
Chamobates callipygis Pavlichenko, 1991
|
publication ID |
https://doi.org/ 10.11646/zootaxa.5115.1.6 |
|
publication LSID |
lsid:zoobank.org:pub:12D7CC63-3143-4F37-B59D-CBC5E5F065ED |
|
DOI |
https://doi.org/10.5281/zenodo.6347048 |
|
persistent identifier |
https://treatment.plazi.org/id/44312216-0076-FFD5-FF71-F894476EFEEA |
|
treatment provided by |
Plazi |
|
scientific name |
Chamobates callipygis Pavlichenko, 1991 |
| status |
|
Chamobates callipygis Pavlichenko, 1991 View in CoL
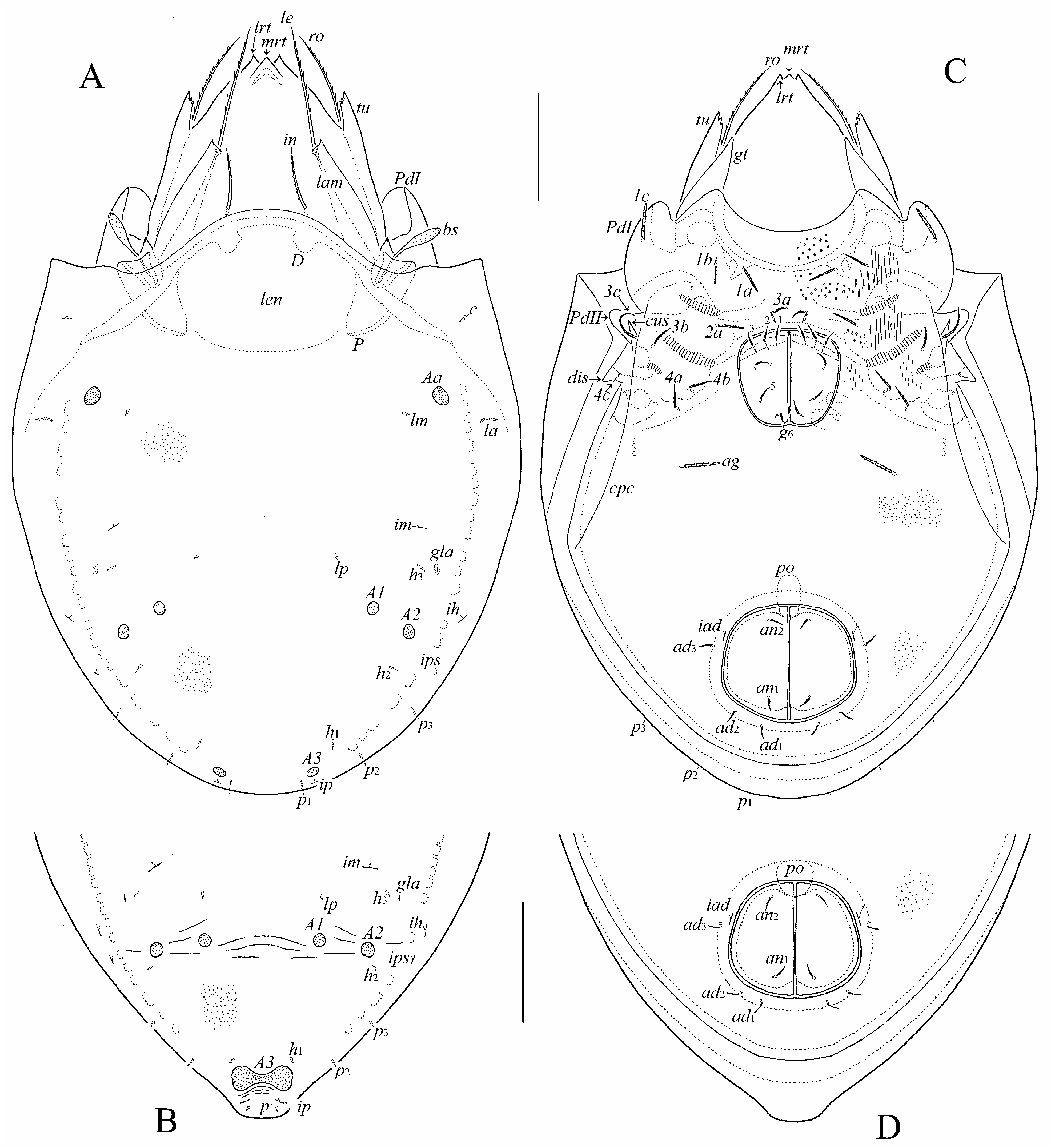
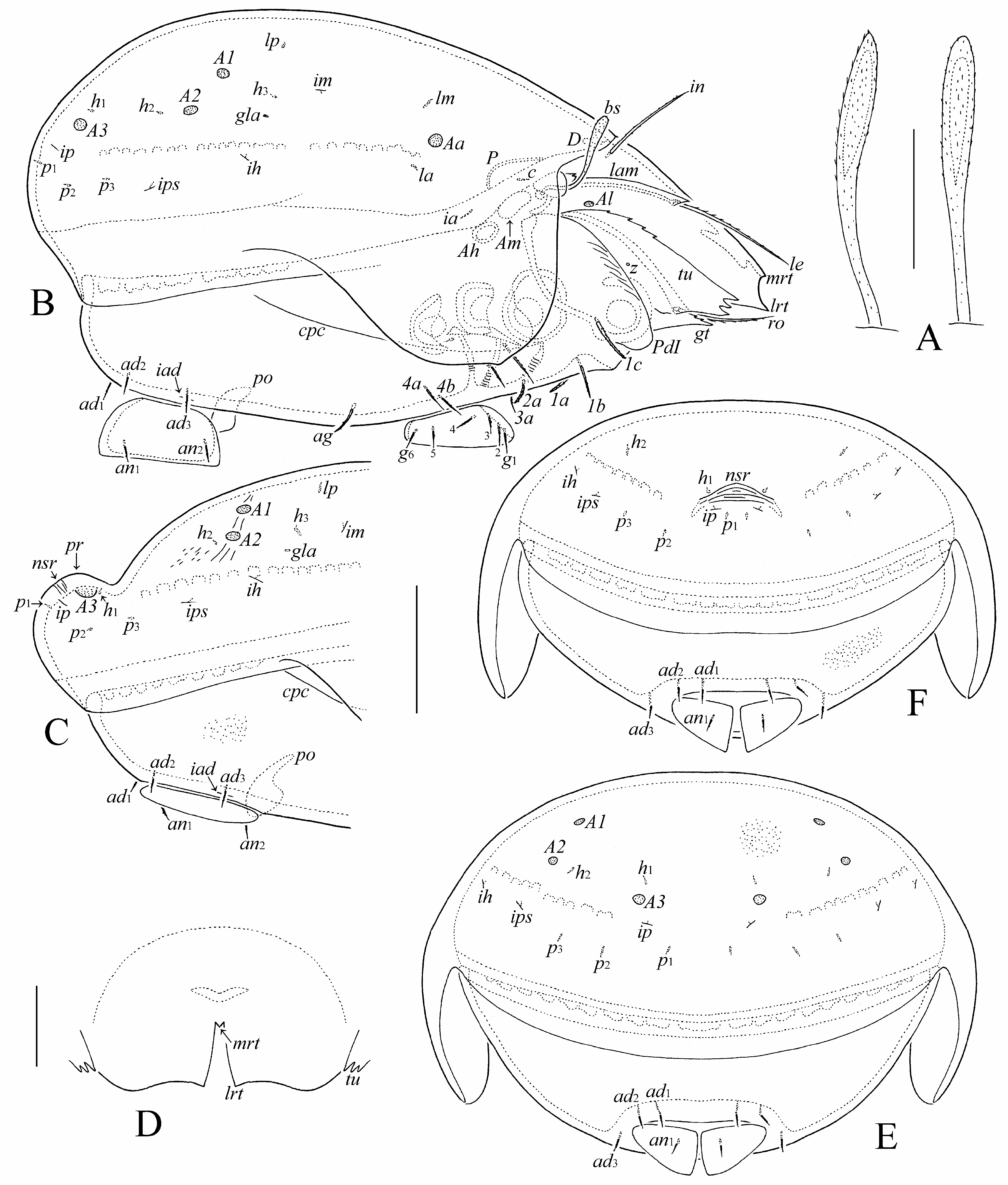
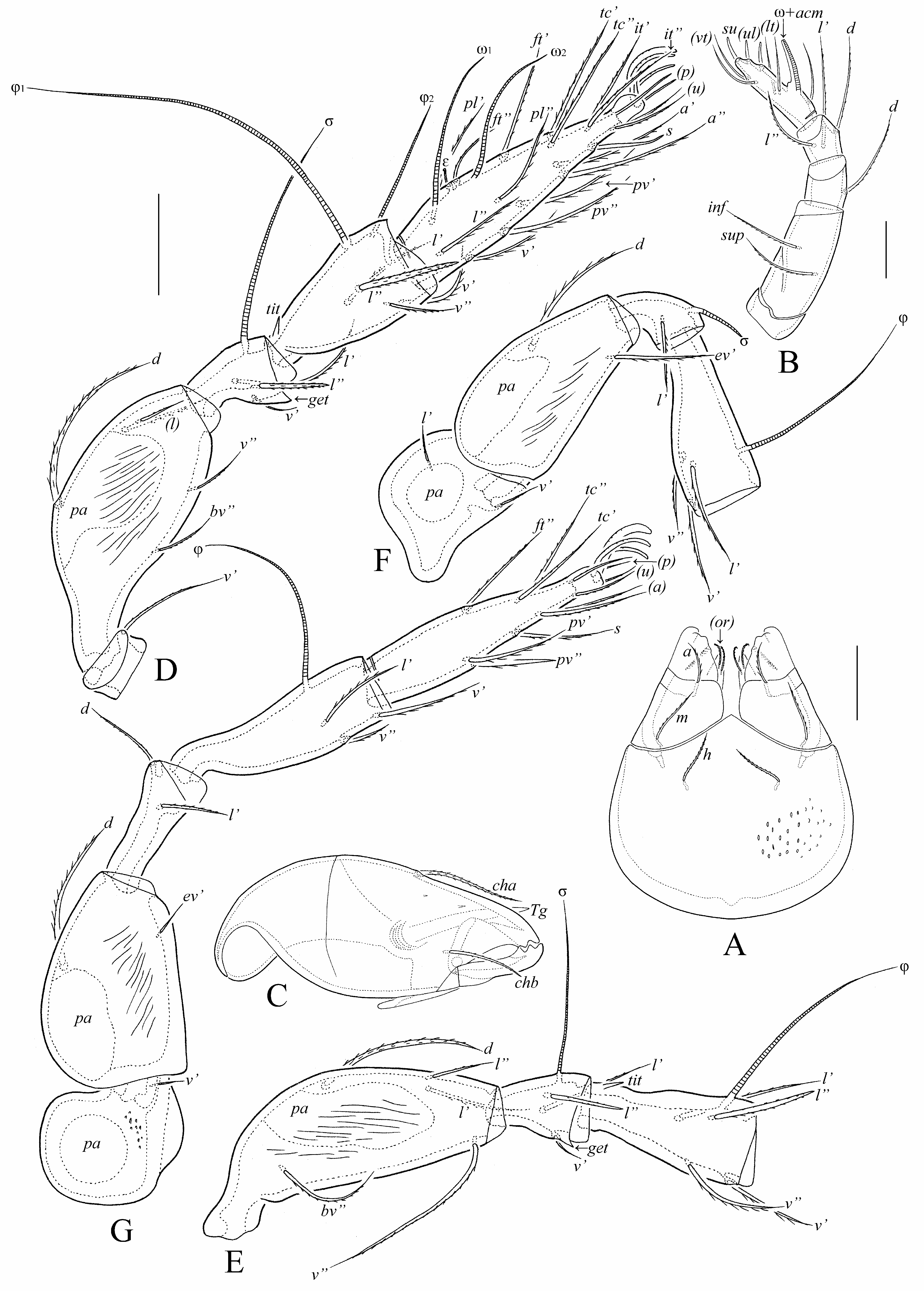
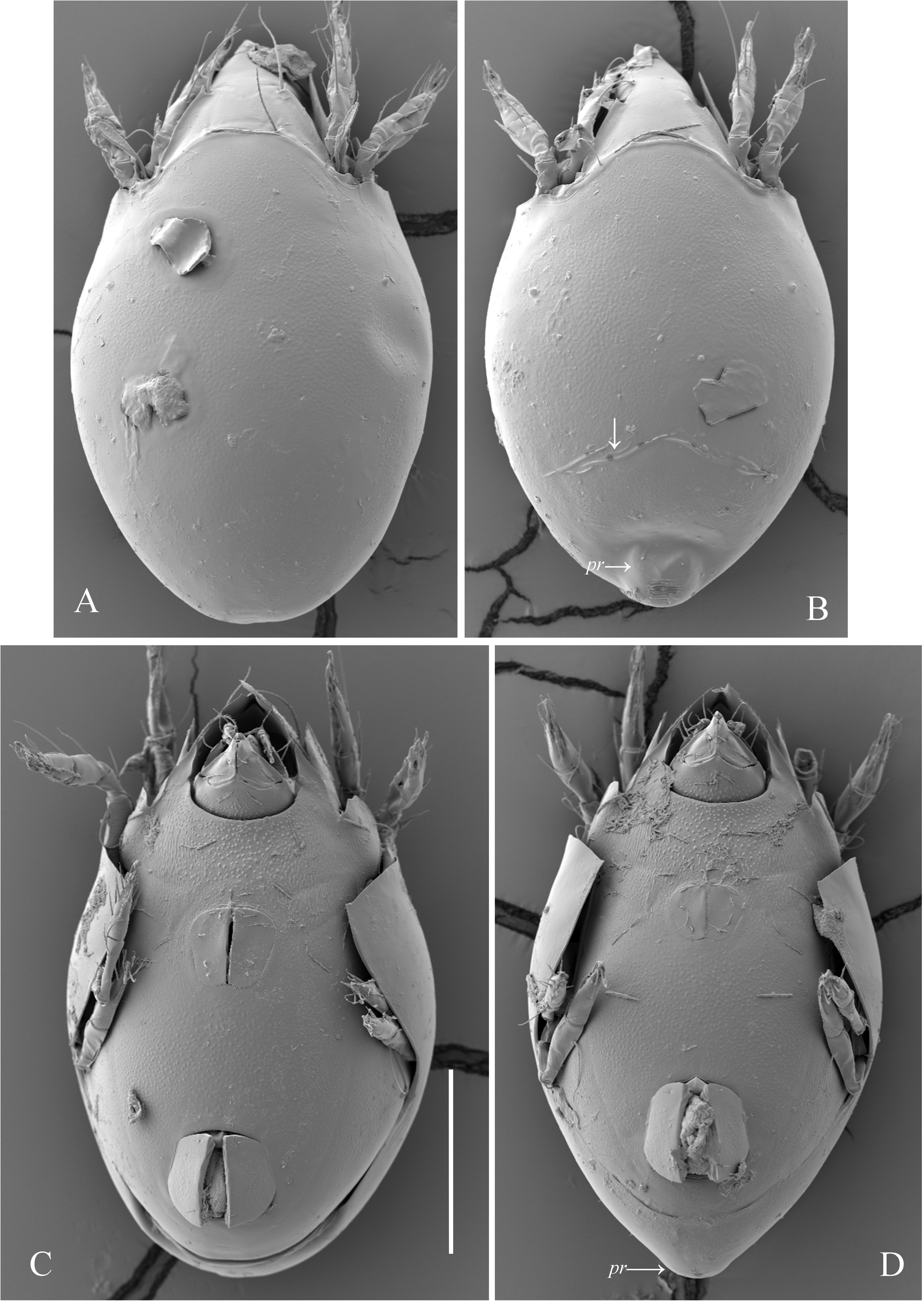
( Figs 1–6 View FIGURE 1 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 )
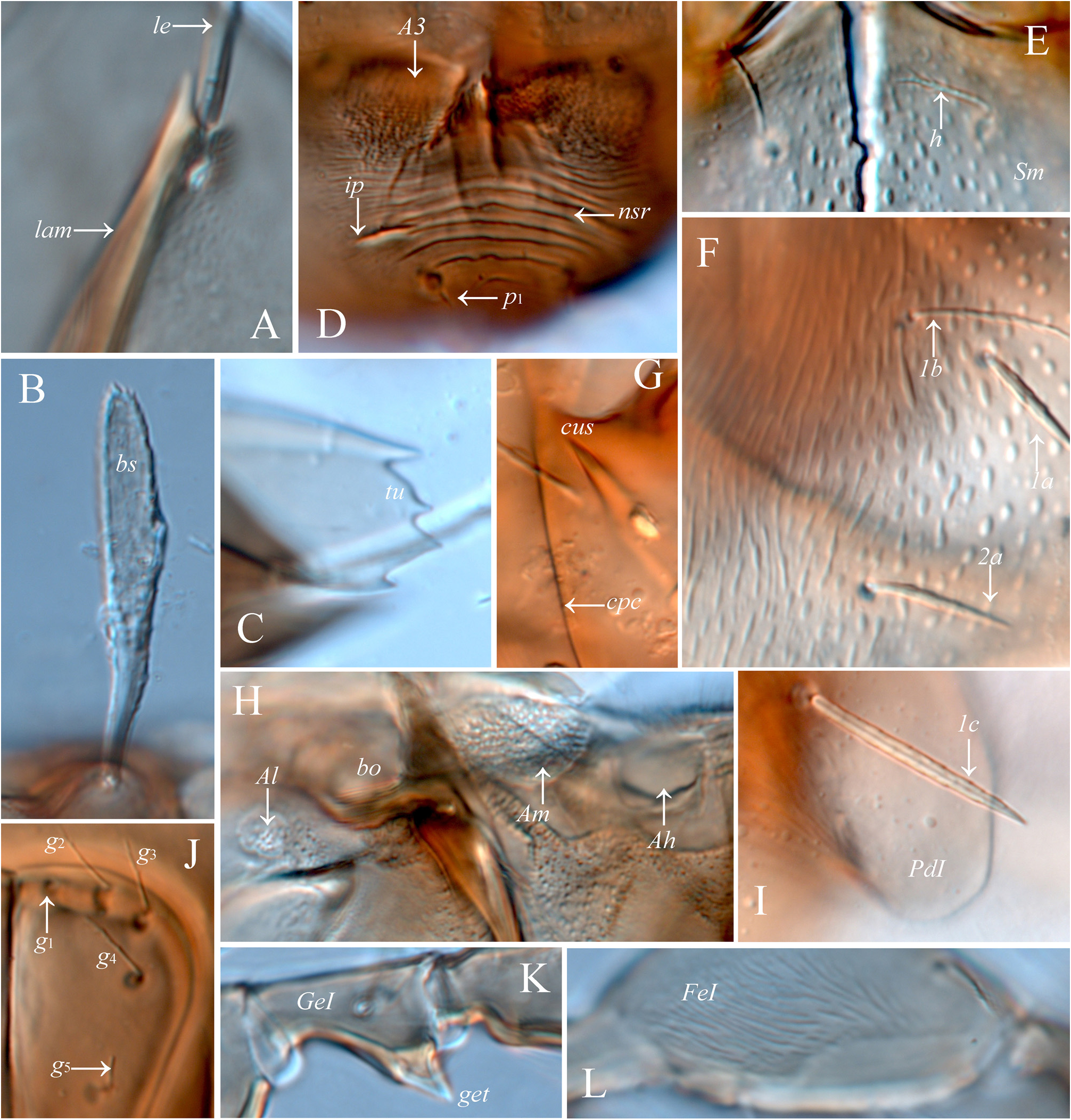
Diagnosis. Adult. Body size: 282–332 × 182–215. With sexual dimorphism: in males, posterior part of notogaster with some striations near porose areas A1, A2 and large posteromedian protuberance bearing pair of fused porose areas A3, and densely striate region behind them (females without protuberance and striations on notogaster, and with four pairs of rounded porose areas). Epimeral region granulate and longitudinally striate. Rostrum with strong lateral teeth and one small tooth at base of deep indentation between lateral teeth. Tutorial cusp with three or four teeth. Genal tooth sharply pointed, with some curved integumental ridges proximally. Rostral, lamellar and interlamellar setae setiform, barbed; le longest. Bothridial seta well developed, flattened, rounded or pointed apically, barbed. Anterior lamina of bothridium with two small teeth. Pteromorph slightly pointed laterally. Notogastral setae simple, minute. Epimeral seta 1c bacilliform; 3a thin, slightly stiff; 3c setiform; 4c simple, thinnest; 1a, 1b, 2a, 3b, 4a, 4b thickened. Genital, anal and adanal setae short, slightly stiff; aggenital seta bacilliform. Leg tibiae I and II with dorsoproximal tooth.
Adult. Measurements. Body length: 282–332 (eight males and eight females); notogaster width: 182–215 (eight males and eight females). No clear difference between males and females in body size.
Integument. Body color light brown. Body surface and legs densely microsculptured. Subcapitular mentum granulate, epimeral region granulate and longitudinally striate, pedotectum I dorsally slightly striate, all leg femora striate antiaxially.In male, posterior part of notogaster sparsely transversely striate near porose areas A1, A2 and densely transversely striate on posteromedian protuberance, behind porose areas A3 (in female, striate notogastral regions absent).
Prodorsum ( Figs 1A View FIGURE 1 , 2A, 2B, 2D View FIGURE 2 , 4A–C View FIGURE 4 , 5A, 5E, 5G, 5H View FIGURE 5 , 6A–C, 6H View FIGURE 6 ). Rostrum tridentate; lateral teeth strong, indentation between them deep; very small median tooth located at base of indentation and hanging over indentation in dorsal view ( Fig. 2D View FIGURE 2 ). Lamella about 1/2 length of prodorsum, distally with strong lateral tooth. Tutorium (including cusp) about 4/5 of length of prodorsum, with some teeth dorsally; cusp broadly quadrangular, distally with three (rarely, with four) teeth. Genal tooth triangular, sharply pointed, with some curved integumental ridges proximally ( Fig. 5E View FIGURE 5 ). Lateral porose area Al present, hardly observed, oval. Rostral (41–49), lamellar (57–65) and interlamellar (36–45) setae setiform, barbed. Bothridial seta (36–45) with flattened, rounded or pointed apically, barbed head. Anterior lamina of bothridium with two small teeth. Exobothridial seta and dorsosejugal porose area not observed.
Notogaster ( Figs 1A, 1B View FIGURE 1 , 2B, 2C, 2E, 2F View FIGURE 2 , 4A–C View FIGURE 4 , 5A–D View FIGURE 5 , 6D View FIGURE 6 ). Anterior notogastral margin convex medially. In male, notogaster with posteromedian protuberance (in female, protuberance absent). Pteromorph slightly pointed laterally. Lenticulus present, with diffuse border. Four pairs of rounded porose areas in females (Aa: 6–12; A1, A2: 6–8; A3: 6–12); in males, Aa, A1, A2 as for female, A3 fused medially forming dumbbell-like structure, i.e., two rounded nodes connected centrally by a beam. All notogastral setae (2) setiform, thin, smooth. Opisthonotal gland opening and all lyrifissures distinct.
Gnathosoma ( Figs 3A–C View FIGURE 3 , 4D, 4E View FIGURE 4 , 5E View FIGURE 5 , 6E View FIGURE 6 ). Subcapitulum size: 77–82 × 61–65. Mentum without lateral apophysis. Subcapitular (a: 14–16; m, h: 18–20) and adoral (8–10) setae setiform, barbed. Palp (57–61) with typical setation (0-2-1-3-9+ω). Postpalpal seta (4) spiniform, smooth. Axillary saccule distinct, elongate. Chelicera (90–94) with two setiform, barbed setae (cha: 28–32; chb: 20).
Epimeral and lateral podosomal regions ( Figs 1C View FIGURE 1 , 2A View FIGURE 2 , 4D, 4E View FIGURE 4 , 5F View FIGURE 5 , 6F–I View FIGURE 6 ). Epimeral setal formula: 3-1-3-3. Seta 1c (20–28) bacilliform, barbed; 3a (8–10) thin, slightly stiff, barbed, pointed apically; 3c (20–24) setiform, barbed; 4c (4) setiform, thin, roughened; 1a, 1b, 2a, 3b, 4a, 4b (14–16) thickened, barbed, pointed apically (1a, 2a thicker than 1b, 3b, 4a, 4b). Humeral porose area Am elongate oval; Ah oval, distinct. Custodium narrowly tooth-like. Discidium triangular. Circumpedal carina long, reaching level of pedotectum II, not connected with custodium.
Anogenital region ( Figs 1C, 1D View FIGURE 1 , 2A, 2C, 2E, 2F View FIGURE 2 , 4D, 4E View FIGURE 4 , 5F, 5I View FIGURE 5 , 6J View FIGURE 6 ). Genital, anal and adanal setae (8–10) thin, slightly stiff, barbed, pointed apically. Aggenital seta (16–24) bacilliform, barbed. Adanal lyrifissure located close and parallel to anal plate. Preanal organ caecum-like. Postanal porose area absent.
Legs ( Figs 3D–G View FIGURE 3 , 6K, 6L View FIGURE 6 ). Median claw distinctly thicker than lateral claws, all slightly barbed dorsally. Porose area present dorsoparaxially on femora I–IV and on trochanters III, IV versus absent proximoventrally on all tarsi and ventrodistally on all tibiae. Tibiae I and II with proximodorsal tooth ( Fig. 3D, 3E View FIGURE 3 ). Genua I and II with triangular ventrodistal process (sometimes hardly observable on genu II). Formulas of leg setation and solenidia: I (1-5-3-4-20) [1-2-2], II (1-5-3-4-15) [1-1-2], III (2-2-2-3-15) [1-1-0], IV (1-2-2-3-12) [0-1-0]; homology of setae and solenidia as indicated in Table 1 View TABLE 1 . Famulus short, erect, slightly swollen distally, inserted between seta ft” and solenidion ω 2. Seta s on tarsus I eupathidial, located before setae a’ and a”. Seta l” on tibiae I, II and genua I, II thickened. Solenidia ω 1 and ω 2 on tarsus II and σ on genu III bacilliform; other solenidia setiform.
Remarks. Generally, the original description of males by Pavlichenko (1991) was adequate except for the following traits: three teeth on rostrum; all notogastral setae and epimeral seta 4c represented by alveoli; 18 setae on leg tarsus I. We have found rostrum with strong lateral teeth, indentation between them and additional very small median tooth in the base of indentation, but tooth in the base of indentation hangs over the indentation in dorsal view creating illusion of tridentate rostrum; notogastral setae and epimeral seta 4c developed, but minute; 20 setae present on leg tarsus I.
Despite the absence of molecular analysis and without checking the type material, we have no doubt that the males and females described herein belong to C. callipygis . This can be confirmed by the similar structure of the rostrum and bothridial setae; the presence of unusual tooth in the dorsobasal part of tibiae I and II; the presence of a dentate tutorium (dorsally and apically); the presence of the curved integumental ridges proximally to the genal tooth; the presence of two unusual teeth in front of bothridium; and especially the specific sculpture of the epimeral region and structure of the epimeral setae, in both sexes. Additionally, all our specimens were collected from the same habitat (in one sample). Our studied material was collected in Crimea, from where this species was originally described, and probably, is an endemic to this region as it has not been found elsewhere .
It is worth to note that according to Pavlichenko (1991), the type material of this species is at the I. I. Schmalhausen Institute of Zoology, National Academy of Sciences of Ukraine. However, we did not examine the type material because the original description was quite sufficient to identify this species .

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |