
XENODONTINAE, Bonaparte, 1845
|
publication ID |
https://doi.org/ 10.1206/3715.2 |
|
persistent identifier |
https://treatment.plazi.org/id/4548181D-D610-2817-1D19-FC8CE0DDFB74 |
|
treatment provided by |
Carolina |
|
scientific name |
XENODONTINAE |
| status |
|
“ XENODONTINAE ”
More than a quarter century ago investigators began to realize that New World “xenodontine” snakes comprise two geographically overlapping clades—the Central American “xenodontines” and the South American “xenodontines”—as documented primarily by microcomplement fixation studies of serum albumins ( Dowling et al., 1983; Cadle, 1984a, 1984b, 1984c, 1985); for summary discussion see Zaher (1999: 5–7).
The predominantly Central American assemblage was recognized as subfamily Dipsadinae by Dowling et al. (1983: 323, 326). Zaher’s (1999: 33) concept of Dipsadinae corresponded “essentially to Cadle’s Central American group,” as supported morphologically by the hemipenial characterization provided by Myers and Cadle (1994: 27). Zaher stated that one of the three synapomorphies suggested by Myers and Cadle—distal division of the sulcus spermaticus within or at the base of the capitulum—“is nearly unique to the dipsadines, being otherwise present only in the Madagascan genus Geodipsas .”
9 The “Hiss drawing” of a hemipenis labeled “ Hydraethiops melanogaster ” in Dowling and Duellman (“1974– 1978” [1978]: fig. 112d.4) is an accidentally transposed illustration of Natriciteres olivacea —apparently the same hemipenis represented by the photograph of AMNH 11915 in Zaher (1999: fig. 19, upper). The adjacent drawing (fig. 112d.5) mislabeled “ Natriciteres ” probably is Hydraethiops melanogaster , but not the same organ illustrated under that name in Zaher (1999: fig. 18, lower).
N.B.: The hemipenis of Natriciteres is not a unilobed organ like that of Limnophis as implied by Zaher (1999: 32). The aforesaid illustrations of N. olivacea show what appears to be a bifurcated organ with the right lobe very incompletely everted, but, as described and illustrated by Bogert (1940: 33 --35 [as Neusterophis ]), the Natriciteres hemipenis is only basally bifid, with one long and one very short lobe controlled by unequal-length slips of retractor muscle. Thus, the shortening leading to loss of a single lobe is one method of becoming unilobed, the alternative being distal shortening of both lobes simultaneously.
10 The flaplike structure shown on the left lobe in Zaher’s (1999: fig. 18) photograph of Hydraethiops melanogaster (AMNH 11959) is an artifact (torn tissue) of preparation.
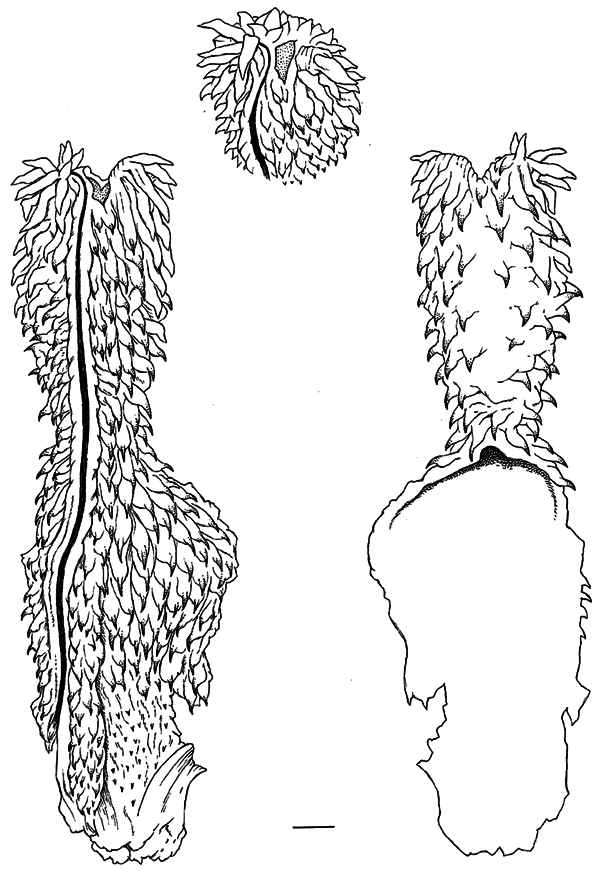
The restricted subfamily Xenodontinae was treated in considerable detail by Zaher (1999), who provided an exhaustive survey of xenodontine (and other) hemipenes and suggested synapomorphies for defining the clade. Owing to the lack of field-everted hemipenes, Zaher prepared and illustrated numerous hemipenes that he everted manually. Such eversions elucidate features often not interpretable from dissections of retracted organs (both kinds of data are essential for complete understanding). Limitations to manual eversion of fully retracted organs of museum specimens include (1) some necessary damage will occur to the base of the hemipenis and (2) complete expansion often cannot be attained (resulting in a smaller circumference), even though eversion may be complete with all structures revealed. Both situations apply to the manually everted hemipenis of Amnesteophis melanauchen (see p. 10 and fig. 5 View FIG ). The techniques and limitations of manually everting hemipenes were discussed by Myers and Cadle (2003) and Zaher and Prudente (2003).
Dowling (2004) seemed unenthused about the usefulness of manual eversions and responded critically to the last two papers and also to Zaher’s (1999) earlier study. Based on “prior knowledge” gained from having “prepared more than 400 ‘fresh’ hemipenes in the last half-century,” and “due to long experience and careful observation,” Dowling was able to provide imaginative explanations for hemipenial features with which he had had no experience. Two examples: he suggested ( Dowling, 2004: 321) that “the disappearing calyces” (the “pseudocalyces” of Myers and Cadle) on an everted Psomophis hemipenis were “an artifact of incomplete specimen preparation” (i.e., not injecting the “outer lymph sinus”), and he predicted (ibid.: 325) how the everted hemipenes of Xenopholis really should look after stating that five of Zaher’s illustrations were “so inaccurate as to place the taxa in the wrong family.” 11 Dowling (2004: 325–327) particularly critiqued Zaher’s (1999) photographs of manually everted hemipenes by “doing a page-by-page survey of [Zaher’s] entire study” and listed those illustrations he judged to be based on distorted or incompletely everted organs. However, few of the supposed “incomplete” eversions or “distortions” mask the synapomorphic structures that Zaher discussed, nor was Dowling’s critique as thorough as he implied. Witness that Zaher’s study (1999: 154–156) included photographs of hemipenes of three species of Xenodon , and yet Dowling (2004: 326) asserted that “it seems strange that Zaher failed to illustrate the hemipenis of the type genus of xenodontine snakes ( Xenodon ) (v. my Fig. 11 [= Dowling’s own accompanying illustration])”!
11 Concerning pseudocalyces: Normal calyces have walls of varying flexibility but do not disappear no matter how the organ is everted or to what extent it is inflated. As noted by Myers and Cadle (1994: 13), retracted Psomophis hemipenes have obliquely transverse folds of large spinulate papillae that are connected longitudinally by fine tissue folds, which bear striking resemblance to calyces “when the inverted lobe is incised and gently parted.” The connecting longitudinal folds, however, “can be flattened by micromanipulation with fine teasing needles.” The calyxlike structures were absent on the only available field-everted organ and were judged to have disappeared during inflation owing to stretching of the fine connecting folds (Myers and Cadle, 1994: 10–13). Two manually everted Psomophis hemipenes are fully everted but not completely expanded. The transverse papillate folds are evident in photographs (Zaher, 1999: fig. 74), but the connecting longitudinal folds show only at high magnification as hairline folds that “probably would disappear in a maximally expanded fresh eversion” (Myers and Cadle, 2003: 296, fig. 3 View FIG ).
Concerning Xenopholis: The appearance of the hemipenis of X. scalaris illustrated by Zaher (1999: 168, fig. 95) compares favorably with one that I field-everted in Rondônia, Brazil (AMNH R-130238).
Zaher et al. (2009: 141) have reworked the classification of “ Xenodontinae ,” returning it to earlier concepts of a single group of New World “xenodontines” that (because of nomenclatural priority) would now have to be called “dipsadines.” Their new groupings follow:
Family Dipsadidae Bonaparte, 1838
Subfamily Carphophiinae Zaher et al., 2009
Subfamily Dipsadinae Bonaparte, 1838
Subfamily Xenodontinae Bonaparte, 1845 (with 14 tribes)
Subfamily Xenodontinae was now said to have “No known synapomorphies” because synapomorphies presented by Zaher (1999) were moved to family Dipsadidae for complex and unresolved issues explained by Zaher et al. (2009: 140). Nonetheless, subfamily Xenodontinae was retained (appropriately in my view) to avoid changing “the well-established taxonomic hierarchy for this group” (Zaher et al., 2009: 141–142).
I have no reason to challenge the phylogeny that led to this classification except to note that ongoing DNA sampling still is insufficient for a thoroughly robust testing of these groupings (cf. Vidal et al., 2000; Kelly et al., 2003; Lawson et al., 2005: 594; Hedges et al., 2009; Zaher et al., 2009). Therefore the Dipsadinae and Xenodontinae are retained herein for practical reasons as subfamilies of Colubridae sensu lato for reasons discussed above (introductory paragraphs under Comparisons and Higher-Level Taxonomy).
Herndon Dowling has been a long-time proponent of assigning colubrid snake genera to tribes (e.g., see Dowling and Duellman, “1974–1978” [1978]). Zaher et al. (2009) have significantly advanced the process by distributing the colubrid subfamily Xenodontinae among 14 tribes, which facilitates discussion and further research. Tribal designations and molecular sampling are much less complete for the Dipsadinae .
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.