
Neophilopterus tricolor ( Burmeister, 1838 )
|
publication ID |
https://doi.org/ 10.1080/00222930500392766 |
|
persistent identifier |
https://treatment.plazi.org/id/4A42CD6D-FF82-FFA8-E88A-FE8440B416CF |
|
treatment provided by |
Felipe |
|
scientific name |
Neophilopterus tricolor ( Burmeister, 1838 ) |
| status |
|
Neophilopterus tricolor ( Burmeister, 1838) View in CoL
( Figures 1–3 View Figure 1 View Figure 2 View Figure 3 )
Docophorus tricolor Burmeister 1838, p 424 View in CoL .
Philopterus tricolor (Burmeister) : Harrison 1916, p 106. Neophilopterus tricolor (Burmeister) View in CoL : Cummings 1916, p 660.
Type host. Ciconia nigra (Linnaeus) .
Redescription
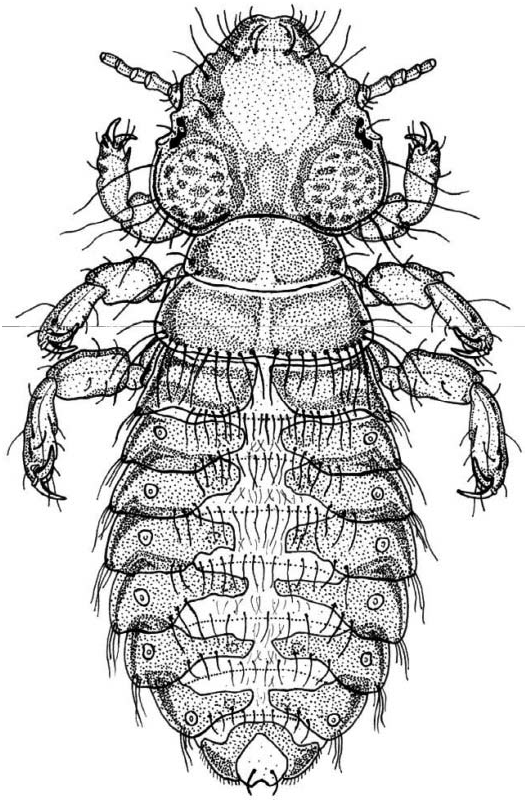
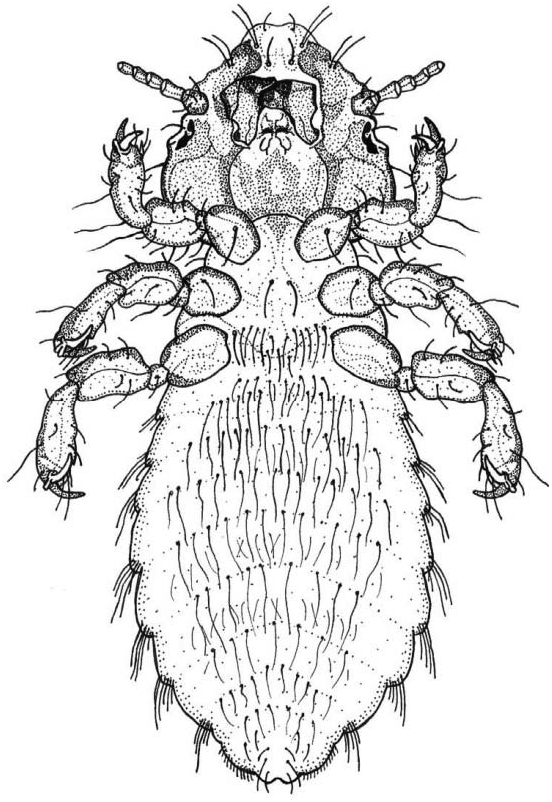
Head large, wider than long ( Table I), with maximum width at the level of the temples; preantennal region shorter than the postantennal region. Frontal zone showing a straight hyaline margin whilst the cephalic anterior edge is lightly concave as an ‘‘osculum’’ (median indentation of the anterior margin of the head: Symmons 1952; Lyal 1985). Scarcely defined clipeal plate; anterior region to dorsal preantennal sutura not medially divided, with two acute angles presents posteriorly, suggesting the fusion of an originally double plate ( Cummings 1916, p 660). Marginal ventral carinae curved, extending from the mandibular framework to support the pulvinus ( Symmons 1952). Large and very sclerotized mandibles. Antennae filiform in both sexes with five antennomeres: scape, pedicel, and three terminal flagellomeres; the scape slightly broader than the remaining ones with two mediate setae; the two terminal flagellomeres showing clearly the sensilar complex ( Soler Cruz and Martín Mateo 1996, 1998; Smith 2001). Temples show a characteristic sculpture of small, more or less circular sclerotizations. Temporal carinas narrow and occiput straight.
Cephalic chaetotaxy: in addition to the five pairs of setae always present throughout the Philopteridae ( Clay 1951) , there is a single submarginal setae lying to each side of the midline (dorsal submarginal setae) on the dorsal surface of the preantenal region. The temporal margins have four long and two short setae, some arising dorsally and some ventrally (all called marginal temporal setae); one small postemporal setae near the midline on each side above the occipital margin. On the ventral surface there are four marginal anterior setae on the marginal carina and two ventral submarginal setae just lateral to the ventral anterior plate; lateral and posterior to the ventral submarginal setae there are two ventral setae on each side (anterior ventral setae).
Thorax wider than long ( Table I). Prothorax with posterolateral corners rounded; 2+2 minute setae near the anterior edge; 2+2 longer setae on the posterolateral corners. Pterothorax laterally divergent; posterior margin indented in the midline with 12+12 long setae; ventrally there are 2+2 minute anterior setae and 5+5 medium setae on the central zone.
Abdomen oval in both sexes; the visible segment I is interpreted as the II, although it is probably I and II fused following interpretation by Wilson (1936) and Smith (2001); also segment IX is the result of IX, X, and XI fused together ( Smith 2001). Segments II–VIII bear the spiracles. Tergopleurites well chitinized; tergites II–VII subtriangular, long and blunt, without reaching the midline, separate by a median gap; pleurites III–VII more sclerotized, each one reaching beyond the anterior segment.
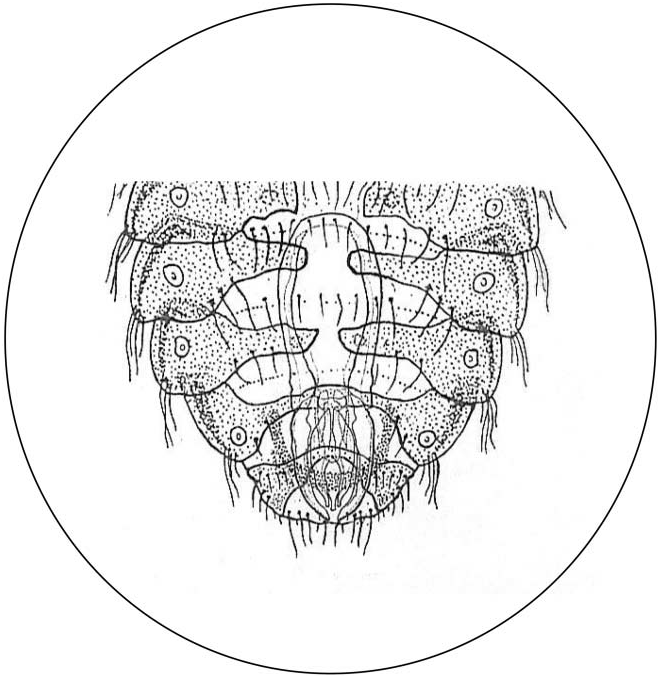
Abdominal chaetotaxy: two rows of tergal setae on each segment, one distal with 10–15 setae arising regularly across the posterior margin of each segment, and another with 8–10 setae on the central surface of each segment. The number of setae in both rows is lower in the terminal segments. There is sexual dimorphism in the terminal segments and in their chaetotaxy. In the male tergites VIII–IX are narrow and continuous across the segment covering almost the entire tergum; tergite VIII with 2+2 lateral setae and 2 central setae; tergite IX with a continuous row of long setae on in the anterior margin; abdominal end rounded with 4+4 very long marginal setae and 1+1 anterior setae; the genital plate is a narrow central plate curved and enlarged ( Figure 4 View Figure 4 ). In the female only tergite IX is continuous but narrow in the middle with lateral groups or clusters of medium long setae (7–8); posterior margin with a fairly deep median notch and many marginal setae (17 on each side); 2+2 central and variably long setae.
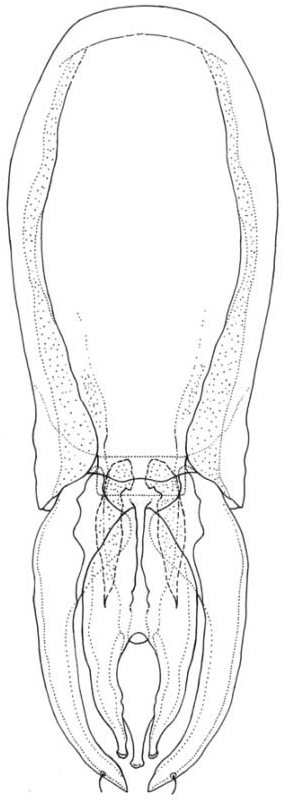
The male genitalia ( Figure 3 View Figure 3 ; Table II) comprise a flattened basal plate or apodema longer than the parameres, with the posterior margin convex and undulated; each lateral margin has a broad band; between these lateral bands the median area of the apodema is trough-shaped; just behind the mesosoma there is a small median plate, with a branch on each side, reaching the lateral regions of the apodema. Parameres are broad, evenly rounded rods, tapering towards the distal end curving slightly inwards; in the base of the mesosoma there is a small process or plate as an upper endomeral chitinization, the ‘‘lower endomere’’ in Cummings (1916); at the end of endomeres is the penis, apparently telomeral, more or less membranous, held by the basal endomeral portion shaped like a pair of pincers.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Neophilopterus tricolor ( Burmeister, 1838 )
| Lanzarot, M. P., Martín-Mateo, M. P., Merino, S. & Fernández-García, M. 2005 |
Philopterus tricolor (Burmeister)
| Harrison L 1916: 106 |
| Cummings BF 1916: 660 |
Docophorus tricolor
| Burmeister HCC 1838: 424 |