
Orisarma intermedium (De Haan, 1835)
|
publication ID |
https://doi.org/10.26107/RBZ-2020-0097 |
|
publication LSID |
lsid:zoobank.org:pub:815E4670-B063-4FD8-B31E-3AD89B3A7942 |
|
DOI |
https://doi.org/10.5281/zenodo.6532424 |
|
persistent identifier |
https://treatment.plazi.org/id/49152B56-FFF4-BA3A-FC57-FECEFF73FAA4 |
|
treatment provided by |
Diego |
|
scientific name |
Orisarma intermedium (De Haan, 1835) |
| status |
|
Orisarma intermedium (De Haan, 1835) View in CoL
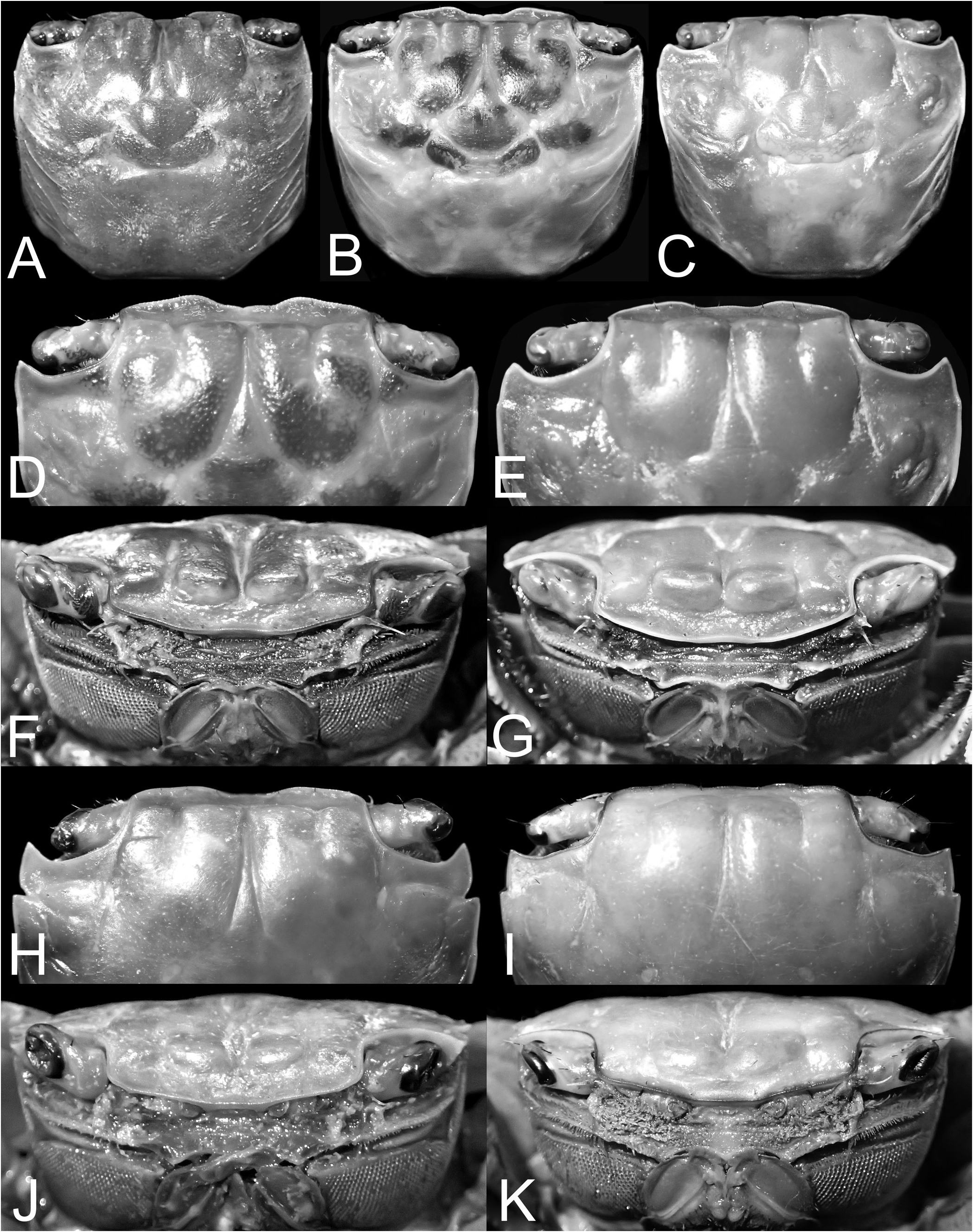
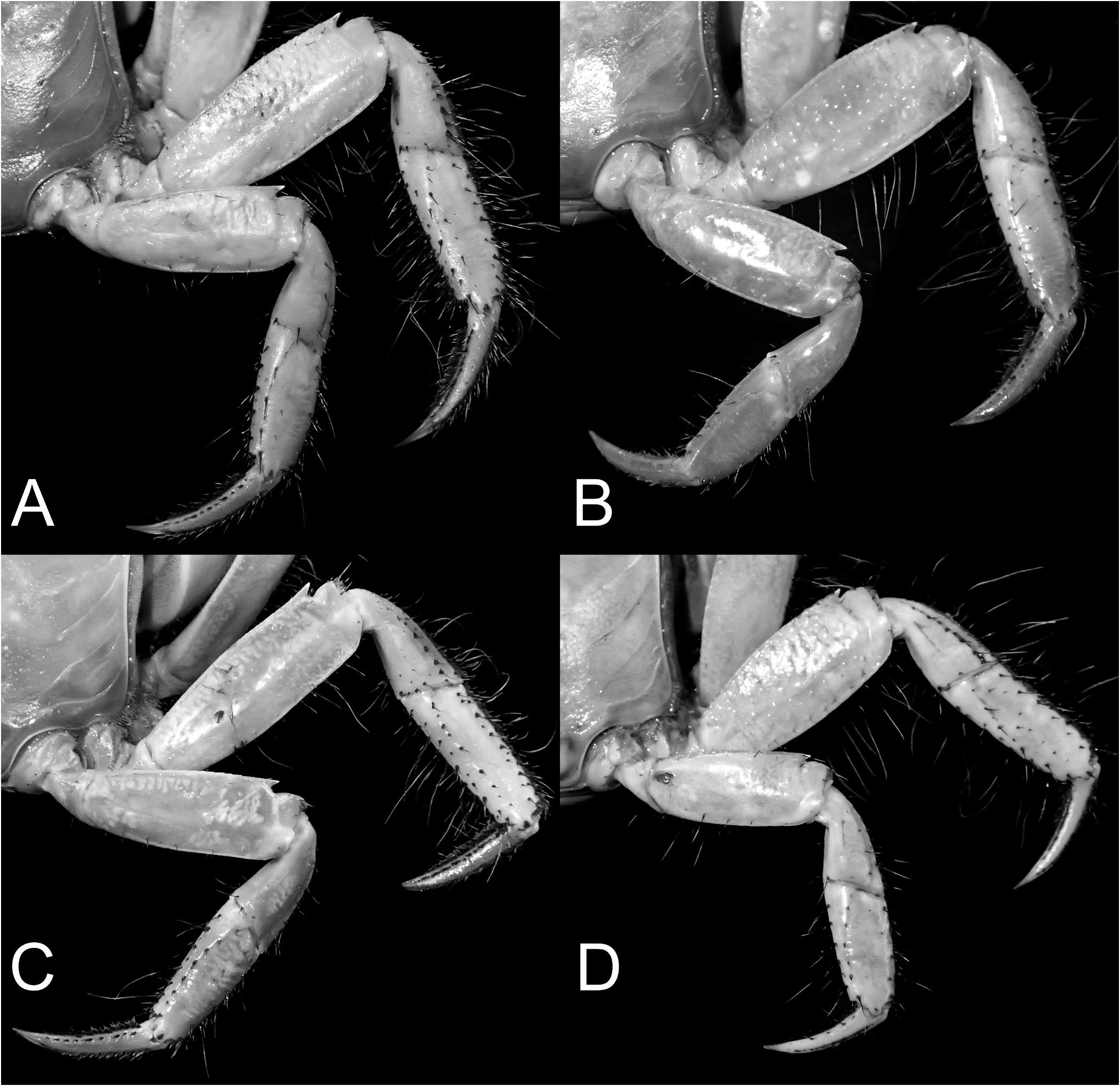
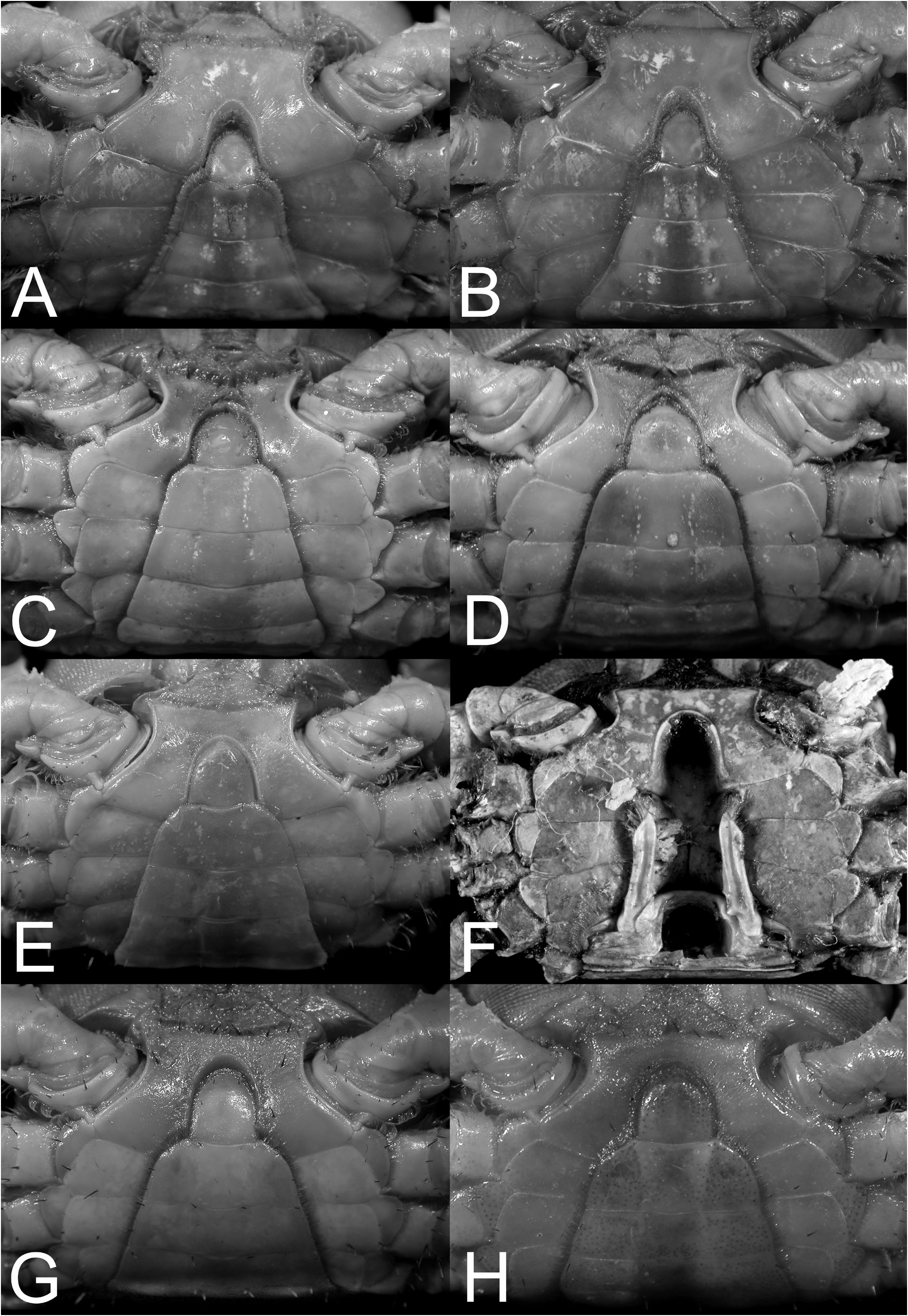
( Figs. 2A–F View Fig , 3D View Fig , 4H, J View Fig , 6A–D View Fig , 8A–C View Fig , 9E View Fig , 12A–E View Fig , 33C, F View Fig , 43C View Fig , 54A–C View Fig )
Grapsus ( Pachysoma) intermedius De Haan, 1835: 61 View in CoL , pl. 16 fig. 5.
Sesarma intermedia View in CoL – H. Milne Edwards, 1853: 186; Stimpson, 1858: 105; Herklots, 1861: 17; De Man, 1880: 25; Miers, 1880: 314; De Man, 1887: 649; De Man, 1892: 337; Ortmann, 1894: 721; Stimpson, 1907: 133.
Sesarma intermedius – Fransen et al., 1997: 130.
Sesarma ( Sesarma) intermedia View in CoL – Tesch, 1917: 162; Balss, 1922: 155 (part); Sakai, 1936: 235, pl. 65 fig. 3; Sakai, 1939: 684, pl. 78 fig. 3; Shen, 1940a: 95; Shen, 1940b: 236; Kamita, 1941: 224, fig. 124; Sakai, 1965: 202, pl. 97 fig. 3; Kim, 1973: 489, text-fig. 218, pl. 94 fig. 168.
Sesarma ( Sesarma) intermedium – Serène, 1968: 105.
Sesarmops intermedium – Serène & Soh, 1970: 401; Sakai, 1976: 659, pl. 224 fig. 3; Matsuzawa, 1977: pl. 111 fig. 1; Soh, 1978: 11, pl. 4b; Takeda, 1982: 221, fig. 655; Miyake, 1983: 179, pl. 60-3; Nagai & Nomura, 1988: 44; Muraoka, 1998: 54; Minemizu, 2000: 298; Kobayashi, 2000: 122, fig. 2o; Tsai et al., 2000: 61; Ng et al., 2001: 43 [includes references for this species from Taiwan up to 2001]; Ho, 2003: 32, 33; Cuesta et al., 2006: 162, fig. 13A–E; Takeda & Ueshima, 2006: 99; Yang et al., 2008: 802; Lee, 2008: 133; Liu, 2009: 49, fig. 19; Naderloo & Schubart, 2009: 62, 63, 67; Liu & Wang, 2010: 62, 63; Rademacher & Mengedoht, 2011: 31; Huang & Liu, 2012: 267; Li & Chiu, 2013: 57; Toyota & Seki, 2014: 192, 193; Ng et al., 2017b: 106 [for other references for Taiwan].
“ Sesarmops ” intermedius – Ng et al., 2008a: 220.
Sesarmops intermedius View in CoL – Hong et al., 2010: 259, fig. 3b; Lee et al., 2012: 183; Lee, 2015: 321.
Material examined. Lectotype male (23.0 × 19.9 mm) ( RMNH D 165 About RMNH ), Japan, coll. H. Bürger. Paralectotype: 1 female ( RMNH D 165 About RMNH ), same data as lectotype. Others: JAPAN – 1 male (27.1 × 25.2 mm) ( ZRC 1970.2.23.6), coll. T. Sakai, 1968 ; 1 female (22.7 × 20.0 mm) ( ZRC 1970.8.27.3), Manazuru, Sagami Bay , coll. T. Sakai, 1968 ; 1 female (23.0 × 20.4 mm) ( ZRC), river mouth, Izaku River, Satsuma Peninsula , Kagoshima Prefecture, coll. H. Suzuki, 30 August 2000 ; 4 males (largest 27.0 × 25.0 mm), 4 females (largest 32.1 × 29.3 mm) ( ZRC 2013.0140 View Materials ), in brackish water stream, Tomioka, Amakusa, Shikizuki , Japan, coll. J. Lai, 2 September 2002 ; 1 male (36.4 × 32.4 mm), 2 females (largest 29.0 × 26.9 mm, smallest 33.5 × 30.4 mm) ( ZRC 2000.2262 View Materials ), Iriomote Island, Ryukyu Islands , coll. Y. Cai & N.K. Ng, 15 June 2000 ; 1 female ( ZRC 2000.2265 View Materials ), Japan, Nagama river second site from headwaters, Okinawa, coll. Y.X. Cai & N.K. Ng, 15 June 2000 ; 1 female ( ZRC 2000.2266 View Materials ), Japan, along the track to the headwaters of Nagama River , found in dead fallen tree trunk, Okinawa, coll. Y.X. Cai & N.K. Ng, 15 June 2000 ; 1 male (29.3 × 26.8 mm) ( ZRC), Yenbaru , Okinawa, coll. B.Y. Lee, 18 September 2012 ; 1 male (37.5 × 35.8 mm) ( ZRC 2014.0265 View Materials ), Zenda Forest Park, Kume Island, Ryukyu Islands , coll. P.K.L. Ng et al., 17 November 2006 . TAIWAN – 1 female ( ZRC 2001.0033 View Materials ), northeastern Taiwan, Ilan county, Tahsi , coll. K.X. Lee, 2000; GoogleMaps 1 ovigerous female, 1 juvenile male, 1 juvenile female ( ZRC), Hualien county, Hualien city, mouth of Meilun stream, 23°58′54″N 121°36′37″E, eastern Taiwan , coll. P.K.L. Ng & H.-C. Liu, 22 June 2002 GoogleMaps ; 1 male, 1 female, 1 juvenile ( ZRC 2000.1834 View Materials ), Rangkon river mouth, coll. H.-C. Liu & C.D. Schubart, 14 September 1999 ; 4 males (largest 23.5 × 21.2 mm, smallest 29.0 × 25.8 mm), 6 females (largest 23.9 × 20.7 mm, smallest 28.0 × 24.2 mm) ( ZRC 1999.527 View Materials ), Tung Kou stream, Kenting National Park , Pingtung, Hengchun Peninsula, coll. P.K.L. Ng et al., 2000 ; 1 male, 5 females ( ZRC 2002.0477 View Materials ), Kang Kou Hsi, Kentin Park, Pingtung, southern Taiwan , coll. P.K.L. Ng & C.-H. Wang, 5 August 2002 ; 2 males (29.5 × 26.5 mm, 34.2 × 30.7 mm) ( ZRC 2001.0034 View Materials ), Pingtung, Hengchun, Chang-Wai , coll. P.K.L. Ng, 7 November 2000 ; 1 male, 1 female ( ZRC 2007.0208 View Materials ), purchased from aquarium trade, supposedly from Taiwan , coll. S.H. Tan, 7 May 2007 ; 2 females (17.8 × 15.8 mm, 15.5 × 13.4 mm), rice field mouth, Checheng-Pingtung, southern Taiwan , coll. H.-C. Liu & C.D. Schubart, 15 September 1999 . HONG KONG – 1 male, 2 females ( ZRC 2019.0540 View Materials ), back mangroves, ca. 22°29′44.6″N 114°02′49.7″E, Mai Po Nature Reserve , coll. S. Cannici et al., 24 May 2019 GoogleMaps .
Diagnosis. Dorsal surface of carapace relatively less convex, surfaces usually slightly more rugose, with scattered stiff setae on anterior and branchial regions; frontal lobes separated by deep median concavity; epibranchial tooth distinct even in small specimens, always separated by deep notch; posterolateral margins subparallel; granules on outer surface of palm (especially those below longitudinal ridge) rounded, prominent; merus of ambulatory legs relatively longer; chitinous distal part of G1 short, tip truncate. In life, carapace and chelae red, sometimes yellow.
Colour. In life, the dorsal surfaces of the carapace, chelipeds, and ambulatory legs are bright red with the fingers white ( Fig. 54A View Fig ) (see also Miyake, 1983; Lee, 2008; Liu, 2009; Liu & Wang, 2010; Rademacher & Mengedoht, 2011), although some orangish-red specimens are sometimes observed in southern Taiwan. There is an interesting large male collected from the Ryukyus which is bright yellow ( Fig. 54B View Fig ), but this specimen was obtained over a kilometre from the sea on a muddy patch up on a hill, and its colour may be a result of fading. Its morphology is identical to more typically coloured specimens. Smaller specimens and some females may have a dark tranverse band across their frontal margin ( Fig. 54C View Fig ).
Remarks. Orisarma intermedium is very close to O. sinense in general carapace form and smaller specimens are not easy to separate. Many of the old records of these two species in the literature (especially those on mainland China) will need to be rechecked. In an unpublished key, which was the basis of the more recent identifications of Serène (1968), Serène & Soh (1970), and Soh (1978), the primary characters were whether the epibranchial tooth is clearly demarcated (present in O. intermedium versus absent in O. sinense ), whether the outer surface of the chela was strongly granulated, and the presence of a low longitudinal median ridge (strongly granulated with a ridge in O. intermedium versus low granules and no ridge in O. sinense ), the relative proportions of the fingers of the chela (dactylar finger longer than height of palm in O. intermedium versus dactylar finger shorter in O. sinense ) and ambulatory merus (merus of third ambulatory leg 2.5 times longer than broad in O. intermedium versus 3 times in O. sinense ), and proportions of the chitinous distal part of G1 (shorter in O. intermedium versus longer in O. sinense ).
Examination of the present material reveals that smaller specimens of O. sinense do have the epibranchial tooth relatively low and while it may appear to be absent and/or almost confluent with the rest of the margin, it is nevertheless always visible (as in the case of small specimen from Hong Kong, ZRC 1975.6.30.10). In medium-sized specimens, the epibranchial tooth is distinct but demarcated from the rest of the margin by a weak notch ( Fig. 2I View Fig ). Only in the largest specimen is the epibranchial tooth really prominent, and with a deeper notch separating it from the rest of the margin ( Figs. 2J View Fig , 4I View Fig ). In O. intermedium , the epibranchial tooth is distinct even in small specimens and always separated by a deep notch (e.g., Figs. 2A–F View Fig , 4H View Fig ). Soh (1978) suggested that the cheliped of O. sinense has relatively shorter fingers but this is not a useful character and was probably due to the small size of the specimens he examined. Larger specimens of O. sinense ( Fig. 6E–H View Fig ) have the same kind of fingers as those of O. intermedium ( Fig. 6A–D View Fig ). The outer surface of the adult male chela of the two species is slightly different, but the low longitudinal median ridge is present on both species ( Fig. 6A–G View Fig ). In adult O. intermedium , the granules on the outer surface of the palm, especially those below the longitudinal ridge, are rounded and prominent ( Fig. 6A–D View Fig ). In the case of O. sinense , the granules are lower and more flattened ( Fig. 6E–G View Fig ). The proportions of the ambulatory legs are not useful, and in O. intermedium (which supposedly has longer legs), this character is too variable to be reliable. There seems to be a tendency (but not always) for the merus of the ambulatory legs to be relatively wider and shorter in smaller specimens (e.g., Fig. 8A, B View Fig ), with those of large specimens becoming longer and more slender ( Fig. 8C View Fig ). In fact, on the basis of the specimens of both species available, the ambulatory meri of O. sinense actually appear proportionately shorter ( Fig. 8D View Fig versus Fig. 8A–C View Fig ) when specimens of similar size are compared. The G1 differences, however, are valid and reliable ( Fig. 12G–J View Fig versus Fig. 12B–E View Fig ).
In addition to the epibranchial tooth, chela, and G1 differences noted above, adults of the two species can be separated by the form of the frontal margin and carapace physiognomy. In O. intermedium , the two frontal lobes are separated by a deep median concavity ( Fig. 4H View Fig ) but in O. sinense , this concavity is distinctly shallower ( Fig. 4I View Fig ). The dorsal surface of the carapace of O. intermedium is relatively less convex, with the surfaces usually slightly more rugose and with scattered stiff setae all over the anterior and branchial regions ( Figs. 2A–F View Fig , 4H, J View Fig ). In O. sinense , the dorsal surface of the carapace is more convex (notably the branchial), smoother, and the setae are more scattered and sparser on the branchial regions ( Figs. 2I, J View Fig , 4I, K View Fig ). These characters are valid for both sexes as well as for small and subadult specimens.
De Man’s (1888: 182) “ Sesarma intermedia ” from Mergui is a misidentification of Manarma moeschii ( De Man, 1892) . The specimen figured by Soh (1978: pl. 4b) is almost certainly O. intermedium on the basis of his photograph, and we have also examined material from Hong Kong. Thus, both O. intermedium and O. sinense occur sympatrically in Hong Kong.
Hoang et al. (2012: 75, 78) reported “ Sesarmops intermedius ” from southern Vietnam but their record is dubious as the species is not known to be present that far south; and it is more likely their material was confused with a species of Pseudosesarma .
Biology. This species is common in freshwater streams and waters near the sea, which may still be subject to some tidal influence, although they can also be found several hundreds of metres away on hilly areas. They normally occur among dense vegetation along these bodies of water, coming out at night to forage on the sand- and mudflats. They are primarily herbivores and scavengers. Hong et al. (2010) discussed its ecology in Korea. The physiology of the species in Taiwan was studied by Tsai et al. (2000) while the visual orientation and habits was done by Huang & Liu (2012). The reproductive behaviour of this species in Japan was discussed by Kyomo (1986). The larvae have been reported on by Fukuda & Baba (1976), Cuesta et al. (2006), and Lee (2015). The microsatellite markers of the species were studied by Lee et al. (2012). Nematode parasites have been reported on by Yoshimura (1990). In Taiwan, O. intermedium is collected in large numbers from southern Taiwan for the aquarium trade (as a freshwater crab), and many populations have been seriously depleted.
Distribution. Despite the lack of genetic differentiation in the current genetic dataset, we are confident O. intermedium and O. sinense are separate species on the basis of their morphology. However, we are less sure about their distribution. In Japan, Korea, and Taiwan, only one species, O. intermedium , is known for certain. On mainland China, notably southern China, both species have been reported ( Shen, 1940a, b; Dai et al., 1986; Dai & Yang, 1991). Certainly, both species appear to be present in Hong Kong (see Soh, 1978). The older material will need to be reexamined and fresh collections made to determine their actual distributional limits. In the most recent treatments of the Chinese fauna ( Dai et al., 1986; Dai & Yang, 1991), O. intermedium is not treated at all, even though this species is certainly present in Taiwan and Hong Kong, whereas O. sinense is treated as a species of Sesarmops (see Dai & Yang, 1991: 539, text-fig. 277(2), pl. 69(6); with their figures of the carapace and G1 leaving no doubt of the identity of this species).







No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
InfraOrder |
Brachyura |
|
Family |
|
|
Genus |
Orisarma intermedium (De Haan, 1835)
| Schubart, Christoph D. & Ng, Peter K. L. 2020 |
Sesarmops intermedius
| Lee HJ 2015: 321 |
| Lee S-K & Jung J & Kang S & Kim W 2012: 183 |
| Hong J-S & Choi J-W & Tsutsumi H 2010: 259 |
“ Sesarmops ” intermedius
| Ng PKL & Guinot D & Davie PJF 2008: 220 |
Sesarma intermedius
| Fransen CHJM & Holthuis LB & Adema JPHM 1997: 130 |
Sesarmops intermedium
| Ng PKL & Shih H-T & Ho P-H & Wang C-H 2017: 106 |
| Toyota K & Seki S 2014: 192 |
| Li J-J & Chiu Y-W 2013: 57 |
| Huang H-D & Liu H-C 2012: 267 |
| Rademacher M & Mengedoht O 2011: 31 |
| Liu H-C & Wang C-H 2010: 62 |
| Liu H-C 2009: 49 |
| Naderloo R & Schubart CD 2009: 62 |
| Yang S & Chen H & Jiang W 2008: 802 |
| Lee J-H 2008: 133 |
| Cuesta JA & Guerao G & Liu H-C & Schubart CD 2006: 162 |
| Takeda M & Ueshima R 2006: 99 |
| Ho P-H 2003: 32 |
| Ng PKL & Wang C-H & Ho P-H & Shih H-T 2001: 43 |
| Minemizu R 2000: 298 |
| Kobayashi S 2000: 122 |
| Tsai M-L & Li J-J & Dai C-F 2000: 61 |
| Muraoka K 1998: 54 |
| Nagai S & Nomura K 1988: 44 |
| Miyake S 1983: 179 |
| Takeda M 1982: 221 |
| Soh CL 1978: 11 |
| Sakai T 1976: 659 |
| Serene R & Soh CL 1970: 401 |
Sesarma ( Sesarma ) intermedium
| Serene R 1968: 105 |
Sesarma ( Sesarma ) intermedia
| Kim HS 1973: 489 |
| Sakai T 1965: 202 |
| Kamita T 1941: 224 |
| Shen CJ 1940: 95 |
| Shen C 1940: 236 |
| Sakai T 1939: 684 |
| Sakai T 1936: 235 |
| Balss H 1922: 155 |
| Tesch JJ 1917: 162 |
Sesarma intermedia
| Stimpson W 1907: 133 |
| Ortmann AE 1894: 721 |
| Man JG De 1892: 337 |
| Man JG De 1887: 649 |
| Man JG De 1880: 25 |
| Miers EJ 1880: 314 |
| Herklots JA 1861: 17 |
| Stimpson W 1858: 105 |
| Milne Edwards H 1853: 186 |
Grapsus ( Pachysoma ) intermedius
| Haan W De 1835: 61 |