
Pristimantis mazar Guayasamin & Arteaga
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3616.4.3 |
|
publication LSID |
lsid:zoobank.org:pub:5FA7195E-1152-4F3C-8860-374A1EAAEF62 |
|
DOI |
https://doi.org/10.5281/zenodo.6154437 |
|
persistent identifier |
https://treatment.plazi.org/id/4663490E-FFFB-3D30-CBE6-0AEDEF09FE1B |
|
treatment provided by |
Plazi |
|
scientific name |
Pristimantis mazar Guayasamin & Arteaga |
| status |
sp. nov. |
Pristimantis mazar Guayasamin & Arteaga , new species
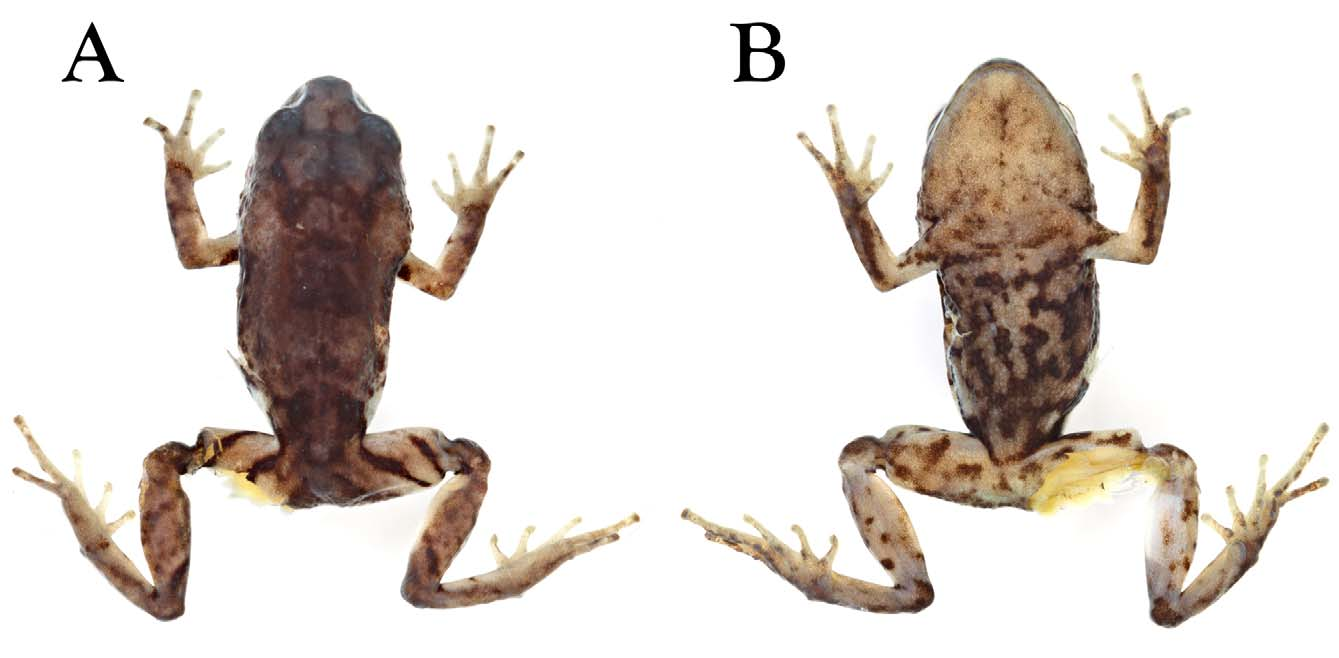
Holotype. QCAZ 27556 (SC 12660), an adult female ( Figs. 1 View FIGURE 1 , 2 View FIGURE 2 ) obtained by Joe Mendelson, Michelle Cummer, and Martín Bustamante on February 2004, at La Libertad, Reserva Mazar (02.546589º S, 78.698375º W; 2895 m.a.s.l.), Cantón Azogues, Provincia Cañar, Ecuador; airline distance from Rivera is 6.6 km.
Paratypes. Paratypes QCAZ 27553–54 have the same collecting data as the holotype. Paratypes QCAZ 27493, 27503–05, 27507–08, 27511, 27514, 27519, 27536, 27555, 27560 were collected by Joe Mendelson, Michelle Cummer, and Martín Bustamante on February 2004, at Rumiloma, Reserva Mazar (02.574616º S, 78.745462º W; 3400 m.a.s.l.), Cantón Azogues, Provincia Cañar, Ecuador. Paratypes QCAZ 29563, 29565, 32619, 49750, 49764 were obtained either by Alejandro Arteaga or Martín Bustamante at the type locality in following years. From all these, ten are adult males (QCAZ 27493, 27504–05, 27507, 27519, 27553–54, 29563, 29565, 49764) and nine adult females (QCAZ 27503, 27508, 27511, 27514, 27536, 27555, 27560, 32619, 49750).
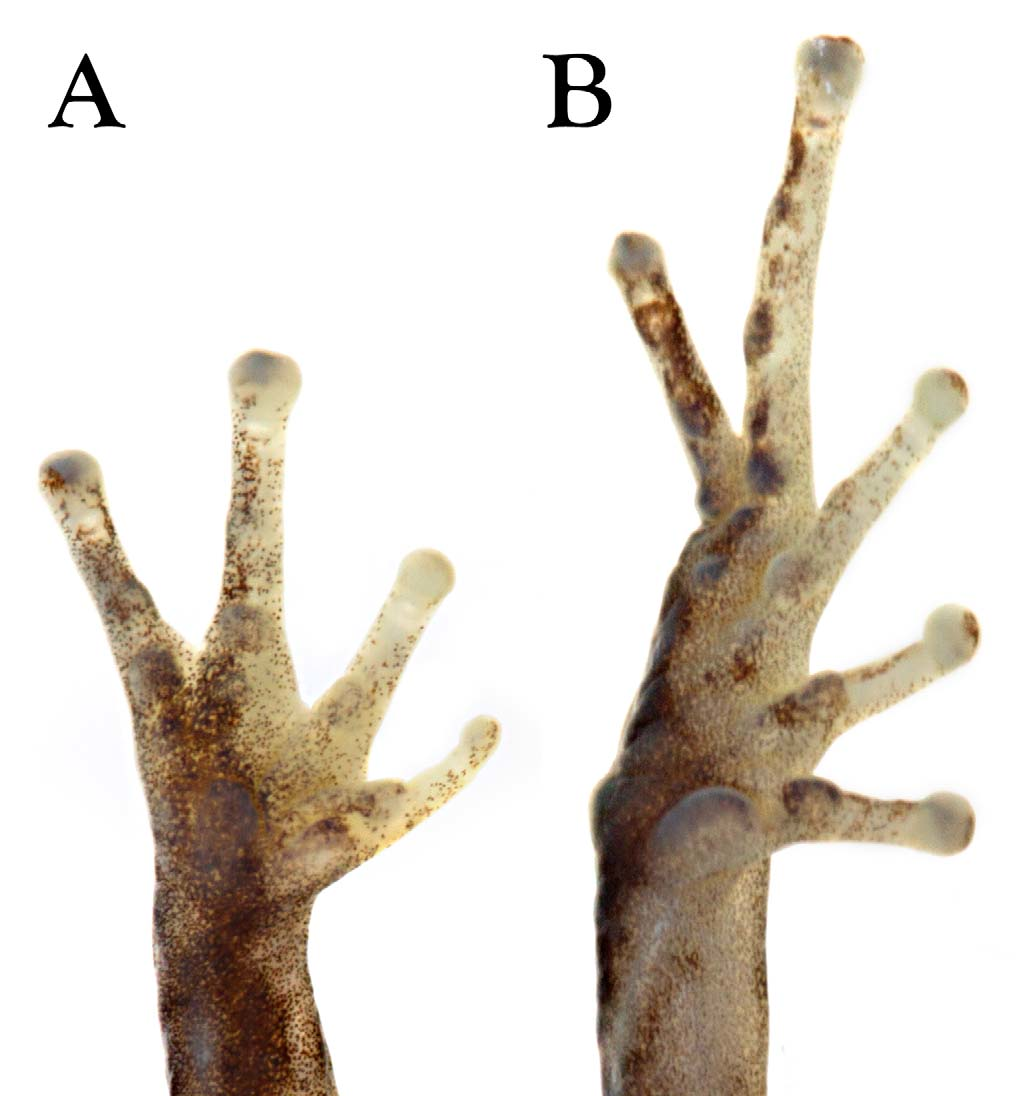
Diagnosis. The new species is placed in the genus Pristimatis, as diagnosed by Hedges et al. (2008), because of the presence of the following traits: cranial crests absent, dentigerous process of the vomers present, and Tshaped terminal phalanges. Pristimantis mazar is diagnosed by having: (1) skin texture of dorsum shagreen with few widely interspersed low tubercles, which became more abundant on flanks and posterior end of dorsum; occipital and dorsolateral folds low or absent; venter areolate; (2) tympanic membrane and tympanic annulus present, with upper rim obscured by supratympanic fold; (3) snout short, pointed in dorsal view and rounded in profile; (4) upper eyelid lacking enlarged tubercles; tubercles are low and almost undistinguishable from surrounding skin texture; cranial crests absent; (5) dentigerous process of vomers well developed, oblique in outline, positioned posterior to level of choanae and separated medially by distance less than width of odontophore, each process bearing 0–7 teeth in females and 0–3 teeth in males; (6) males with a small subgular vocal sac; vocal slits present; nuptial pads absent; (7) first finger shorter than the second; discs on fingers rounded, slightly expanded ( Fig. 2 View FIGURE 2 ); (8) fingers bearing narrow lateral fringes; palmar tubercle bifurcated distally; supernumerary tubercles round, fleshy ( Fig. 2 View FIGURE 2 ); (9) ulnar tubercles low or absent in females, present and low in males; inner tarsal fold present, short; (10) heel and tarsal tubercles low or absent in females, present and low in males; (11) toes bearing narrow fringes; webbing absent; Toe V slightly longer than Toe III; toe discs rounded and slightly expanded ( Fig. 2 View FIGURE 2 ); (12) inner metatarsal tubercle elliptical, about 1.4–2 times the size of outer, rounded metatarsal tubercle; supernumerary plantar tubercles round and fleshy ( Fig. 2 View FIGURE 2 ); (13) in ethanol, dorsum creamish gray to dark brownish gray, with or without darker marks and small white spots; lower flanks with a black-and-white pattern; venter from uniformly whitish cream to cream with darker marking that may form a reticulated pattern. In life, dorsum brown to reddish brown, with or without small white dots and dark brown marks; black spots on groin; flanks and venter cream to gray, usually with blackish mottling that may form a reticulated pattern; iris cream to pale brown, with reddish brown to orange horizontal streak; (14) small SVL in females 19.9–23.7 (mean = 21.5 ± 1.18, n = 10), in males 14.9–18.1 mm (mean = 16.6 ± 1.03, n = 10).
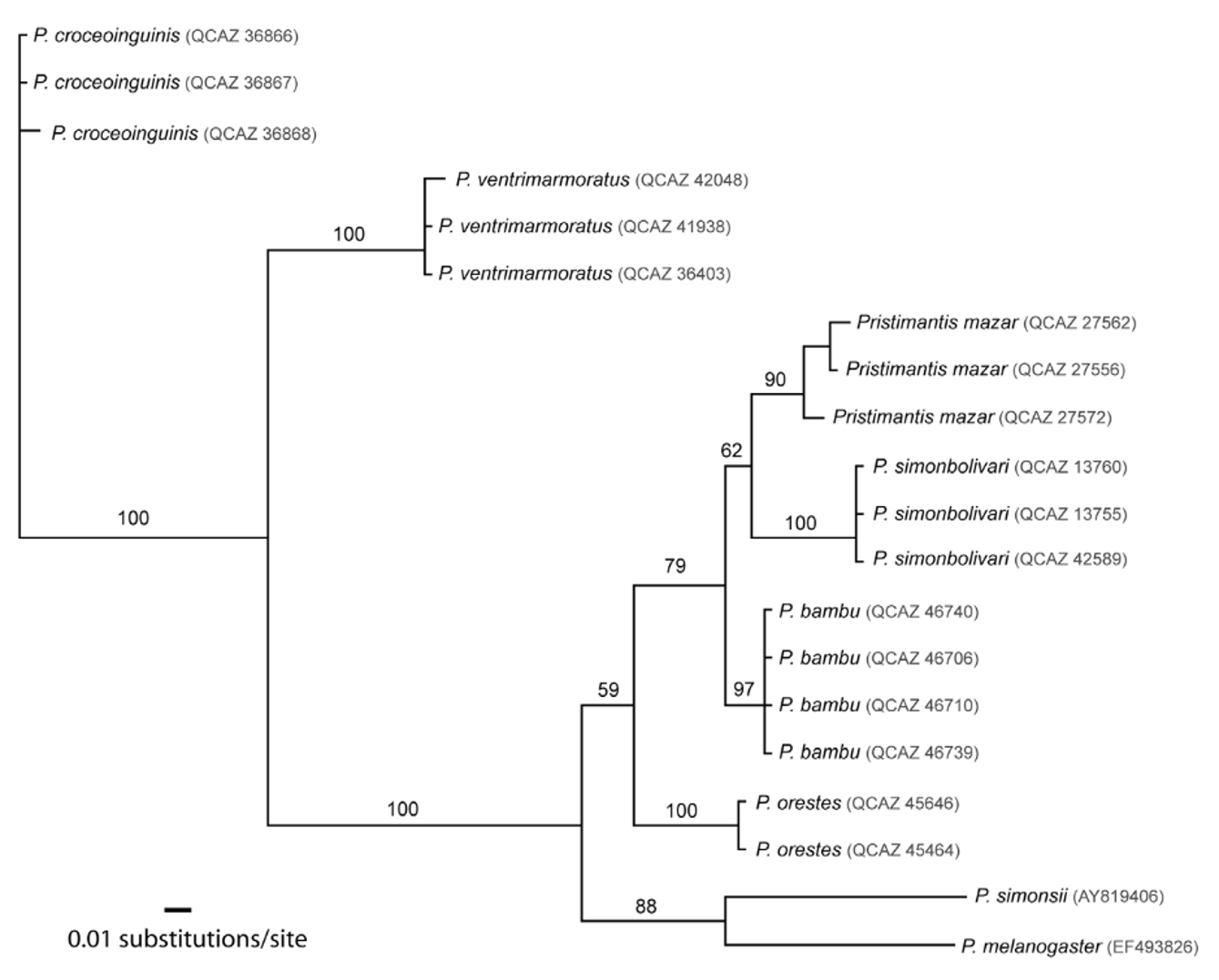
Similar species. The smaller, non-overlapping SVL of adult males of Pristimantis mazar readily distinguishes them from most species of the P. o re s te s group ( P. atrabracus , P. chimu , P. cordovae , P. corrugatus , P. melanogaster , P. orestes , P. pataikos , P. pinguis , P. simonsii , P. stictoboubonus ; Table 1). Similarly, adult females of P. m a z a r are smaller than females of P. bambu , P. corrugatus , P. mariaelenae , P. melanogaster , P. pinguis , P. simonsii , P. stictoboubonus , P. stipe , and P. ventriguttatus (Table 1). The new species differs from P. v i d u a by its ventral color pattern (usually with a reticulated pattern in P. mazar ; with minute brown flecking in P. v i d u a), skin texture (dorsolateral folds present in P. v i d u a and usually absent in P. m a z a r), and tympanum (visible in P. m a z a r, concealed beneath skin in P. v i d u a). The call of the new species has a higher dominant frequency (3122–3171 Hz in P. mazar ; 2500–2560 Hz in P. bambu ) and is longer (0.14– 0.022 s in P. mazar ; 0.005– 0.006 s in P. b a m b u) than the call of Pristimantis bambu . Finally, P. m a z a r differs from its sister species mostly by having a reticulated venter ( Fig. 3 View FIGURE 3 ; uniform in males and females of P. simonbolivari ), notorious black stripes on flanks ( Fig. 3 View FIGURE 3 ; uniform dark coloration in P. simonbolivari ), and by inhabiting on the Cordillera Oriental of the Ecuadorian Andes ( P. simonbolivari is restricted to one locality in the Cordillera Occidental). The two sister species are reciprocally monophyletic ( Fig. 7 View FIGURE 7 ).
Description of the holotype. Adult female (QCAZ 27556; Figs. 1 View FIGURE 1 , 2 View FIGURE 2 ). Head as long as wide, slightly wider than body; upper eyelid bearing few low tubercles; head width 38.5% of SVL; head length 39% of SVL; snout relatively short (snout to eye distance 15% of SVL), rounded in dorsal and lateral views; tongue longer than wide, posterior half notched and not adherent to floor of mouth; eye diameter larger than eye–nostril distance; nostrils not protuberant, directed anterolaterally; canthus rostralis weakly concave in profile; loreal region slightly concave; upper eyelid width 68% of interorbital distance; cranial crests absent; tympanic annulus distinct, except for upper border, which is obscured by supratympanic fold; tympanic membrane distinct; several low postrictal tubercles situated posteroventrally to tympanic annulus; choanae round, not concealed by palatal shelf of maxillary; vomerine odontophores posteromedial to choanae, oblique in outline, about the same size of choana, separated medially by distance less than width of odontophore, each bearing 7–10 teeth; skin on dorsum finely shagreen with interspersed low tubercles; dorsal folds absence, except for thin middorsal fold starting at tip of snout and ending at cloaca; skin of lower flanks and venter areolate; no discoidal or thoracic folds; cloacal sheath absent; two low ulnar tubercles barely evident; outer palmar tubercle oval, large ( Fig. 2 View FIGURE 2 ); subarticular tubercles prominent, round; supernumerary palmar tubercles low and rounded, much lower than subarticular tubercles; fingers bearing narrow lateral fringes; Finger I shorter than Finger II; disc of Finger I not expanded; all other discs slightly expanded, rounded to slightly truncate ( Fig. 2 View FIGURE 2 ); ventral pads defined by circumferential grooves.
Tibia length 40.8% of SVL; foot length 39% of SVL; tarsal and heel tubercles absent; inner metatarsal tubercle oval, about twice the size of the outer, rounded tubercle; subarticular tubercles round; plantar supernumerary tubercles indistinct; toes bearing narrow lateral fringes; webbing absent; all other toe discs expanded, rounded; toes with ventral pads well defined by circumferential grooves; relative length of toes: I <II <III <V <IV; Toe V longer than Toe III.
Measurements of holotype (in mm). SVL 21.3; tibia length 8.7; foot length 8.3; head length 8.3; head width 8.2; interorbital distance 2.5; upper eyelid width 1.7; eye diameter 2.3; tympanum diameter 1.0; radioulna length 4.6; hand length 5.1; Finger I length 3.2; Finger II length 3.6; Disc of Finger III width 0.7.
Coloration of holotype in preservative. Dorsal surfaces pale brown with dark gray marks, including interorbital bar, W-shaped occipital mark, sacral chevron, as well as thin middorsal line. Arms faintly barred. Thighs cream with black, oblique stripes. Flanks cream with contrasting black marking; throat cream with dark brown flecks. Venter cream with black reticulum ( Fig. 1 View FIGURE 1 ).
Coloration in life (based on field notes by Alejandro Arteaga and photographs; Fig. 3 View FIGURE 3 ). Frogs have a dorsal brown to reddish brown coloration, with or without small white dots and dark brown marks (i.e., interorbital bar, chevrons). Black spots on groin area, indistinct when the animal is at rest. Flanks and ventral surfaces cream to gray, with fine blackish mottling that in most individuals forms a reticulated pattern; ventral portion of fingers light reddish brown. Iris cream to pale brown with reddish brown to orange horizontal streak.
Variation. Males present vocal slits and median, subgular vocal sac, and lack nuptial pads. Ulnar and tarsal tubercles are more conspicuous in males than in females. Some males present a thin vertebral fold (QCAZ 27553, 27505) and/or dorsolateral folds (QCAZ 27553, 12602, 27554). Dorsal and ventral color variation in preservative is shown in Figure 4 View FIGURE 4 .
Vocalization. (QCAZ 49764; adult male recorded on Reserva Mazar on February 2010, temperature not recorded). Pristimantis mazar emits calls at an approximate rate of 9.3 calls per minute (n = 1). Each call is composed by a pulsed, non-frequency modulated note ( Fig. 5 View FIGURE 5 ). Call duration is 0.14– 0.022 s (mean = 0.0185 ± 0.0032; n = 12); intervals between calls vary from 4.89 to 12.24 s (mean = 6.36 ± 2.097; n = 11). The fundamental dominant is at 3122–3171 Hz (mean = 3148.9 ± 12.551; n = 12); the 1st harmonic is at 6169–6317 Hz (mean = 6244.4 ± 37.792; n = 12) and the 2nd harmonic is at 9294–9415 Hz (mean = 9328.2 ± 37.358; n = 12).
Etymology. This specific epithet mazar refers to the type locality of the species, Reserva Mazar . The epithet is a noun in apposition.
Distribution. Pristimantis mazar occurs at elevations between 2895 and 3415 m in three localities within Reserva Mazar (Provincia Cañar, Ecuador; Fig. 6 View FIGURE 6 ): La Libertad (S02.54804, W78.69741), Rumiloma (S02.57120, S78.74563), and Gasualpampa (S02.55558, W78.70150). The area is a patchwork herbaceous paramo, native evergreen montane forest, shrub, secondary forest and pastures.
Natural history. At Reserva Mazar , two distinct seasons are recognizable: a dry season (Oct/Nov - Jan/Feb) and a wet season (FCT 2008). Pristimantis mazar is a nocturnal, cryophilic and hygrophilic specialist of herbaceous/shrubby paramos and adjacent high evergreen montane forests. This dominant, and over-abundant rainfrog is active whenever the ambient temperature and humidity levels are favorable (mostly at night), at ground level and on low vegetation (0–67 cm from ground). Diurnal retreats for Pristimantis mazar include grass tussocks, leaf-litter and bromeliads. The species is highly vocal and ubiquitous in open paramo, but considerably less vocal and dominant within unbroken forest. Sympatric species of Pristimantis mazar include P. bambu , P. pycnodermis , P. gagliardoi , and P. r i v e t i.
Discussion. The complexity of the genus Pristimantis is exacerbated by its outstanding diversity and recurrent morphological similarities. At this moment, the most recent topologies of Pristimantis (Heges et al. 2008; Pinto- Sanchéz et al. 2012), which contains about 20% of the described diversity of the genus, shows that the P. o re s t e s group is not monophyletic. Under this hypothesis, similar morphologies (those that diagnose the group) have evolved multiple times, probably because of comparable environments and selective forces acting on species of different clades. Furthermore, at least one additional species groups within Pristimantis (i.e., P. myersi group) shares multiple morphological features that characterize the P. o re s t e s group, also suggesting convergence. Within the P. o re s t e s group, the most closely related species to P. m a z a r is P. simonbolivari ( Fig. 7 View FIGURE 7 ), a species restricted to Caschca-Totoras, on the Ecuadorian Andes. However, our interpretation on the species relationships and evolution of morphological traits in Pristimantis may change as taxon sampling increase and more species are discovered. Currently, we hope that contributions like ours help in having a better understanding of this frog genus, as well as identifying areas of high endemism for conservation proposes. Given the available information on Pristimantis mazar , and following IUCN (2001) criteria, we place this species in the Data Deficient category. As the English common name for this species, we suggest Mazar Rainfrog. We suggest the name Cutín de Mazar as the common name in Spanish.




No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.