
Ryukyua circularis ( Pillai, 1954 )
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4526.2.5 |
|
publication LSID |
lsid:zoobank.org:pub:F2AAFE2D-228D-484F-84C5-83276C25F941 |
|
DOI |
https://doi.org/10.5281/zenodo.5984898 |
|
persistent identifier |
https://treatment.plazi.org/id/4B6C2C30-FF8D-E634-EF8B-B152FD7C6D12 |
|
treatment provided by |
Plazi |
|
scientific name |
Ryukyua circularis ( Pillai, 1954 ) |
| status |
|
Ryukyua circularis ( Pillai, 1954) View in CoL
( Figs 1–6 View FIGURE 1 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 )
Livoneca circularis Pillai, 1954: 17 View in CoL ; 1964: 218–220, figs. 4 L, 5, 7 F. — Bruce 1990: 286. Lironeca circularis Bunkley-Williams & Williams, 1986: 213 –215, fig. 1 A–E. Ryukyua circularis View in CoL .— Williams & Bunkley-Williams, 1994: 160, figs 28–32.— Rameshkumar, Ravichandran & Allayie, 2013:
127–132.— Aneesh, Helna & Sudha, 2016: 1270–1277, fig. 1 (m, n).—Praveenraj, Saravanan, Kumar, Ravichandran,
Baruah, Devi, Sankar & Roy, 2017: 593–595, fig. 1.
Type and type locality: Pillai’s Indian type specimens of Livoneca circularis are not extant. A neotype was not designated here as the species is easily recognizable. Should a neotype be designated, it would ideally be from the type locality (Trivandrum, Kerala Coast, India) and the type host ( Amblygaster leiogaster (Valenciennes, 1847)) .
Material examined: Non–type (All localities from the state of Tamil Nadu, India): 1 ovig. ♀ (11 mm) Parangipettai (11.5084°N, 79.7568°E), 14 April 2017, from Amblygaster clupeoides , coll. S. Ravichandran (ZSI/ MBRC D 1 –551); 5 ovig. ♀, (9–13 mm) Mandapam (9.2770°N, 79.1252°E), 18 May 2017, from Amblygaster clupeoides , coll. P. Vigneshwaran ( CAS / MBRM C-222 to C-226 ) GoogleMaps ; 2 non–ovig ♀ (9, 12 mm) ( CAS / MBRM C-227 to C-228 ) ; 2 ♂ (7, 8 mm) ( CAS / MBRM C-229 to C-230 ); 4 mancae (3.2–4.7 mm) ( CAS / MBRM C-231 to C-234 ) Nagappatinam (10.7656°N, 79.8424°E), 26 March 2016, from Amblygaster clupeoides , coll. P. Vigneshwaran. GoogleMaps
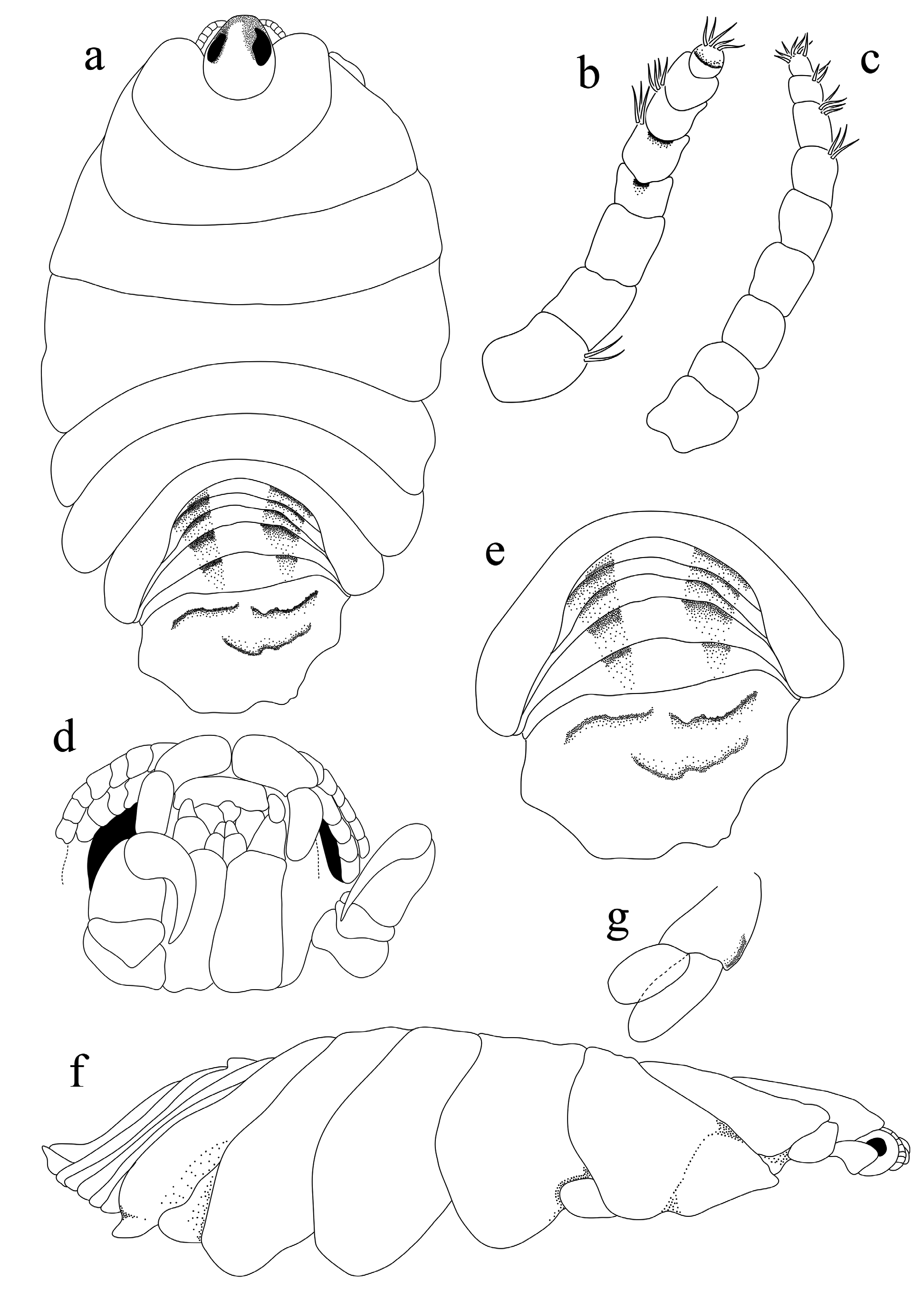
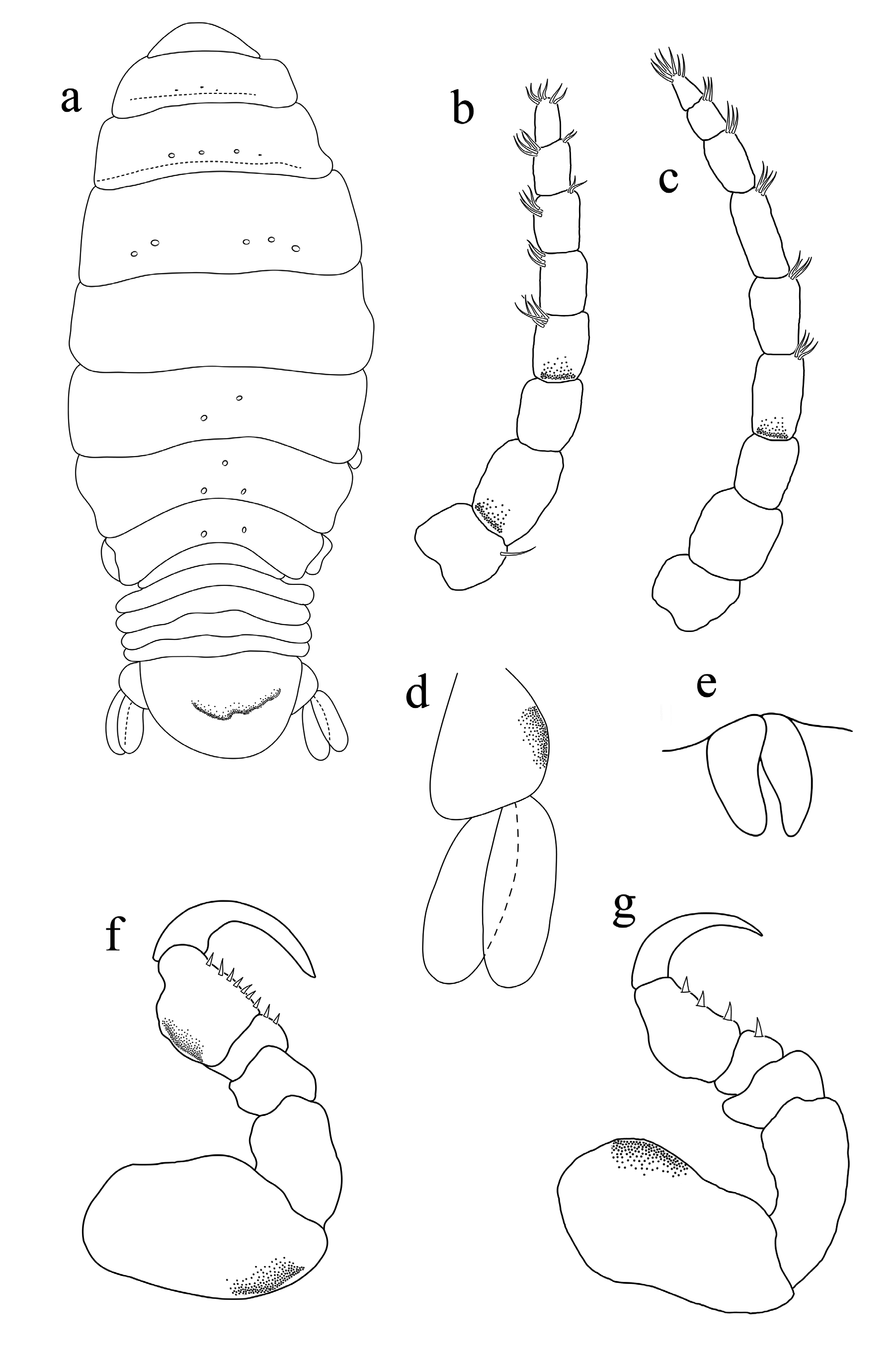
Redescription. Female ( Figs 1–4 View FIGURE 1 View FIGURE 2 View FIGURE 3 View FIGURE 4 ): Body oval to rounded, slightly asymmetrical, 1.2–1.4 times as long as greatest width, lateral margins slightly convex. Cephalon 2.1 times longer than wide, deeply immersed in pereonite 1, anterior margin rounded to form blunt rostrum. Eyes relatively well-developed, 0.2 times width of cephalon.
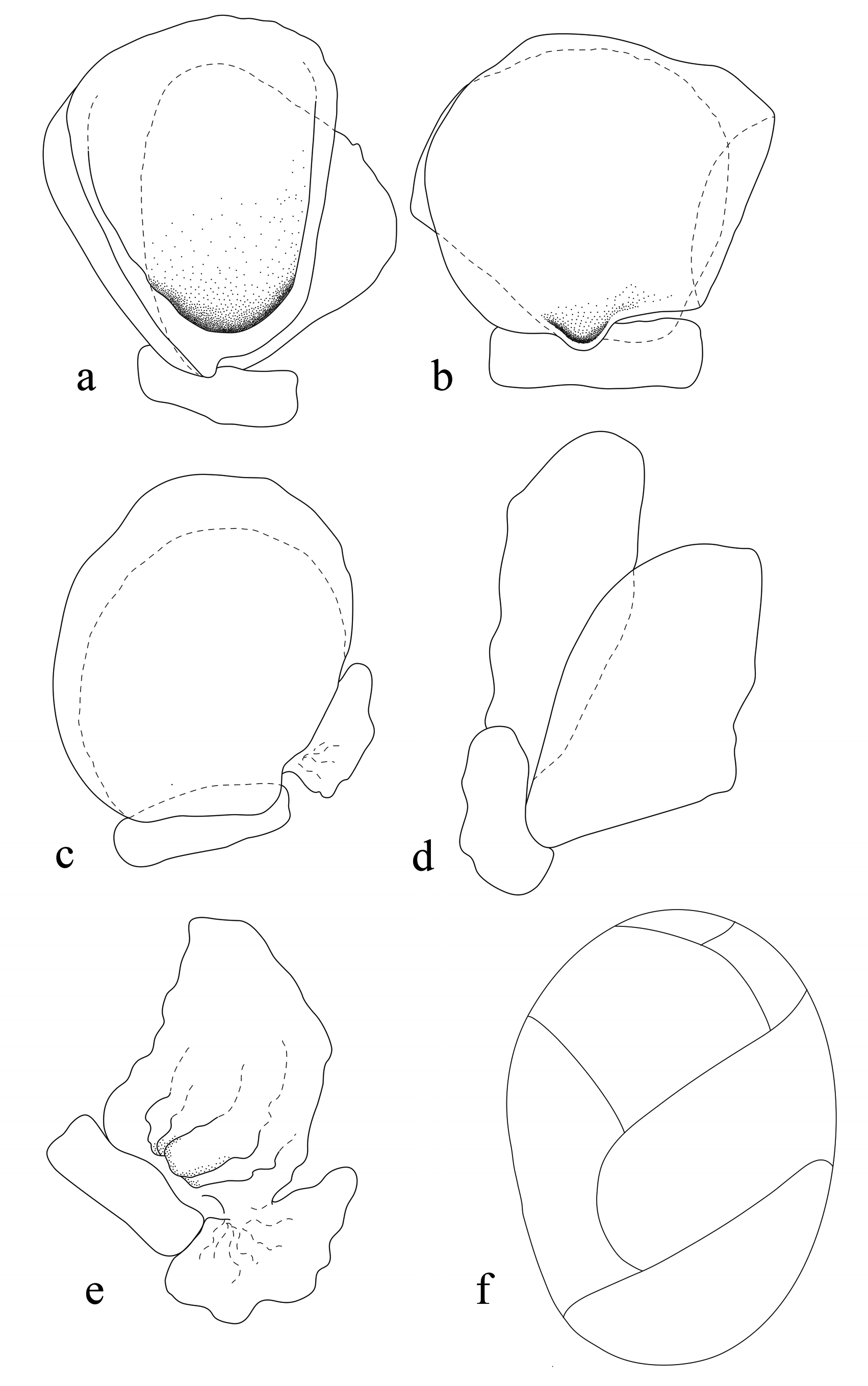
Pereonite lobes increasing in size; pereonite 1 with rounded antero-lateral margins; pereonites 2–5 increasing in width and length; longest at pereonites 4 and 5; widest at pereonite 5; pereonites 6 and 7 decreasing in width and length, lateral margin of pereonite 7 produced posteriorly and overlapping pleonite 1–3. Coxae 2–3 narrow with posteroventral angles rounded; coxae 4–7 acute, partially visible in dorsal view. Pleonites visible in dorsal view; pleonite 1–4 most narrow, posterior margin concave, longer than pleonite 5; pleonite 1–3 partially overlapped by pereonite 7 posterolateral margin, pleonite 5 not overlapped by lateral margins of pleonite 4. Pleotelson 0.6 times as long as anterior width, dorsal surface smooth, anterior lateral margins weakly convex, posterior margin evenly rounded, without median point.
Antennula extending to middle of the eye, composed of 8 articles, slightly stouter than antenna, article 1 and 2 widest and others decreasing in width progressively, article 1 with two setae; article 5, 6 and 8 terminating with 3– 5 short simple setae. Antenna extending to posterior margin of cephalon, composed of 9 articles progressively decreasing in width; clusters of 4–6 short simple setae on articles 3–5, article 9 terminating in 5–7 short simple setae. Mandible with bifid incisor process, molar absent, mandible palp with spiny surface, article 3 with long terminal setae. Maxillula simple, with 3 terminal robust setae. Maxilla apex with spiny surface, medial lobe partly fused to lateral lobe; medial lobe with 2 re-curved robust setae, lateral lobe with 1 large re-curved robust setae. Maxilliped palp article 2 without setae; article 3 with 4 recurved robust setae.
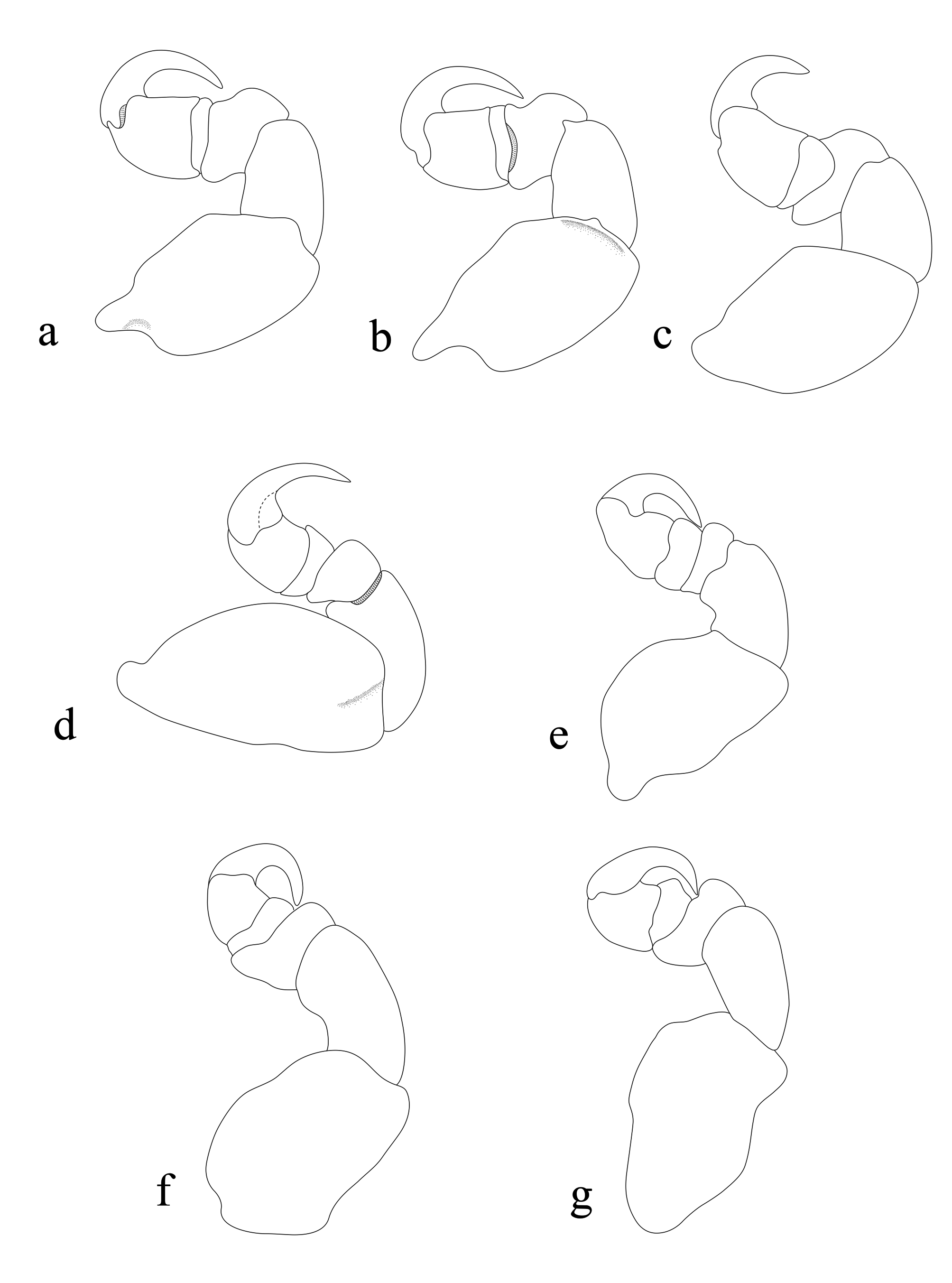
Pereopods with a rather broad basis, elongate ischium and short propodus. Pereopod 1 basis 1.1 times as long as the greatest width; ischium 1.2 times as long as a basis; merus proximal margin with a slight bulbous protrusion; carpus with rounded proximal margin; propodus 1.2 times as long as wide; dactylus slender, 1.1 times as long as propodus, 2.8 times as long as basal width. Pereopod 2 propodus 0.9 times as long as wide; dactylus 1.2 times as long as propodus. Pereopod 5 basis 1.3 times as long as greatest width, ischium 1.5 times as long as basis, propodus 1.1 as long as wide, dactylus 1.3 as long as propodus. Pereopod 6 basis 0.5 times as long as greatest width, ischium 0.9 times as long as a basis, propodus 1.4 times as long as wide, dactylus 2.5 times as long as propodus. Pereopod 7 basis 0.5 times as long as the greatest width; ischium 1.3 times as long as a basis, without protrusions; merus proximal margin with a bulbous protrusion, merus 0.2 times as long as ischium, 0.3 times as long as wide; carpus 0.3 times as long as ischium, without bulbous protrusion, 0.5 times as long as wide; propodus 0.7 times as long as ischium, 1.2 times as long as wide; dactylus slender, 1.6 times as long as propodus, 3.3 times as long as basal width. Oostegites attach to pereopods 2–5.
Pleopods all lamellar, without folding and accessory lobes, endopods of pleopods 3–5 with proximomedial lobe but not folded; all pleopods with exopod longer than endopod. Uropod 0.5 times longer than pleotelson, peduncle 0.6 times longer than outer ramus, peduncle lateral margin without setae; rami not extending beyond pleotelson, apices narrowly rounded. Endopod apically not bifid, lateral margin proximally convex, distally concave. Exopod similar in length to endopod, apically not bifid, terminates without setae.
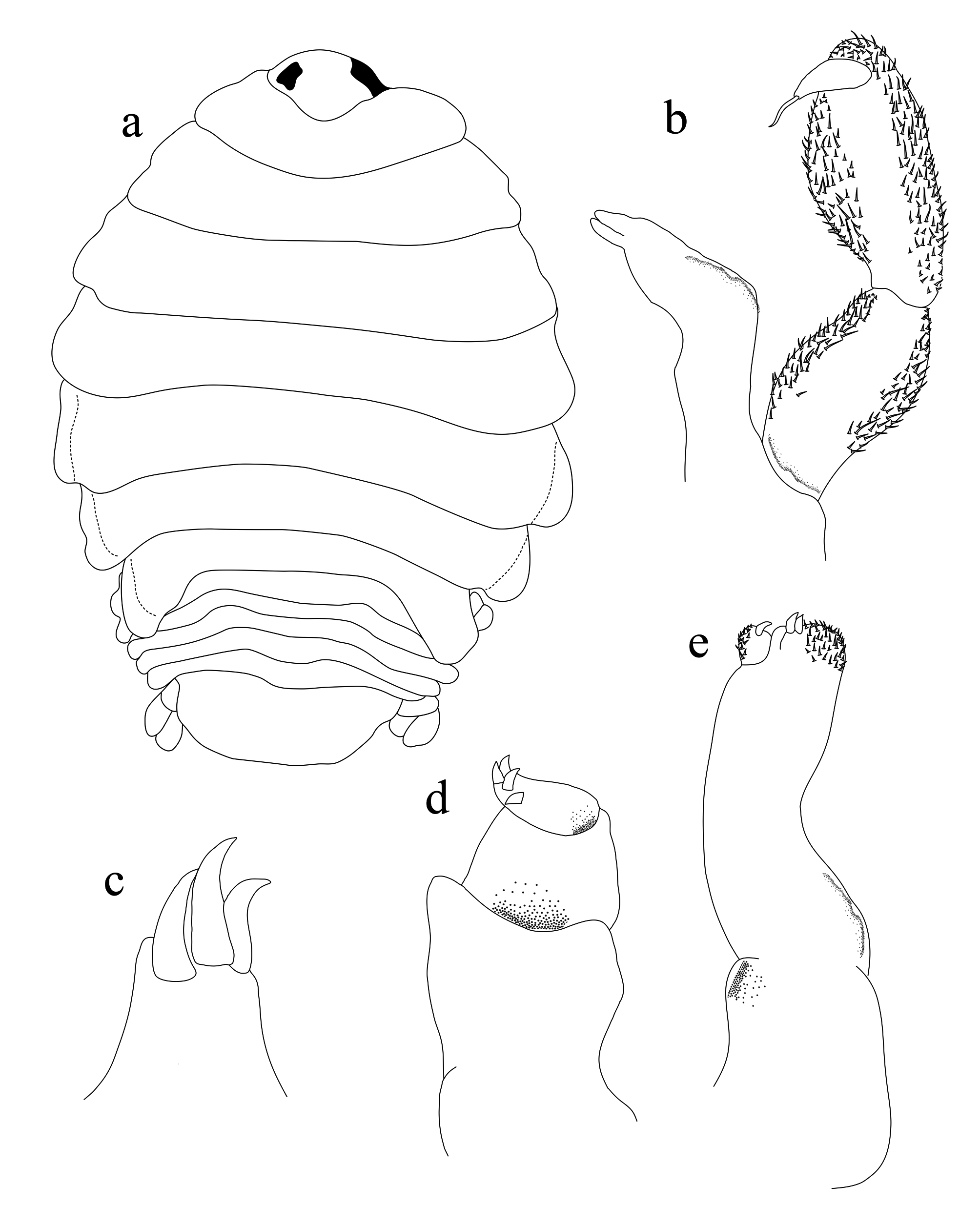
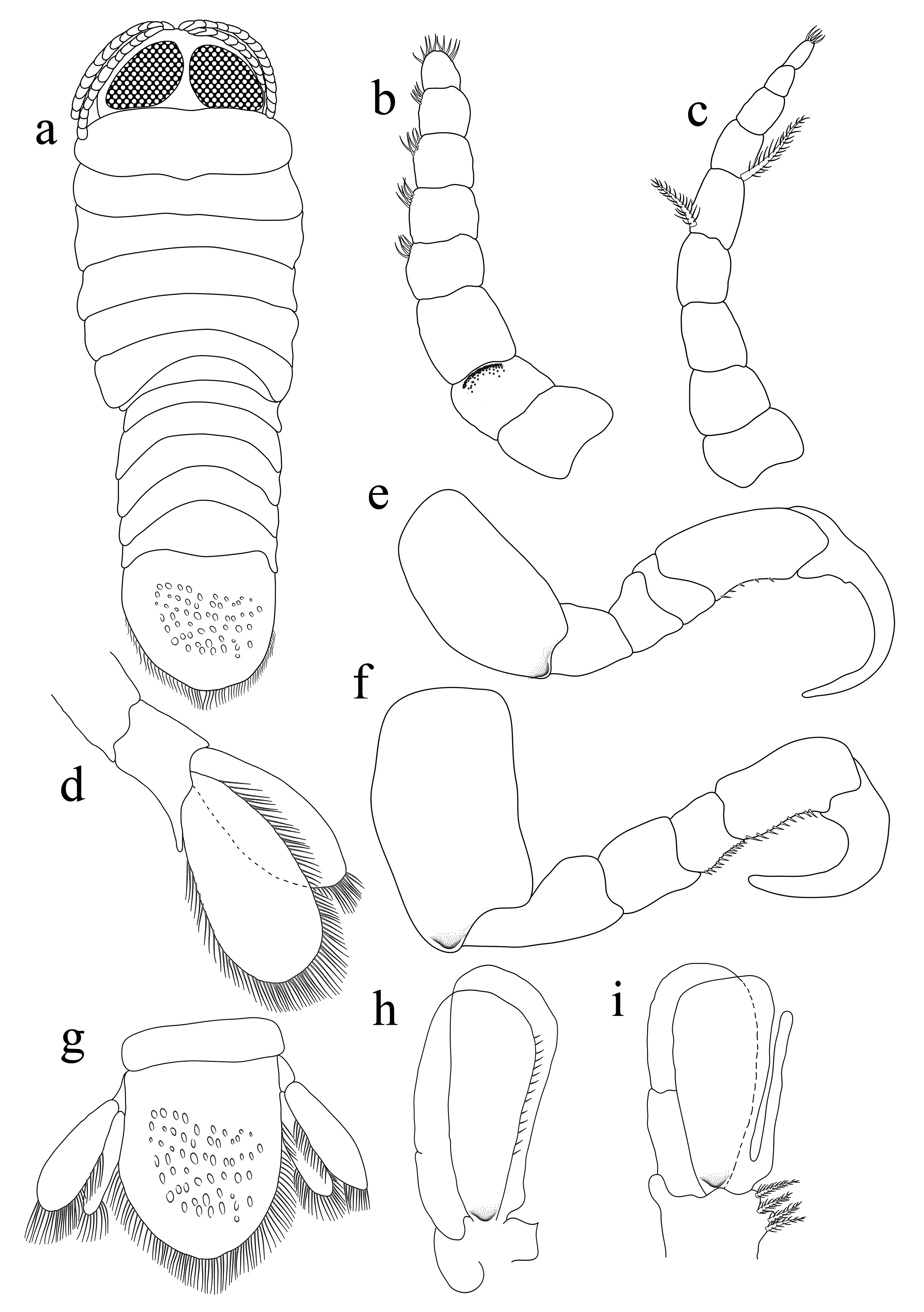
Male ( Fig. 5 View FIGURE 5 ): Body elongate, 2.2 times as long as wide. Pereonite 1 longest, anterolateral corners angular but not produced. Cephalon not immersed in pereonite 1, anterior margin more or less broadly rounded, eyes relatively larger and more visible than in female. Antennula stouter than antenna; with 8 articles, article 2 longest; articles 1– 3 slightly wider than other articles; distal margin of articles 4–7 with 3–5 setae; apex of article 8 curved with terminating in 4–6 short simple setae. Antenna composed with 9 articles, decreasing gradually in width, articles 4– 8 with 3–5 setae; apex of article 9 with terminating in 4–6 setae. Coxae 1–3 slightly conspicuous in dorsal view, coxal plates not reaching beyond posterior border of pereon. Pleonites subequal in width and length, pleonite 1 lateral margins partly concealed by pereonite 7; pleonites 2–5 visible. Pleotelson narrower than pleon, longer than wide. Pereopod 1 with propodus expanded, with 7 blunt spines, carpus with 2 spines, dactylus long and falcate. Pereopod 7 propodus with 3 spines; carpus with 1 spine. Uropods with elongate oblong rami, reaching tip of pleotelson. Penes prominent, 2.2 times as long as basal width.
Manca ( Fig. 6 View FIGURE 6 ): Body approximately 3.3 times as long as wide. Cephalon 1.3 times as wide as long, anterior margin rounded, not immersed in pereonite 1. Eyes ovate, conspicuous in dorsal view. Pereonite 1 longest, pereonite 3 widest, pereonites 2–4 subequal, pereonites 5–7 gradually decreasing in length, pereonites 4–7 decreasing in width. Pleonite 1 widest; pleonites 2–4, subequal to pereonite 7 length; pleonites 5–6 gradually decreasing in width. Pleotelson 1.1 times as long as wide; apical margin more or less rounded with many setae. Penes not developed.
Antennula stouter than antenna; 8 articles; article 3 longest; 1–3 articles slightly wider than other articles; distal margin of articles 4–7 with 3–5 setae; apex of article 8 curved, with 7–10 short simple setae. Antenna longer than antennula, composed of 9 articles, decreasing gradually in width, articles 5 and 6 with 1 plumose seta; article 9 with few terminal setae.
Pereopods 1 propodus with 5 small spines. Pereopod 7 propodus and carpus with several spines. Pleopods 1–3 with a long peduncle, lateral lobes not developed; peduncle of pleopods 4–5 with well-developed lateral and proximomedial lobes. Uropod rami extending beyond distal margin of pleotelson, subequal in length. Endopodite slightly short, rounded apically; exopodite with 25–35 setae, endopodite with 35–40 setae.
Colour: Both female and male appear white to yellowish–tan in alcohol. Manca appear mahogany brown, with chromatophores scattered posteriorly on the pereon and pleotelson.
Size: Ovigerous females: 9–12 mm; male 6–8 mm; manca 3.2–4.4 mm (present material).
Hosts: Known only from the family Clupeidae . Present material was collected from Amblygaster clupeoides , while Bunkley-Williams & Williams (1986), Williams & Bunkley-Williams (1994), Aneesh et al. (2016) and Praveenraj et al. (2017) recorded the species from Amblygaster sirm and Pillai (1954, 1964) recorded it from Amblygaster leiogaster .
Distribution: Southern coast of India ( Pillai, 1954, 1964; Aneesh et al. 2016, present study), Andaman Islands ( Praveenraj et al. 2017) and Thailand ( Williams & Bunkley-Williams 1994).
Prevalence and mean intensity: Out of 22 A. clupeoides , eight were found to be infested with R. circularis , with the prevalence and mean intensity 36% and 1.75 respectively.
Remarks. Ryukyua circularis can be recognised by its oval to rounded body shape; widest at pereonite 3–5; cephalon deeply immersed in pereonite 1; width of eyes 0.2 times width of cephalon; pleotelson anteriolateral margins weakly convex, posterior margin evenly rounded, without median point (refer to Fig. 2a View FIGURE 2 ); uropods half the length of pleotelson, rami not extending beyond the pleotelson. Pleonites 1–3 are partially overlapped by the posterolateral margin of pereonite 7. Ovigerous females are similar in appearance to non–ovigerous females (refer to Williams & Bunkley-Williams (1994)).
Males differ from females in having an elongated body, with a few chromatophores scattered towards the middle posterior margins of pereonite except pereonite 4; anterior margin of cephalon almost triangular, posterior margin not immersed in pereonite 1. The manca has eyes that cover nearly 90% of the whole cephalon, and has distinct chromatophores scattered on the pereonites and pleotelson, and appearing mahogany brown in colouration, in comparison to its adult female and male juvenile counterparts.
Ryukyua circularis and R. globosa have a near identical morphology of the cephalon deeply immersed in pereonite 1; body widest at pereonite 3–6; pereopods with rather a broad basis, elongate ischium and short propodus; and all pleopods are laminar. R. globosa can be distinguished from R. circularis by being greatly having a broader and rounded body shape; the posterolateral margin of pereonite 7 produced and overlap all pleonites (vs pleonite 1–3 partially overlapped by pereonite 7), pleotelson a slightly wider than long and having a rectangular (vs evenly rounded, without median point); a lobe present between bases of antenna 1, and antennae not extending beyond the posterior margin of the cephalon.
Host usage in the Cymothoidae is not random, and species of tropical Cymothoidae generally exhibit a narrow range of host use (see Bruce 1986; Trilles et al. 2011; Martin et al. 2015), often restricted to a family or related group of genera. The two species of Ryukyua are genus specific to the host Amblygaster , a small group of sardinellas from the family Clupeidae containing three valid species worldwide ( Eschmeyer et al. 2018): Amblygaster clupeoides (Bleeker's smoothbelly sardinella), Amblygaster leiogaster (Smoothbelly sardinella) and Amblygaster sirm (Spotted sardinella). Including this present study, R. circularis is present on all hosts, whereas R. globosa is commonly reported from A. sirm . Although it may not come as a surprise to find R. globosa on A. leiogaster , more reports are needed to clarify the host specificity and infection rate of this parasitic relationship.
With regards to geographical occurrence of this parasite (in relation to the availability of the host), both Ryukyua sp. have a Indo-West Pacific distribution, with both species reported from India (Rameshkumar et al. 2016, Praveenraj et al. 2017, current study); Japan for R. globosa and Thailand for R. circularis ( Williams & Bunkley-Williams 1994) .
Ryukyua circularis View in CoL attaches to the ventral part of the host branchial cavity, with reports of the species’ cephalon facing the anterior end of the host and the abdomen outwards toward the operculum in a lateral position ( Pillai 1954, Williams & Bunkley-Williams 1994). Four of the currently collected specimens were also dorsally positioned, although two were positioned with the ventral surface facing towards the operculum.
Since records of R. circularis View in CoL from the Indian sub–continent were made possible due to the collection of its fish host for subsistence and commercial use by local fisherman, it was an opportunity not to be missed to re-describe the species. Other reports that have further aided the ecological knowledge of the parasite include feeding behaviour ( Rameshkumar et al. 2013), molecular identification ( Praveenraj et al. 2017) and prevalence, mean intensity and abundance ( Aneesh et al. 2016). We hope clarity of its taxonomy and ecology (particularly hostspecificity) creates awareness on parasitic cymothoids and their potential harm towards the aquaculture and fisheries industry should an outbreak occur.
| CAS |
California Academy of Sciences |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Ryukyua circularis ( Pillai, 1954 )
| Vigneshwaran, P., Ravichandran, S. & Martin, Melissa B. 2018 |
Livoneca circularis
| Williams, E. H. Jr. & Bunkley-Williams, L. 1994: 160 |
| Bruce, N. L. 1990: 286 |
| Bunkley-Williams, L. & Williams, E. H. Jr. 1986: 213 |
| Pillai, N. K. 1954: 17 |