
Eufriesea zhangi Nemésio & Santos Júnior, 2013
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3609.6.2 |
|
publication LSID |
lsid:zoobank.org:pub:42A080B9-FFFB-44FD-9221-5B98CE840296 |
|
DOI |
https://doi.org/10.5281/zenodo.5621822 |
|
persistent identifier |
https://treatment.plazi.org/id/473887CA-FFC7-C477-FF79-FCC3FB24CEDD |
|
treatment provided by |
Plazi |
|
scientific name |
Eufriesea zhangi Nemésio & Santos Júnior |
| status |
sp. nov. |
Eufriesea zhangi Nemésio & Santos Júnior View in CoL , sp. n.
Diagnosis (males only). Eufriesea zhangi sp. n. is easily assigned to the genus Eufriesea due to the following characters (Faria Jr. 2009): bilobed gonostylus, tentorial pit situated on a conspicuous depression at the edge of clypeus, and lower limit of clypeus arch-shaped, with edges projected over the labrum. It is also easily assigned to the E. mussitans species group due to (Kimsey 1982): head and mesosoma darkly colored, three clypeal ridges, short tongue, not exceeding hindcoxa. Eufriesea zhangi sp. n. can be readily distinguished from the very similar E. nordestina by the presence of a prominent, elevated medial ridge on the clypeus (only an impuctated stripe in E. nordestina ), mesobasitarsus black without metallic hues clothed in dense golden hairs medially and black bristles laterally (in E. nordestina mesobasitarsus is black with purple hues clothed in sparser golden and black hairs mixed), mesotibial brush vestigial, connected to mesotibial tuft (in E. nordestina mesotibial brush is well developed and not connected to mesobitial tuft), metatibia with very dense punctation (punctures separated by 0.05 mm to 0.07 mm) and entirely clothed in golden setae (in E. nordestina punctation on metatibia is very sparse – punctures separated by 0.07 mm to 0.18 mm—and it is clothed in sparse black setae), S7 with two long lobes (in fact, the longest S7 lobes of all known species of Eufriesea ) basally glabrous, as well as the region between lobes, and clothed in dense and long black setae distally( Figure 6 View FIGURE 6 A) (in E. nordestina lobes are shorter and the region between lobes is clothed in dense and long setae; Figure 6 View FIGURE 6 B), gonostylar ventral lobe slightly curved and pointed ( Figure 6 View FIGURE 6 E, G) (in E. nordestina it is straight and rounded; Figure 6 View FIGURE 6 F, H). Eufriesea zhangi sp. n. is consistently larger than E. nordestina (see Appendix 1). Eufriesea zhangi sp. n. can also be easily distinguished from all other predominantly black species of Eufriesea — E. auriceps , E. buchwaldi , E. fuscatra , E. nordestina , and E. venezolana due to its metatibia entirely clothed in dense golden setae ( Figure 4 View FIGURE 4 E–F) (sparser blackish setae in the other species). Although molecular data placed Eufriesea zanghi sp. n. as closer to E. nigrohirta than to any of the blackish species of Eufriesea , both species can be easily distinguished due to mesosomal (entirely blackish in Eufriesea zanghi sp. n. and with a dense stripe of yellow setae on basal portion of scutum in E. nigrohirta ) and metasomal coloration (entirely blackish in Eufriesea zanghi sp. n. and greenish with reddish hues on T2-T 7 in E. nigrohirta ). A black morph also occurs in E. nigrohirta , but even these specimens present strong reddish hues on metasoma (particularly on T2) and also have the yellowish stripe of setae on mesosoma (see Nemésio 2005: 43).
Type material. HOLOTYPE: male, with the following label data: “ BRASIL, CE, Ubajara, Parque Nacional Ubajara, 03º49’53.4”, 40º54’ 02.4W, 456m 26.iv.2012 Vanilina, J.E. Santos Jr, UFMG IHY 1207628.
Description (Male, Figures 4 View FIGURE 4 , 6 View FIGURE 6 ): Color and vestiture. Face metallic green; lower clypeus and malar area metallic purple ( Figure 4 View FIGURE 4 B); gena and upper frons blackish without metallic hues; mesoscutum metallic dark green basally and black without metallic hues distally entirely clothed in dense black setae ( Figure 4 View FIGURE 4 A), tegulae bright green anteriorly and black with strong dark blue and purple hues distally; scutellum black without metallic hues clothed in dense black setae ( Figure 4 View FIGURE 4 A); metasomal terga and sterna black without metallic hues clothed in sparse, short and appressed black setae ( Figure 4 View FIGURE 4 C).
Head. Width 7.13 mm; interorbital distance at base of scape 3.35 mm; maximum interorbital distance 3.72 mm; scape 2.17 mm; eye length 4.46 mm. Clypeus with strong medial ridge ( Figure 4 View FIGURE 4 B), densely punctate; punctures irregular in shape and size (0.03 mm to 0.1 mm).
Body. Body length ca. 18.35 mm; anterior wing ca. 18.23 mm; tongue length 8.06 mm, reaching S1; scutellum 5.33-mm wide and 2.6-mm long; abdominal width 7.9 mm.
Legs. Foretibia and forebasitarsus black with purple hues fringed with long and dense, black hairs; mesobasitarsus black without metallic hues, clothed in dense golden setae medially and long black bristles laterally; velvet area occupying the entire outer surface of mesotibia, mesotibial tuft long and triangular, occupying about 1/6 of velvet area, marginally connecte to an almost vestigial brush ( Figure 4 View FIGURE 4 D); metatibia oblong, swollen, black without metallic hues clothed in dense golden hairs, dense punctation comprised of circular and small (0.05 mm in diameter) punctures separated from each other by 0.05 mm to 0.07 mm; post-glandular area reaching apex ( Figure 4 View FIGURE 4 E–F).
Terminalia . S7 arch-shaped and strongly bilobed, both lobes basally glabrous, as well as region between lobes, and medially to apically covered with dense black setae ( Figure 6 View FIGURE 6 A); S8 pointed apically ( Figure 6 View FIGURE 6 C); gonostylar dorsal lobe longer than ventral one and clothed in dense setae; gonostylus three to four times longer than gonocoxal dorsal lobe ( Figure 6 View FIGURE 6 G).
Female. Unknown.
Etymology. The specific epithet honors Dr. Zhi-Qiang Zhang, editor-in-chief of Zootaxa. One decade ago, Krell (2002), in an influential paper in Nature, stated that one of the reasons for the low impact factor of taxonomy was the lack of “core journals for general taxonomy. These exist for cladistics, biogeography, chemical systematics, and so on, but not for species descriptions, revisions of genera, identification keys or inventories.” This situation has dramatically changed in the course of only ten years and Zootaxa became the most influential core journal on zoological alpha-taxonomy ever, publishing around 20% of all new nomina in zoology yearly. This is thanks to Dr. Zhang’s visionary perspective and we here dedicate this beautiful new orchid bee to him in recognition of his accomplishments in this fantastic enterprise.
Type locality. Parque Nacional de Ubajara, municipality of Ubajara, state of Ceará, northeastern Brazil, at 03º49’02.4”S / 40º54’02.4”W, 456 m above sea level.
Attractive baits: Vanillin.
Discussion
Although outstandingly conspicuous orchid-bee species have recently been described from time to time (e.g. Moure 1996, Nemésio 2006, Faria & Melo 2011, Oliveira 2011), the recent boom in orchid-bee alpha-taxonomy is mostly due to the recognition of cryptic or sibling species among long established taxa (e.g. Roubik 2004; Rasmussen & Skov 2006; Hinojosa-Díaz & Engel 2007; Bembé 2007, 2008; Ayala & Engel 2008; Nemésio & Bembé 2008; Nemésio 2008, 2010, 2011b, c, d, 2012; Eltz et al. 2011; Hinojosa-Díaz et al. 2011, 2012; Nemésio & Engel 2012). We here continue this recent tradition by describing a species similar to E. auriceps and E. nordestina , but as far as we know, geographically restricted to an Atlantic Forest remnant at the top of a mountain in the state of Ceará, northeastern Brazil. Although the most conspicuous character distinguishing this newly described species from other black Eufriesea is the presence of dense golden setae on metatibiae, it was here shown that this character is not just an extreme color variation within the sympatric E. nordestina . In fact, E. nordestina presents a wide range of color variation, especially concerning the metasoma, which can be entirely clothed in black setae (most specimens) or present yellowish setae as well. One extreme of these yellowish color variants was even described by Moure et al. (2001) as E. aridicola —specimen illustrated by Nemésio (2009: 37). This phenomenon has also been seen among E. auriceps specimens (Nemésio 2009: 36) and usually affects most strongly T2 and T3, although yellowish setae may be found on T4-T7 as well in some specimens (A. Nemésio, pers. obs.). The distinctness of Eufriesea zhangi sp. n., on the other hand, is supported not only by the diagnostic golden setae of the metatibia (a trait not subject to such a wide variation in both E. auriceps and E. nordestina ), but it is corroborated by its larger size ( Figure 2 View FIGURE 2 , Appendix 1), a set of distinctive morphological features, including morphology of genitalia (see ‘Diagnosis’ section), and also by molecular data. Since it was anticipated by Faria & Melo (2011) that a full revision of the black species of the E. mussitans species group may be in course, recognition of Eufriesea zhangi sp. n. as a distinct (and probably rare) species from a remote site in northeastern Brazil may be of importance for a thorough understanding of the richness and limits of speciation within this particular group of bees.
Although Eufriesea zhangi sp. n. seems to be morphologically closer to E. auriceps and E. nordestina , the phylogenetic analyses carried out with COI suggested that E. nigrohirta might be closely related to the new species. Interestingly, E. nigrohirta is also restricted to mountain tops, but in the states of Minas Gerais and Bahia (Nemésio 2005; Azevedo et al. 2010). Nevertheless, E. nigrohirta displays several contrasting morphological characteristics that distinguish it from Eufriesea zhangi sp. n. (see Nemésio 2005). It should be emphasized, however, that the association between both species in our molecular analyses presented a relatively low support and future studies must be carried out to further investigate their relationships. In fact, detailed phylogenetic studies are still needed to elucidate the relationships among species and groups of Eufriesea to overcome limitations revealed in previous studies (e.g. Faria Jr. 2009; Ramírez et al. 2010).
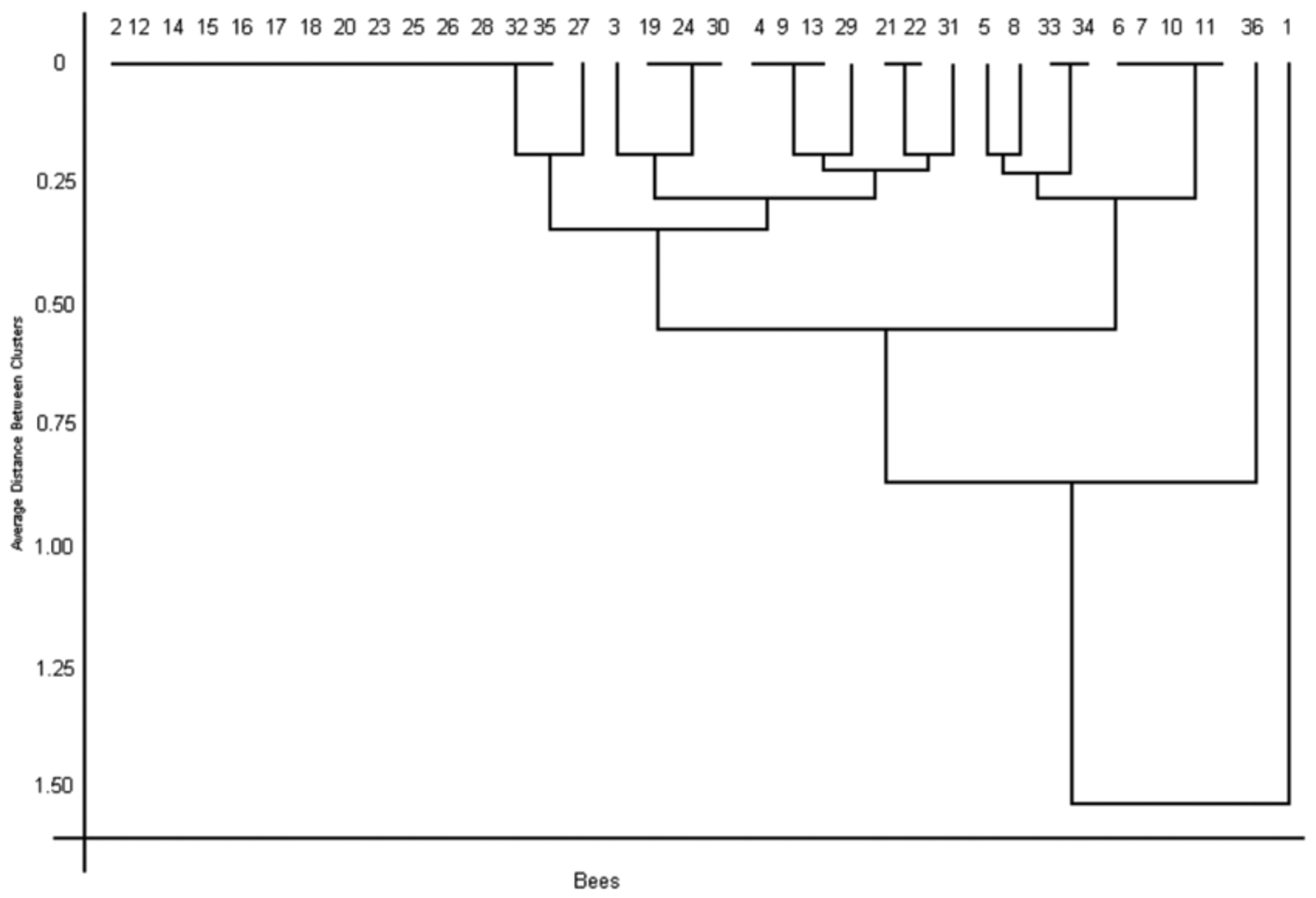
Sheffield et al. (2009), studying the bee fauna of Nova Scotia, established that individuals of a given species share little intra-specific variation with COI divergences averaging just 0.49%. Our results have shown an average distance exceeding 1.4% between Eufriesea zhangi sp. n. and the other taxa analyzed ( Table 1 View TABLE 1 ). The relationship among the studied specimens of E. auriceps is also noticeable. The average distance among them was 1.2%, whereas the average distance between both groups of E. nordestina was 0.4%. These results support Moure’s (1999) and Faria & Melo’s (2011) hypothesis that E. auriceps is a species complex (contra Nemésio 2009).
Finally, the position of Eufriesea zanghi sp. n. as potentially close to E. nigrohirta suggests that this latter species may be part of the E. mussitans species group. If this is true, the re-arrangement of the Eufriesea auripes (Gribodo, 1882) species-group proposed by Nemésio & Bembé (2008) is not correct. In fact, E. auripes speciesgroup was supported as monophyletic neither by Faria (2009) nor by Ramírez et al. (2010).


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |