
Linyphiidae Blackwall, 1859
|
publication ID |
https://doi.org/ 10.5281/zenodo.212105 |
|
publication LSID |
lsid:zoobank.org:pub:B1A07FCF-EBF8-4006-8F9D-F6F56131B17C |
|
DOI |
https://doi.org/10.5281/zenodo.6176775 |
|
persistent identifier |
https://treatment.plazi.org/id/4C2DBE06-FF80-2A32-FF40-FAD3FED8F973 |
|
treatment provided by |
Plazi |
|
scientific name |
Linyphiidae Blackwall, 1859 |
| status |
|
Family Linyphiidae Blackwall, 1859 View in CoL View at ENA
Genus Pacifiphantes Eskov & Marusik, 1994 View in CoL
Pacifiphantes magnificus (Chamberlin & Ivie 1943) View in CoL Bathyphantes magnificus Chamberlin & Ivie 1943 ; Ivie 1969 P. magnificus Eskov & Marusik 1994
Type material.— Holotype female, CANADA, British Columbia, Vancouver Island, Lake Cameron [49.292 N 124.619 W, 189m el.], 13 Sept 1935, Coll: R. V. Chamberlin and W. Ivie. (not examined, the specimen was deposited in the AMNH collection but so far we have not been able to locate it).
Material examined.— CANADA: BRITISH COLUMBIA: 1F, Terrace (54.31N, 128.36W) AMNH-Ivie (1969); 1F, Inverness (54N, 130W) AMNH; USA: ALASKA: 1M, Prince of Wales Island, Forest Rd. 2056100 (55.894N, 133.071W) UAM 100046555; 1F, Sumez Island, Port Santa Cruz (55.253N, 133.421W) UAM 100046066; 1F, Lituya Bay, N. of Echo Creek (58.709N, 137.702W) Burke230A; 3F, Lituya Bay, N. of Echo Creek (58.714N, 137.694W) Burke245; OREGON: 1F, 5 mi E of Detroit (44.44N, 122.05W) AMNH-Ivie(1969); WASHINGTON: 1F, King Co, N. slope Tiger Mtn. (47.471N, 121.933W) Burke168d; 1F, 5 mi E. of McCleary (47.03N, 123.1W) AMNH-Ivie (1969);
Diagnosis.— Specimens of Pacifiphantes magnificus can be separated from all other Pacifiphantes species by the embolus forming a single loop, located on the prolateral axis of the palp ( Figure 1 View FIGURES 1 – 7 ), and the copulary ducts forming a large anterior area, often appearing folded or twisted ( Figures 5 and 6 View FIGURES 1 – 7 ).
Species Database Specimen Record Agnyphantes arboreus BOLD SPIAL070
Agnyphantes arboreus BOLD SPIAL195
Agnyphantes arboreus BOLD SPIAL271
Australolinyphia remota GenBank FJ838649 View Materials
Bathyphantes brevipes BOLD SPIAL198
Bathyphantes brevipes BOLD SPIAL210
Bathyphantes brevipes BOLD SPIAL222
Bathyphantes brevipes BOLD SPIAL281
Bathyphantes brevipes BOLD SPICH1097 Bathyphantes brevipes BOLD SPICH426
Bathyphantes brevipes BOLD SPICH499
Bathyphantes brevipes BOLD SPICH796
Bathyphantes brevipes BOLD SPICH797
Bathyphantes brevipes BOLD SPICH803
Bathyphantes brevipes BOLD SPICH812
Bathyphantes brevis BOLD SPICH255
Bathyphantes brevis BOLD SPICH257
Bathyphantes brevis BOLD SPICH263
Bathyphantes brevis BOLD SPICH501
Bathyphantes brevis BOLD SPICH533
Bathyphantes brevis BOLD SPICH546
Bathyphantes canadensis BOLD BBCAN388 Bathyphantes canadensis BOLD SPICH292
Bathyphantes canadensis BOLD SPICH296
Bathyphantes gracilis GenBank FJ838650 View Materials
Bathyphantes gracilis GenBank FJ899797 View Materials
Bathyphantes gracilis GenBank FJ899798 View Materials
Bathyphantes orica BOLD SPIAL025
Bathyphantes orica BOLD SPIAL037
Bathyphantes orica BOLD SPIAL061
Bathyphantes orica BOLD SPIAL073
Bathyphantes pallidus BOLD BBCAN109 Bathyphantes pallidus BOLD BBCAN189 Bathyphantes pallidus BOLD BBCAN245 Bathyphantes pallidus BOLD BBCAN738 Bathyphantes pallidus BOLD BBCAN753 Bathyphantes pallidus BOLD SPICH485
Bathyphantes pallidus BOLD SPICH799
Bathyphantes pallidus BOLD SPISH038
Bathyphantes pallidus GenBank AY944732 View Materials
Bathyphantes tongulensis GenBank EF128165 View Materials
Bathyphantes tongulensis GenBank EF128166 View Materials
Diplostyla concolor BOLD BBCAN428 Diplostyla concolor GenBank FJ838651 View Materials
Haplinis diloris GenBank FJ838657 View Materials
Incestophantes washingtoni BOLD SPICH222
Incestophantes washingtoni BOLD SPICH273
Incestophantes washingtoni BOLD SPICH289
Incestophantes washingtoni BOLD SPICH290
Incestophantes washingtoni BOLD SPICH329
Incestophantes washingtoni BOLD SPICH352
Incestophantes washingtoni BOLD SPICH369
Incestophantes washingtoni BOLD SPICH374
Incestophantes washingtoni BOLD SPICH996
Continued on next page... Description.— Total body length, 3.50–3.90. Carapace low, lacking cephalothoracic sucli. Anterior median eyes smallest, less than one diameter apart, other eyes approximately equal in size. Lateral eyes adjacent, posterior median eyes less than one diameter apart. Posterior median eyes 1.5 diameters from lateral eye rows. Posterior eye row slightly recurved. Chelicerae with three prolateral and no retrolateral teeth. Leg lengths I, II, IV, III. Male leg I about seven times carapace length, female leg I about five time carapace length. TmI ~0.25, TmIV absent, all tibiae with two dorsal spines and no metatarsal trichobothria. Legs with distinct dusky bands.
Male (n=1): Total length = 3.50, carapace length = 1.70, carapace width = 1.20, TmI = 0.25, TmIV absent. Chaetotaxy: F I, 0-1-0-0; F II-IV, 0-0-0-0; Pt I-IV, 1-0-0-0; Ti I, 2-1-1-0, Ti II, 2-0-1-0, Ti III-IV, 2-0-0-0; Mt I- IV, 0-0-0-0. Carapace smooth, yellow. Dusky lines extending off lateral eye rows to fovea. Dark triangular patches extending anteriorly and posteriorly of all eyes. Lateral margins dusky with lines radiating from fovea for each coxa. Sternum dusky. Legs with terminal and medial dusky bands. Abdomen dark with six light chevrons and two light patches adjacent to the heart mark ( Figure 7 View FIGURES 1 – 7 ). Venter dusky. Palp with prominent embolus originating from ventral prolateral edge, making almost a complete loop along the lateral axis, exterior of the radix/suprategular assembly ( Figure 1 View FIGURES 1 – 7 ). Radix and suprategulum fused along entire length of radix, without tailpiece. Suprategular apophysis large, hook shaped ( Figure 1 View FIGURES 1 – 7 ). Lamella short and wide, located medially on prolateral side of palp. Tegulum smooth with distal globular apophysis, somewhat wrinkled. Paracymbium small, cymbium with ectal rectangular expansion extending ventrally by paracymbium ( Figure 2 View FIGURES 1 – 7 ).
Female (n=6): Total length = 2.92–3.90, carapace length = 1.33–1.70, carapace width = 1.20–1.33, TmI = 0.24–0.26, TmIV absent. Chaetotaxy: Same as male except; F I, 1-1(2)-0-0. Coloration similar as male. Pedipalp with tarsal claw. Epigynum lacking a ventral plate scape. Parmula low and wide with pit located along posterior edge, often projecting posteriorly in a "V" shape. Atrium roughly triangular ( Figures 3 and 4 View FIGURES 1 – 7 ). Large bursal plate ( Figure 5 View FIGURES 1 – 7 ). Copulatory opening located on lateral margin of atrium. Copulatory ducts extend laterally from atrium, ventrally of the spermathecae, and proceed anteriorly, expanding in the anterior region, then often folding as they curve medially to the spermathecae. Fertilization ducts extend posteriorly along widest lateral edge of atrium ( Figure 6 View FIGURES 1 – 7 ).
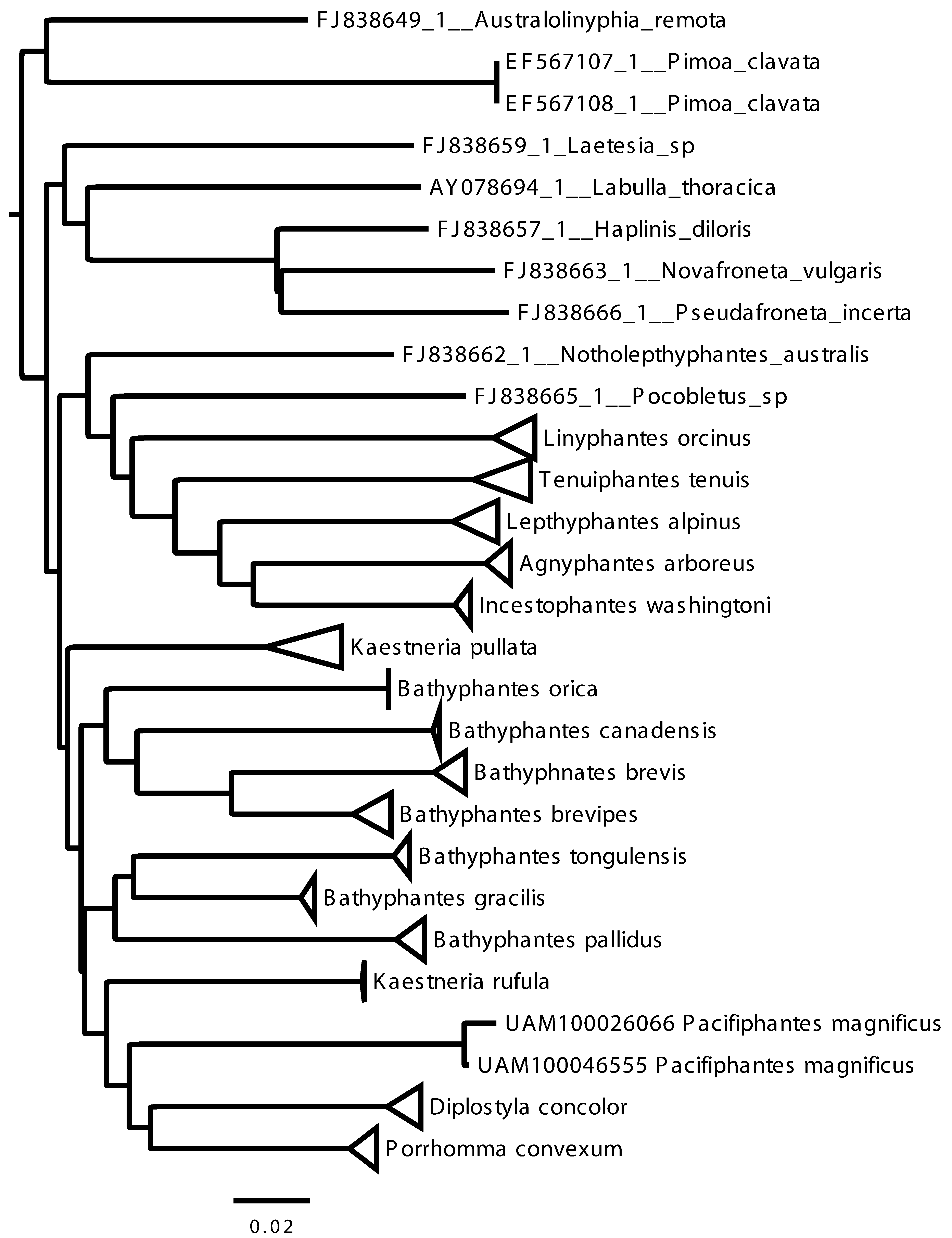
Results of the distance analysis ( Figures 8 View FIGURE 8 ) clearly show the monophyly of the independently collected P. magnificus specimens in relation to other linyphiid taxa (specimens UAM100046555 and UAM100046066). Additionally, the low amount of genetic difference between the two P. magnificus specimens, 0.3%, is a indicator that these are conspecifics. The species clade sister to Diplostyla and Porrhomma within the general Bathyphantes lineage, which is separate from both the Lepthyphantes and Linyphantes lineages. Both Bathyphantes and Kaestneria appear polyphyletic in the NJ analysis limiting the use of these DNA barcode results to direct the generic placement of Pacifiphantes pacificus .
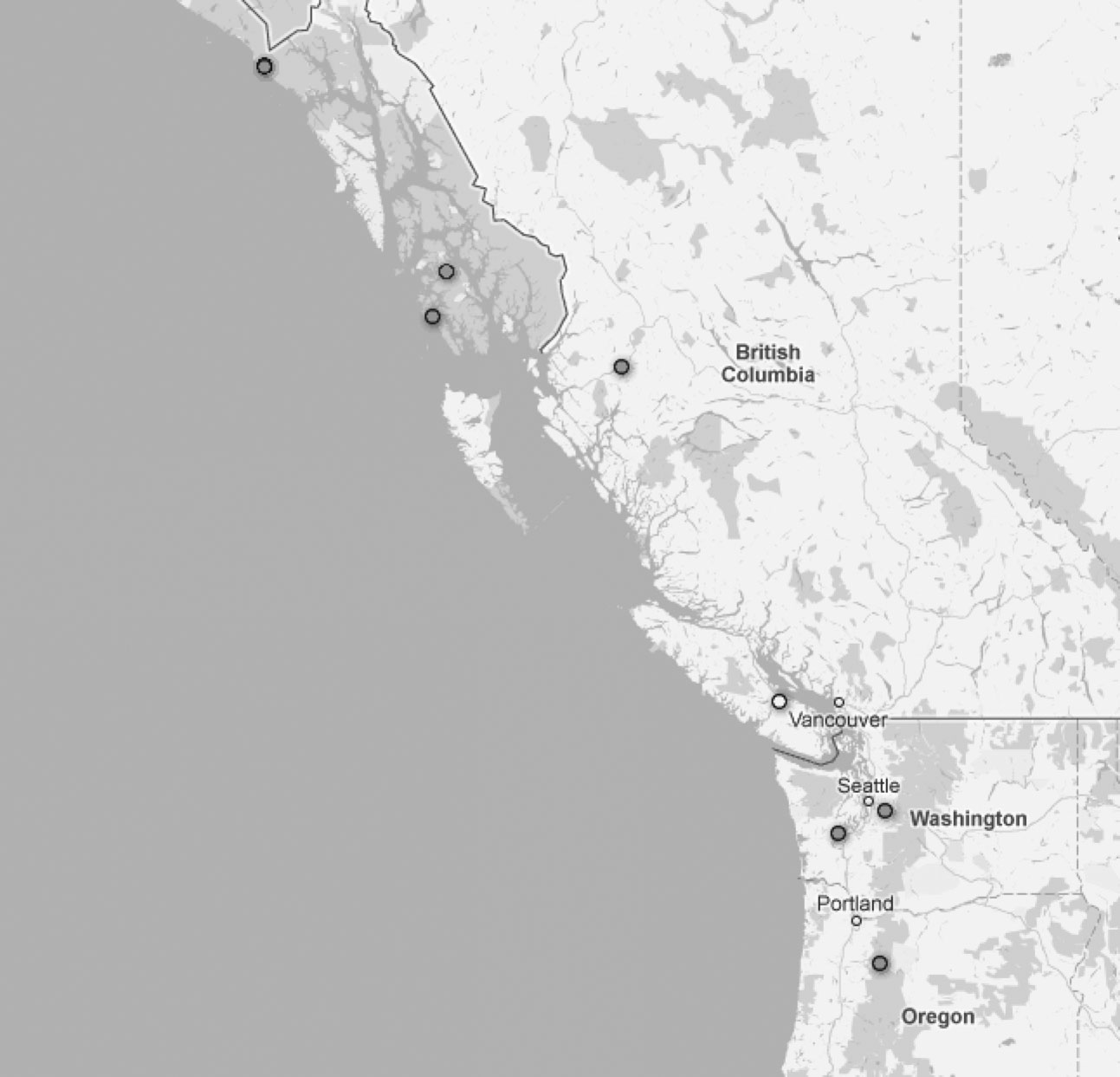
Distribution.— Known only from the Pacific Coast region of North America from Oregon north to Alaska, USA ( Figure 9 View FIGURE 9 ).
Habitat.— It would appear from the few specimens collected, the difficulty in collecting the single male specimen, and fairly extensive collecting done along the Pacific Coast ( Mann & Gara 1980, Crawford 1988, Bennett 2001, Slowik 2006) that this is a rare species. There is nothing specific known about the preferred habitat in which this species may occur as many of the specimens are known from pitfall traps or casual collections. Based on the locations and descriptions of the specimens examined, this species occurs in closed canopy mature forests. Specimens examined were collected from shrubs along creeks, and overwinter pitfall traps.
Discussion.— This morphological examination has shown that the placement of P. magnificus in Pacifiphantes is likely incorrect. The generic designation for Pacifiphantes comments that males have a "very short embolus and suprategular apophysis" ( Eskov & Marusik 1994: 49), in which specimens of P. magnificus have a long embolus, and large hook shaped suprategular apophysis. Additionally the characters used for the female designation of Pacifiphantes are brief and refer only to the delimited parmula ( Eskov & Marusik 1994).
Although the generic designation of Pacifiphantes excludes P. magnificus , clear placement based on morphological characters alone is not obvious. P. magnificus shares many epigynal characters with Porrhomma . specifically the shape of the atrium and the location and shape of the copulary ducts and spermathecae, for example compare Figure 6 View FIGURES 1 – 7 with Eskov & Marusik 1994 figure 48 of Porrhomma longjiangensis Zhu & Wang 1983 (= P. rakanum Yaginuma & Saito 1981 ).
Although the females may show similarities to Porrhomma , males of P. magnificus have a morphology not typical for the genus, including a reduced radix and a longer embolus free of a terminal sheath (or embolic membrane). Ivie's (1969) hypothesis that P. magnificus was sister to Bathyphantes approximatus (O. Pickard- Cambridge 1871) is also questioned as the morphology of the palp, particularly the embolus and radix/ suprategulum assembly, differ significantly. Additionally, examination of the internal epigynal structures of the two species fail to support this grouping as B. approximatus has highly coiled copulatory ducts and anteriorly placed spermathecae, more typical of the the genus Microlinyphia (e.g., see Blauvet 1936: figs. 72 and 77). Additionally P. magnificus lacks cephalothoracic sulci, which have been found on Porrhomma , Bathyphantes , and Pacifiphantes species ( Hormiga 1999).
Thus, combined characters from the male and female morphology could be used to associate or dissociate P. magnificus to or from many of the other genera contained in the Porrhomma group ( Millidge 1977), or Bathyphantes clade ( Hormiga 1999, Arnedo et al. 2009). Our DNA barcode results are insufficient to provide a generic placement for P. magnificus . Currently relatively few species from the Bathyphantes lineage are available for comparison. Additionally, our results find several genera polyphyletic suggesting that more data are needed to address these taxonomic problems. Given the data at hand, the placement of P. magnificus remains elusive and we feel that at this time it is best to leave the spider in Pacifiphantes until a more thorough analysis can be done.
Pacifiphantes magnificus males will key out in the Spiders of North America ( Ubick et al. 2005) in the Linyphiidae generic key to couplet 25, Poeciloneta . However, Pacifiphanytes magnificus lack trichobothria on the metatarsus. Continuing with the key leads to couplet 27, in which the genus differs from either Porrhomma or Kaestneria by the embolus forming almost a complete loop. Females are included in the key under couplet 222, Pacifiphantes .

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
Linyphiidae Blackwall, 1859
| Slowik, Jozef & Blagoev, Gergin A. 2012 |
Pacifiphantes
| Eskov & Marusik 1994 |
P. magnificus
| Eskov & Marusik 1994 |
Pacifiphantes magnificus
| Chamberlin & Ivie 1943 |
Bathyphantes magnificus
| Chamberlin & Ivie 1943 |