with rough longitudinal wrinkles and some large punctures; vertex smooth, shiny ( Fig. 2A, E View FIGURE 2 ). Pronotum irregularly wrinkled in dorsal half, smooth with several longitudinal ridges at middle, and very smooth and impunctate in ventral half; propleuron roughly wrinkled but without distinct punctures and surface rather smooth; mesoscutal median lobe coarsely reticulate on dorsal surface; lateral lobes reticulate with large elongate even area with coriaceous and mat surface; mesoscutellum densely rugoso-reticulate; mesepisternum rather shallowly reticulate, pilose, with posterior dorsal area largely smooth; mesepimeron nearly impunctate and glabrous, with row of short oblique ridges; metepisternum coarsely rugoso-reticulate and metepimeron rugose. Abdomen with tergum 1 coarsely finely punctate, each half centrally and with broad inner margin partly nearly impunctate and shiny; all other terga weakly shiny with very fine surface microsculpture, posterior dorsal margin of each tergum nearly smooth and shiny.
Male ( holotype). Length about 7 mm. Head creamy white, with black marks as in Figures 3C View FIGURE 3 , 4A, B View FIGURE 4 ; anterior margin of clypeus pale amber; mouth parts creamy white, with apex of mandible black. Antenna dark brown to black; posterior surfaces of scape and pedicel pale brown. Prothorax creamy white, with laterodorsal surfaces of propleuron darkened and dorsal margin and large mark in dorsal lateral surface of pronotum blackish; mesonotum very pale reddish brown, with axilla and dorsal surface of mesoscutellum creamy white and posterolateral sunken area to lateral and posterior surfaces of mesoscutellum black ( Fig. 4D View FIGURE 4 ); metanotum black, with metascutellum and anterolateral swollen part (base of cenchrus) creamy white and sunken area between cenchri pale brown; lateral and ventral surfaces of mesothorax and metathorax creamy white, partly obscurely pale brown. Legs very pale brown, with coxae and trochanters creamy white. Wings very slightly infuscated; veins and stigma dark brown. Abdomen creamy white, with large blackish marks as in Figure 3C, D View FIGURE 3 ; base and apex of each sternum slightly darkened and sterna 8 and 9 largely marked with black; genitalia black.
In structure, generally similar to female, except for usual sexual differences notably in slenderer and longer abdomen. Labial palpus with 4 palpomeres ( Fig. 4E, F View FIGURE 4 ). Both antennae with 15 antennomeres. Tarsal claw with large subapical tooth near apex. Abdominal tergum 8 large and long, nearly flattened above, apical margin nearly truncate; composite terga 9+10 small and short, mostly concealed under tergum 8; sternum 8 with posteromedian part swollen and protruding posteriorly, with conspicuous tuft of long stiff hairs ( Fig. 4G View FIGURE 4 , arrowed); sternum 9 with posterior margin slightly concave medially.
Variation. Females (nine specimens examined): Length without ovipositor ranges from about 7 to 17 mm. The antennae have 15 to 19 (most frequently 17) antennomeres (n=15). Of the three whitish spots on the mesepisternum, one or two spots along the dorsal ridge are sometimes missing or, on the contrary, all the three spots are enlarged and nearly connected into a large elongate mark. Sometimes, the mesoscutal median lobe has an obscure whitish spot, the mesoscutellum has paired spots and the metepisternum has an obscure whitish spot. The mesoscutal lateral lobe is usually entirely black. The abdomen has the whitish area on the tergum 2 sometimes interrupted medially above and the lateral whitish spot on the tergum 3 is often missing. The ovipositor sheath is sometimes entirely black. Males (eight specimens examined including type series): Length ranges from about 6.5 to 14 mm. The antennae have 14 to 17 antennomeres (n=13). The mesoscutal median lobe is sometimes largely creamy white ( Fig. 2A View FIGURE 2 ) and the abdominal tergum 1 is very often largely marked with pale brown ( Fig. 3A View FIGURE 3 ).
Comparative comments. Hyperxiphia hirashimai closely resembles H. triangularis from Taiwan ( Smith 2019; Shinohara et al. 2020b). These two species show a remarkably similar pattern of sexual dimorphism. In the female of H. triangularis , the antennal flagellum is creamy white except for blackish apex, the mesonotum is largely marked with pale brown and creamy white, the trochanters of all legs are creamy white, the abdominal tergum 8 has the broad posterior margin creamy white, and the abdominal tergum 9 is largely creamy white laterally. In H. hirashimai , however, the antennal flagellum is entirely black, the mesonotum is mostly black, the trochanters of all legs are black, the abdominal tergum 8 has separate dorsal and lateral creamy white marks, and the abdominal tergum 9 has the lateral creamy white spot limited to its posterior margin. The males of H. hirashimai and H. triangularis are barely distinguishable by the coloration of the veins C and R in the fore wing and the antennal flagellum. The veins C and R in the fore wing is concolorous pale brown and the flagellum has no whitish areas in H. hirashimai , whereas the vein C is pale brown and the vein R is contrastingly blackish brown and the flagellum often has whitish areas in H. triangularis .
In Maa’s (1949) key to Hyperxiphia species, H. hirashimai goes close to H. melanaria ( Mocsáry, 1904) from Tonkin, but the two taxa are different in details of the coloration of the legs and abdomen, judging from the descriptions of Mocsáry (1904) and Maa (1949). In Smith’s (2019) key to Taiwanese species, the female of H. hirashimai may run to H. sculpturata Smith, 2019 , but differs from it in the pale color pattern of the head (compare Figure 2 View FIGURE 2 C–E with figures 24 and 25 in Smith 2019), the creamy white metascutellum (entirely black in H. sculpturata , figure 23 in Smith 2019), and the densely rugoso-reticulate mesoscutellum (mostly shiny with scattered large punctures separated by shiny flat interspaces in H. sculpturata , figure 23 in Smith 2019). The male of H. hirashimai may key to H. formosana ( Enslin, 1911) , except for the number of the antennomeres (“usually with 17 antennomeres”) in couplet 4, but H. hirashimai has much more pale areas on the dorsum of thorax and abdomen than H. formosana ( Enslin 1911; photographs in Taeger et al. 2018).
Remarkable sexual dimorphism. As described above and illustrated in Figures 2–4 View FIGURE 2 View FIGURE 3 View FIGURE 4 , H. hirashimai exhibits a remarkable sexual dimorphism, particularly in coloration. The pattern of sexual differences in H. hirashimai is similar to that observed in a Taiwanese species, H. triangularis ( Shinohara et al. 2020b) . In the two species, the female is a black xiphydriid with creamy white marking ( Fig. 2 View FIGURE 2 ; the legs are also black and creamy white) while the male is a creamy white or pale brown xiphydriid with black marking only dorsally ( Fig. 3 View FIGURE 3 ; the legs are entirely pale brown to creamy white), giving the female and male very different general appearances ( Figs 2 View FIGURE 2 , 3 View FIGURE 3 ). The inner tooth of the tarsal claws is situated near middle in the female and near apex in the male.
Large sexual differences in color pattern and shape of the tarsal claws were reported also for Platyxiphydria tiphiiformis Takeuchi, 1938 ( Shinohara & Matsumoto 2019), whereas the sexual dimorphism is less conspicuous in the genera Xiphydria ( Shinohara 2019a; Shinohara & Kameda 2019; Shinohara et al. 2020a), Konowia Brauns, 1884 ( Shinohara 2019b) and Lataxiphyda Smith, 2008 ( Shinohara 2019c) .
The genus Hyperxiphia is currently represented by 24 species ( Taeger et al. 2010; Shinohara 2019d; Liu et al. 2019; Li et al. 2019; Smith 2019, 2020). Of these, both sexes are known for five species, including H. hirashimai , while only females are known for 15 species and only males are known for four species. Almost all the species of Hyperxiphia are known from one or a few specimens each. Much more material of both sexes, particularly reared pairs of specimens or fresh material which can provide molecular data, is needed for correct association of sexes.
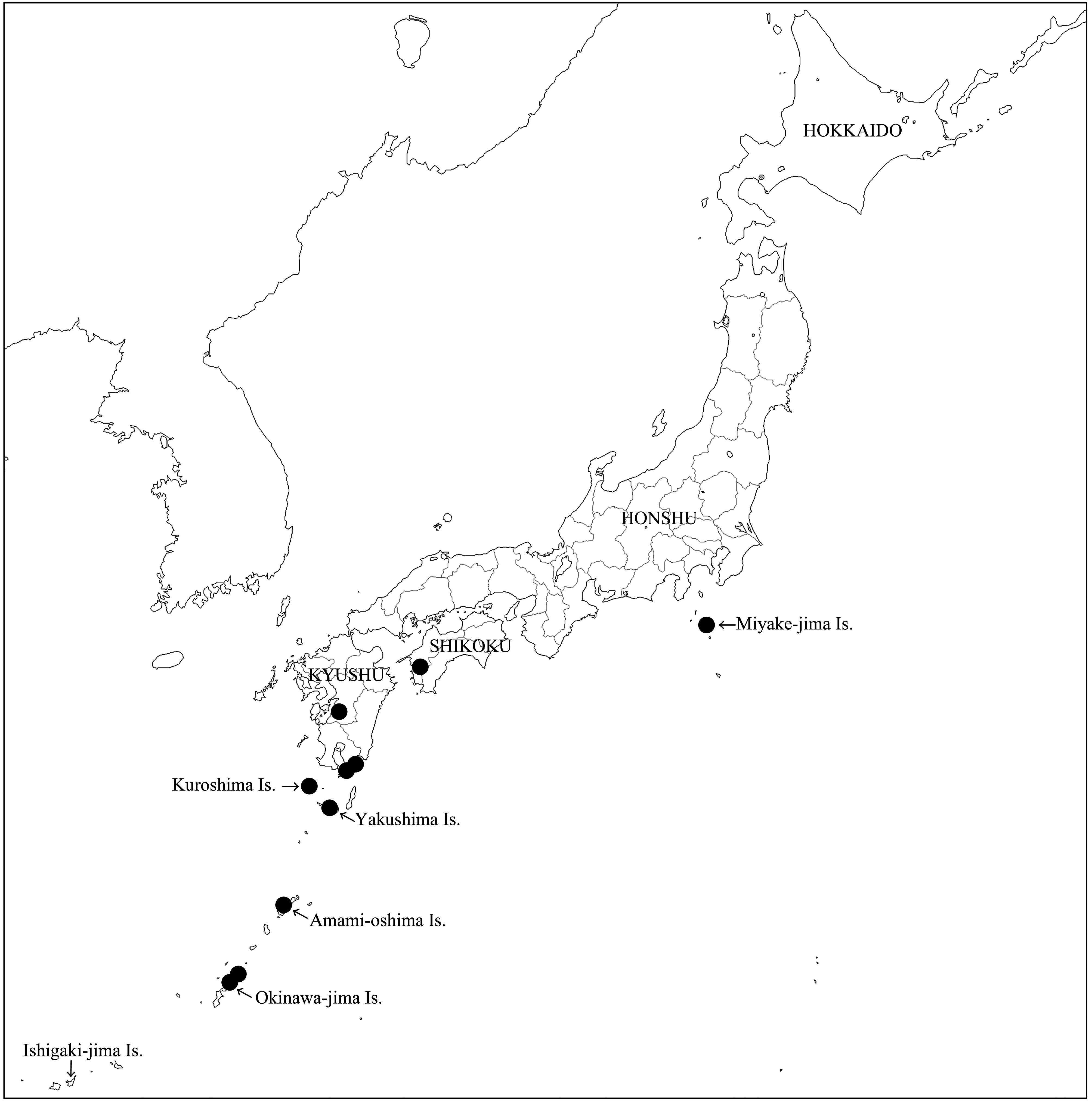
Distribution. This species was described from Amami-oshima Island ( Okutani 1965). We examined an additional nine females and six males, which reveal that H. hirashimai occurs widely from Miyake-jima Island, southwestern Shikoku, southern Kyushu, through Yakushima and Amami-oshima Islands to Okinawa-jima Island ( Fig. 5 View FIGURE 5 ). The seemingly strange isolated distribution of this species in Miyake-jima Island may be due to the paucity of the available material. The four available COI sequences from Yakushima, Kuroshima and Okinawa-jima Islands were the same, suggesting the absence of large genetic divergence among the populations of the three islands. Yakushima and Kuroshima Islands are about 70 km apart and these two islands are about 435 km and 470 km, respectively, away from Okinawa-jima Island. The peculiar southwest-northeast extending distribution range of this species may have been established, perhaps partly, by rafting of logs containing the larvae with the north-flowing Kuroshio Current running along the southeastern margin of the Japanese Archipelago. The possibility of transoceanic dispersal of xylophagous insects with sea current has been discussed by various authors (e.g., Ohbayashi & Niisato 2007, Makihara et al. 2015), though Holzapfel & Harrell (1968) noted “marine drift has probably been the least successful method of insect dispersal”.
Based on both morphological and molecular data (see above), H. hirashimai is regarded as very close to H. triangularis from Taiwan. It is interesting that these two species are allopatric in distribution and the ranges of the two species are widely separated by a “vacant” area including such comparatively large islands as Miyako-jima, Ishigaki-jima and Iriomote-jima Islands, which have rich representation of plants and animals. The minimum distance between Taiwan and Okinawa-jima Island is about 600km. We need more material and information for understanding the actual distribution range of H. hirashimai and for having a plausible explanation for this peculiar distribution pattern.



No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Hyperxiphia hirashimai ( Okutani, 1965 )
| Shinohara, Akihiko & Yamasako, Junsuke 2020 |
Genaxiphia hirashimai
| Shinohara, A. 2019: 21 |
| Taeger, A. & Blank, S. M. & Liston, A. D. 2010: 113 |
| Abe, M. & Togashi, I. 1989: 559 |
| Smith, D. R. 1978: 105 |
| Okutani, T. 1965: 74 |