
Daspletosaurus wilsoni, Warshaw & Fowler, 2022
|
publication ID |
https://doi.org/10.7717/peerj.14461 |
|
DOI |
https://doi.org/10.5281/zenodo.7372908 |
|
persistent identifier |
https://treatment.plazi.org/id/4D6C87E9-FF90-FF8B-74B2-46221031F8DB |
|
treatment provided by |
Jonas |
|
scientific name |
Daspletosaurus wilsoni |
| status |
sp. nov. |
D. wilsoni sp. nov.
Etymology
wilsoni, Latinization of “Wilson,” after John Wilson, the discoverer of the holotype specimen.
Holotype
BDM 107 , preserving a partial disarticulated skull and postcranium, including both premaxillae, a right maxilla, jugal, lacrimal, quadrate, quadratojugal, dentary, and splenial, and a left postorbital and squamosal. Also preserved are partial cervical, sacral, and caudal series, a rib, a chevron, and a first metatarsal. Cranial bones are very finely preserved, with intricate and detailed surface textures especially on the maxilla and postorbital, with teeth preserved in the maxilla, dentary, and one premaxilla. The sacral and caudal centra are preserved in a heavy and hard concretion and are not yet prepared. The holotype specimen is stored in the collections of the Badlands Dinosaur Museum ( BDM) in Dickinson, North Dakota.
Geological Setting
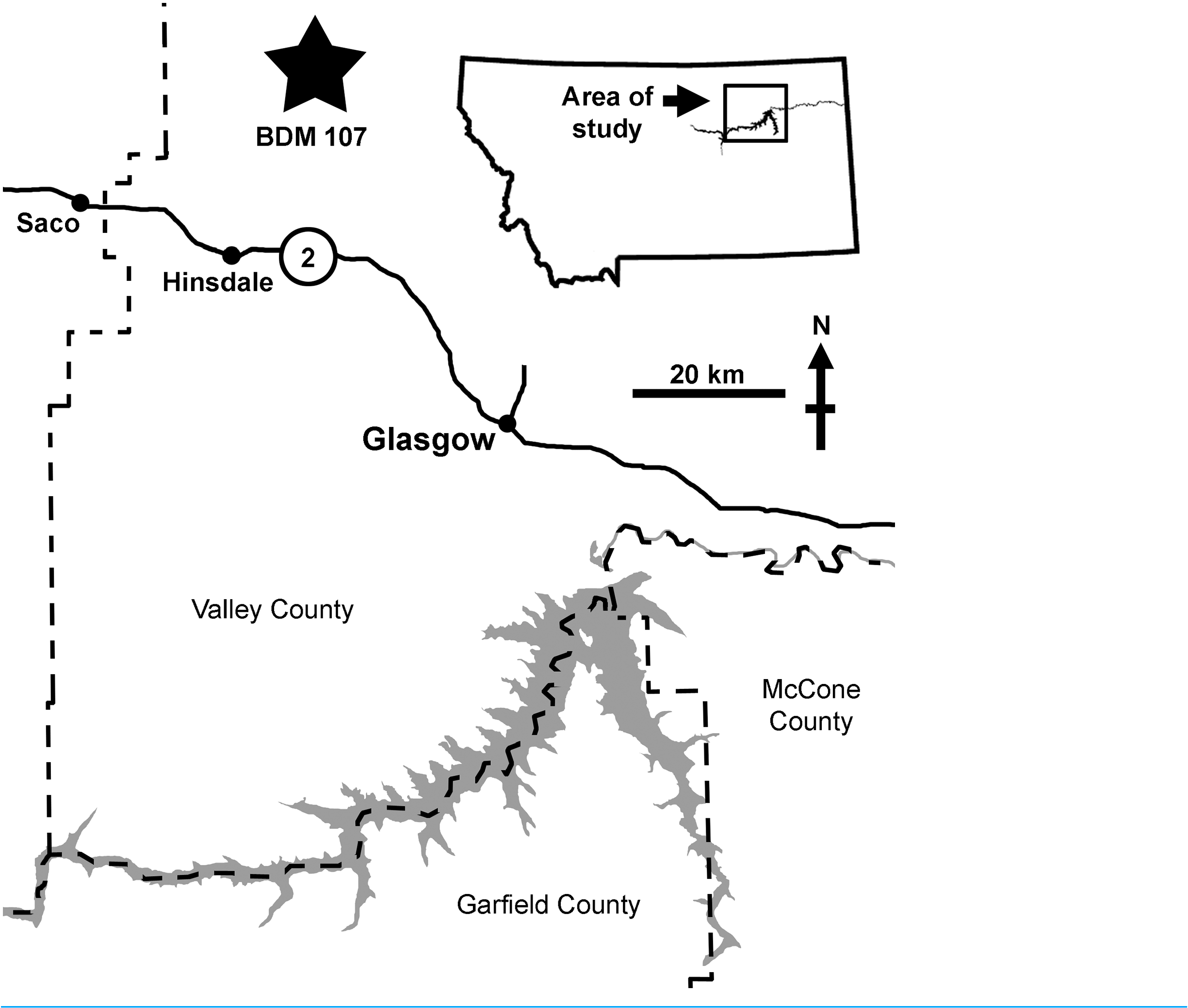
BDM 107 was recovered from the site “Jack’ s B2,” discovered in 2017 by John Wilson in exposures of the Judith River Formation near Glasgow (Valley County, MT, USA). This is significantly further east than classic ‘Judith’ localities ( Fig. 1 View Figure 1 ), and is sedimentologically atypical, representing distal floodplain and delta sediments deposited during the maximum Campanian regression of the Western Interior Seaway. Here, the Judith River Formation is up to ~48 m thick, with the “Jack’ s B2” site occurring ~30 m below the contact with the overlying Bearpaw Shale.
Precise stratigraphic placement of this easternmost Judith is currently unclear, although an age of ~76.5 Ma seems most likely, which would correlate in time with the lower to middle part of the Dinosaur Park Formation, Alberta ( Eberth, Currie & Koppelhus, 2005; Fowler, 2017). A youngest age limit of 75.64 Ma ( Ogg & Hinnov, 2012) is delineated by ammonites tentatively identified as Didymoceras stevensoni (J. Slattery, 2020, personal communication) collected by BDM from local outcrops of the overlying Bearpaw Shale (although these were not at the base of the Bearpaw, so older ammonite specimens may be encountered during future prospecting). At present, more precise stratigraphic position can be inferred from the timing of the maximum regression of the Western Interior Seaway during the Campanian (correlated with the R8 regression of Kauffman (1977) and Rogers et al. (2016)). In Alberta and Saskatchewan, the Foremost, Oldman, and Dinosaur Park formations represent early to late subcycles (respectively) of the R8 regression, and of these, the Foremost (~80.5–79.5 Ma) and lower Oldman (~79.5–79.0 Ma; and regional equivalents) are restricted to the west (Alberta and west central Montana), and did not extend as far east as Saskatchewan or our study area in eastern Montana ( Eberth, Currie & Koppelhus, 2005). During late R8, the upper Oldman (~77.5–77.0 Ma) and Dinosaur Park (~76.9–76.0 Ma) Formations were deposited much further to the east, with the lowermost Dinosaur Park recording the R8 maximum regression at ~76.9–76.4 Ma ( Eberth, Currie & Koppelhus, 2005; Fowler, 2017). This correlates well with the Judith River Formation of Montana, where Rogers et al. (2016) show the maximum regression of R8 occurring shortly before 76.2 Ma, based on radiometric dates acquired either side of the mid-Judith discontinuity. As such, it seems likely that the study section corresponds in age to the lower to middle part of the Dinosaur Park Formation (although not necessarily lithostratigraphically correlated). A radiometric analysis of a newly discovered volcanic ash is currently underway, and it is hoped that this will provide definitive stratigraphic placement.
Regardless of the precise age of BDM 107, it can be expected to lie intermediate stratigraphically between D. torosus (known from the upper Oldman Formation, ~77.0 Ma; Paulina Carabajal et al., 2021) and D. horneri (known from the Two Medicine Formation, ~75.0 Ma; Carr et al., 2017).
Diagnosis
D. wilsoni can be assigned to Daspletosaurus based on the following characteristics: extremely coarse subcutaneous surface of the maxilla with no elevated ridges or corresponding fossae ( Carr et al., 2017; Voris et al., 2020); cornual process of the postorbital approaching the laterotemporal fenestra ( Carr et al., 2017); dorsal postorbital process of the squamosal terminating caudal to the rostral margin of the laterotemporal fenestra ( Carr et al., 2017; Voris et al., 2019); and extremely coarse symphyseal surface of the dentary ( Voris et al., 2020).
D. wilsoni possesses a single autapomorphy: a rostrocaudally elongate and dorsoventrally narrow mylohyoid foramen of the splenial (this foramen is much deeper in other Daspletosaurus, Carr et al., 2017 ; see below), and can additionally be diagnosed by a unique combination of ancestral and derived Daspletosaurus characteristics. D. wilsoni and D. torosus share a pneumatic inflation of the lacrimal reaching the medial edge of the bone (this inflation does not reach the medial edge of the bone in the holotype of D. horneri , but this may represent an allometric, ontogenetic, or taphonomic bias; E. A. Warshaw, 2022, unpublished data; Carr et al., 2017), cornual process of the postorbital approaching the laterotemporal fenestra (this process terminates much more rostrally relative to the fenestra in D. horneri , contra Carr et al., 2017; see below), cornual process of the postorbital subdivided into two distinct processes (this subdivision is absent in D. horneri, E. A. Warshaw, 2022 , personal observations; see Description); prefrontal oriented rostromedially (determined from the angle of the prefrontal articular surface on the lacrimal of the holotype of D. wilsoni , which does not preserve a prefrontal; the prefrontal of D. horneri is oriented mediolaterally), pneumatic excavation of the squamosal that does not undercut its rostromedial margin (entire margin undercut in D. horneri ; Carr et al., 2017), and quadratojugal lacking a pneumatic foramen in its lateral surface (although the presence of this foramen is highly intraspecifically variable in both D. horneri and Tyrannosaurus , such that further discoveries of D. wilsoni individuals may reveal its presence in this taxon; Carr et al., 2017; Carr, 2020). D. horneri and D. wilsoni share, to the exclusion of D. torosus , a premaxillary tooth row oriented entirely mediolaterally, such that all but one premaxillary tooth is concealed in lateral view (rostromedial orientation in D. torosus and less derived tyrannosaurids), antorbital fossa of the maxilla terminating at the rostral limit of the external antorbital fenestra (this fossa extends ahead of this boundary onto the subcutaneous surface of the maxilla in D. torosus and less derived tyrannosaurids; Carr et al., 2017; E. A. Warshaw, 2022, unpublished data), rostrodorsal ala of the lacrimal inflated (uninflated in D. torosus and less derived tyrannosaurids), ventral ramus of the lacrimal longer than the rostral ramus (determined largely by the height of the postorbital bar in the reconstructed skull, given that the ventral ramus is largely unpreserved in the holotype of D. wilsoni ; the rostral ramus of the lacrimal is longer than the ventral ramus in D. torosus ; Carr et al., 2017), short cornual process of the lacrimal (tall in D. torosus , although this process is taller in D. wilsoni than D. horneri and may best be described as intermediate between the previously named species of this genus; Carr et al., 2017), and dorsal quadrate contact of the quadratojugal visible in lateral view (concealed in D. torosus and less derived tyrannosaurids).
Description
Given the wealth of detailed osteologies describing tyrannosaurine specimens ( e.g., Carr (1999); Brochu (2003) and Hurum & Sabath (2003)), our description of the holotype of D. wilsoni places heavy emphasis on characteristics (or combinations of characteristics) unique to this specimen, as well as those that are otherwise taxonomically or phylogenetically informative within Tyrannosaurinae, so as to avoid the reiteration of plesiomorphic tyrannosaurine morphologies (or synapomorphies of Daspletosaurus ) already described by previous authors ( e.g., Carr et al. (2017); Voris et al. (2019) and Voris et al. (2020)).
Ontogenetic Stage of BDM 107
In order to facilitate comparison with other tyrannosaurine individuals of equivalent ontogenetic stages (and in doing so, to avoid the misattribution of a phylogenetic signal to ontogenetically derived characteristics), brief comment is warranted on the ontogenetic stage represented by BDM 107; two lines of evidence suggest that this specimen is of advanced ontogenetic age. Firstly, BDM 107 is among the largest known Daspletosaurus individuals (articulated skull length 105 cm; D. torosus holotype CMN 8506 skull length 104 cm, Voris et al., 2019; D. horneri holotype MOR 590 skull length 89.5 cm, Carr et al., 2017). Although Carr (2020) criticized the use of size as an indicator of ontogenetic status in Tyrannosaurus , this criticism was based on the absence of a correlation between size and maturity among adult individuals; all the largest specimens of this genus were unambiguously recovered as adult by Carr’ s (2020) analysis ( i.e., within the final stages of ontogenetic development), such that this feature remains ontogenetically informative in distinguishing adults from juveniles and subadults. Secondly, BDM 107 displays several morphologies known otherwise to characterize mature tyrannosaurines, including a deeply scalloped maxilla-nasal suture ( Carr & Williamson, 2004; Carr, 2020), a maxillary fenestra positioned rostrally within the antorbital fossa ( Carr, 2020), a cornual process of the lacrimal inflated and positioned dorsal to the ventral ramus ( Carr, 1999; Currie, 2003; Carr, 2020), and a grossly exaggerated cornual process of the postorbital ( Carr, 1999; Currie, 2003; Voris et al., 2019; Carr, 2020). The totality of this evidence supports an adult ontogenetic stage or later for BDM 107 (adult sensu Carr, 2020; ontogenetic Stage 4 sensu Carr, 1999); this hypothesis may be tested in future work through histological analysis and/ or comparison with further discoveries of D. wilsoni individuals of different ontogenetic stages, both of which lie outside of the scope of the present study.
Premaxilla
The premaxillae of D. wilsoni are similar to those of D. horneri ( Carr et al., 2017, Fig. 1 View Figure 1 ; Fig. 2 View Figure 2 ), Tarbosaurus ( Hurum & Sabath, 2003, Fig. 3 View Figure 3 ), and Tyrannosaurus ( Brochu, 2003, Fig. 4 View Figure 4 ) in that the alveolar row is oriented largely mediolaterally, such that the rostrum of the skull is broad and the labial surfaces of the premaxillary teeth face rostrally.
In Tyrannosaurus and similarly derived tyrannosaurines ( Tarbosaurus and D. horneri ), the premaxillary teeth largely overlap each other in lateral view such that only the distalmost tooth is clearly visible; the same would be true of the holotype of D. wilsoni , were more than a single premaxillary tooth preserved within its socket. Conversely, the premaxillary tooth row of D. torosus and less derived tyrannosauroids is oriented rostromedially, such that multiple teeth are clearly visible in lateral view ( Voris et al., 2019, Fig. 6 View Figure 6 ).
Although previous authors have regarded a mediolaterally oriented premaxillary tooth row as a synapomorphy of Tyrannosauridae or more inclusive groups ( e.g., Carr et al., 2017: character 15), this is in error; mature specimens of Gorgosaurus (UALVP 10, Voris et al., 2022, Fig. 1 View Figure 1 ; AMNH 5458, Matthew & Brown, 1923, Fig. 2 View Figure 2 ) and Qianzhousaurus (GM F10004, Foster et al., 2021, Fig. 2 View Figure 2 ), have rostromedially oriented premaxillary tooth rows such that in specimens with preserved teeth, all premaxillary teeth are visible in lateral view (although all tyrannosaurids do have premaxillary tooth rows oriented more medially than basal tyrannosauroids; this is the phylogenetic signal recorded in character 15 of Carr et al. (2017)). Comparison with other tyrannosaurids is hampered by the absence of preserved premaxillae and/or published descriptions of this element for several species ( e.g., Thanatotheristes, Voris et al., 2020 ; Dynamoterror , McDonald, Wolfe & Dooley, 2018; Nanuqsaurus, Fiorillo & Tykoski, 2014 ; Lythronax and Teratophoneus, Loewen et al., 2013 , for which all published specimens lack premaxillae); however, D. wilsoni and more derived tyrannosaurines ( D. horneri , Tarbosaurus , Tyrannosaurus ) represent the greatest exaggeration of the medial inclination of the premaxillary tooth row among tyrannosaurids for which comparative material is available (although this condition, with only one clearly visible premaxillary tooth in lateral view, is present in at least one Gorgosaurus: TCMI 2001.89.1, Voris et al., 2022, Fig. 10 View Figure 10 ). D. torosus is intermediate between the (presumably) ancestral rostromedial orientation and the mediolateral condition of later Daspletosaurus species; two to three premaxillary teeth are visible in lateral view in the holotype specimen, CMN 8506 ( Carr & Williamson, 2004, Fig. 6 View Figure 6 ; Voris et al., 2019, Fig. 6 View Figure 6 ).
It should be noted that the orientation of the premaxillary tooth row is not necessarily equivalent to the orientation of the premaxillae themselves. In Tyrannosaurus AMNH 5027, for example, the premaxillae appear to be rostromedially oriented in dorsal view ( Carr & Williamson, 2004, Fig. 7 View Figure 7 ); however, the premaxillary alveoli are mediolaterally arranged when viewed ventrally (E. A. Warshaw, 2022, personal observations; Osborn, 1912, Fig. 5A View Figure 5 ; Molnar, 1991, Fig. 9A View Figure 9 ).
The taxonomic utility of this character is a hypothesis that will require further testing as individuals of D. wilsoni and other tyrannosaurids with preserved premaxillae are discovered; notably, two specimens previously referred to D. torosus display the derived condition (mediolateral orientation), sharing it with D. wilsoni and more derived tyrannosaurines: FMNH PR308 ( Matthew & Brown, 1923, Fig. 5 View Figure 5 ; Carr, 1999, Fig. 1 View Figure 1 ) and TMP 2001.36.1 ( Voris et al., 2019, Fig. 6 View Figure 6 ). If these individuals were to represent D. torosus , the distinction between this species and D. wilsoni in the orientation of the premaxillary tooth row would be heavily undermined; however, both of these specimens have previously been noted as belonging to a novel taxon from the Dinosaur Park Formation (FMNH PR308, Currie, 2003; TMP 2001.36.1, Paulina Carabajal et al., 2021). Therefore, although relevant comparisons will be made with these specimens hereafter, they will be considered separately from D. torosus (and will be referred to below as the Dinosaur Park taxon). A precise taxonomic designation for these specimens is reserved for future work in accordance with comments by previous authors ( Currie, 2003; Paulina Carabajal et al., 2021).
There is a small (~2 cm diameter) indentation in the nasal process of the right premaxilla of BDM 107; this is most likely pathological, as it is irregular in form and not present on the left premaxilla.
Maxilla
As in other Daspletosaurus , the subcutaneous surface of the maxilla in D. wilsoni is densely covered in anastomosing sulci extending from neurovascular foramina ( Carr et al., 2017; Voris et al., 2020; Fig. 3 View Figure 3 ). The degree of sculpturing of this surface in BDM 107 is similar to CMN 8506 ( D. torosus ), although in the former, there is no smooth region rostral to the external antorbital fenestra indicating a rostral continuation of the antorbital fossa as D. torosus and alioramins ( Carr et al., 2017). As in Thanatotheristes and other Daspletosaurus species, the shallow excavations that characterize the maxillae of the most derived tyrannosaurines ( Zhuchengtyrannus , Tyrannosaurus , Tarbosaurus ; Hone et al., 2011; Voris et al., 2020) are absent from the holotype maxilla of D. wilsoni . Also absent are the textural ridges present on the maxillae of Zhuchengtyrannus ( Hone et al., 2011) , Tarbosaurus , Tyrannosaurus , and Thanatotheristes ( Voris et al., 2020) , but not any Daspletosaurus species.
The rostral end of the maxilla of BDM 107 is bowed subtly medially towards its contact with the premaxilla and nasal; this may be a structural consequence of the greater medial inclination of the premaxillary tooth row (see above), as a similar condition characterizes D. horneri (MOR 590, E. A. Warshaw, 2022, personal observations), Tarbosaurus ( Hurum & Sabath, 2003, Fig. 15), and Tyrannosaurus (MOR 008, MOR 980, E. A. Warshaw, 2022, personal observations). Tyrannosaurids with more rostromedially inclined premaxillary tooth rows lack this bowing ( e.g., D. torosus CMN 8506, J. T. Voris, 2022, personal communication).
The maxilla of BDM 107 is irregular relative to other species of Daspletosaurus in that it is proportionally elongate, being 64.1 cm in length and 24.8 cm in height (ratio of length to height = 2.6). This bone is 58.6 cm long rostrocaudally and 27.5 cm tall dorsoventrally in the holotype of D. horneri (ratio of length to height = 2.1; MOR 590, Carr et al., 2017). Given the broad range of variation in the proportions of this element in other tyrannosaurine species for which larger sample sizes are known ( e.g., Tyrannosaurus ; Carpenter, 1990; Paul, Persons & Van Raalte, 2022; E. A. Warshaw, 2022, personal observations), this characteristic was not included as an autapomorphy of D. wilsoni . Consistency in this trait across further discoveries of D. wilsoni individuals may require a reevaluation of the taxonomic utility of this character.
D. wilsoni possesses 15 maxillary alveoli, as in other species of Daspletosaurus ( Carr et al., 2017) . The 13 th alveolus bears a swollen abscess in BDM 107, and the 15 th maxillary tooth conceals a small replacement tooth within its root that is visible in medial (lingual) view. In general, the maxillary teeth are similar to those of other tyrannosaurid species in being labiolingually broad, although not to the degree present in more derived tyrannosaurines ( e.g., Tyrannosaurus and Tarbosaurus ), in which the labiolingual width of the maxillary teeth is subequal to their mesiodistal length ( Carr et al., 2017). The first maxillary alveolus is not small and also bears an incrassate tooth ( i.e., it does not bear a d-shaped crown similar to those present in the premaxillae, as in Gorgosaurus ; Currie, 2003; Voris et al., 2022).
Jugal
The jugal of D. wilsoni is most similar to that of D. torosus among tyrannosaurines in that it has a mediolaterally thin ventral margin of the orbit (as opposed to a rounded margin as in Thanatotheristes , Lythronax , most Tarbosaurus , and some Tyrannosaurus ; Voris et al., 2020; Voris et al., 2022; J. T. Voris, 2022, personal communication). A thin ventral margin of the orbit likely represents the ancestral tyrannosaurid condition, (as it is also present in Bistahieversor , Albertosaurus , Gorgosaurus , and D. horneri ; J. T. Voris, 2022, personal communication) and does not bow medially along its rostrocaudal length (the jugals of D. horneri , Tyrannosaurus , and Tarbosaurus are angled rostromedially rostral to the orbit, such that the maxillae are medially inset from the orbitotemporal region; D. horneri MOR 590, E. A. Warshaw, 2022, personal observations; Tyrannosaurus AMNH 5027, Molnar, 1991, Fig. 9 View Figure 9 ; Tarbosaurus GIN 107/1, Hurum & Sabath, 2003, Fig. 15; E. A. Warshaw, 2022, unpublished data).
As in D. torosus , the caudal portion of the lacrimal contact surface of the jugal is shallowly inclined ( Fig. 4 View Figure 4 ); this surface is very steep in D. horneri , as well as in Albertosaurus and Gorgosaurus ( Carr et al., 2017) . Although Carr et al. (2017) recovered this feature as unique to D. horneri among tyrannosaurines, it is also present in some Tyrannosaurus individuals (MOR 980, MOR 1125, AMNH 5027, E. A. Warshaw, 2022, personal observations).
Lacrimal
As in all tyrannosaurids except for D. horneri , Tarbosaurus , and Tyrannosaurus , the cornual process of the lacrimal in D. wilsoni is large and rises to a distinct apex along its dorsal margin ( Carr et al., 2017; Fig. 5 View Figure 5 ). This apex is situated directly dorsal to the lacrimal’ s ventral ramus, as is characteristic of mature tyrannosaurines ( Currie, 2003; Carr, 2020). The cornual process of the lacrimal is shorter in D. wilsoni (5.2 cm from the dorsal margin of the lacrimal antorbital recess to the apex of the cornual process in BDM 107) than D. torosus (6.9 cm, CMN 8506; Voris et al., 2019, Fig. 6 View Figure 6 ), but similar to the Dinosaur Park taxon (5.1 cm, TMP 2001.36.1; Voris et al., 2019, Fig. 6 View Figure 6 ) (these three specimens are each within 2 cm of each other in skull length, such that measurements of this process need not be corrected for differences in absolute specimen size; see above; Voris et al., 2019, Fig. 6 View Figure 6 ). The lacrimal cornual process of the D. horneri holotype MOR 590 is shorter still (3.7 cm; Carr et al., 2017, Fig. 1 View Figure 1 ), although it should be noted that this specimen is also ~15% shorter in skull length than any of the specimens previously mentioned ( Carr et al., 2017; Voris et al., 2019; see above), such that the difference in this feature between D. horneri and other Daspletosaurus is relatively less pronounced than isolated measurements of this process would suggest (scaled isometrically to the same skull length as MOR 590, however, BDM 107 would still have a taller cornual process of the lacrimal, at 4.4 cm).
Carr et al. (2017) regarded an accessory cornual process of the lacrimal as a synapomorphy of Daspletosaurus . However, this process is indistinguishable from the caudally directed supraorbital process of the lacrimal upon which it is purported to sit; the supraorbital processes of the lacrimals of Tyrannosaurus (MOR 555, MOR 980, MOR 1125, AMNH 5027, E. A. Warshaw, 2022, personal observations), Tarbosaurus (ZPAL MgD-I/4, Hurum & Sabath, 2003, Fig. 6 View Figure 6 ), and Teratophoneus (UMNH VP 16690, Loewen et al., 2013, Fig. 3 View Figure 3 ) are all morphologically identical to those of Daspletosaurus , although they are scored by Carr et al. (2017) as lacking an accessory cornual process. In lieu of any quantitative demonstration of this process’ s presence in Daspletosaurus , the taxonomic utility of this character is rejected here.
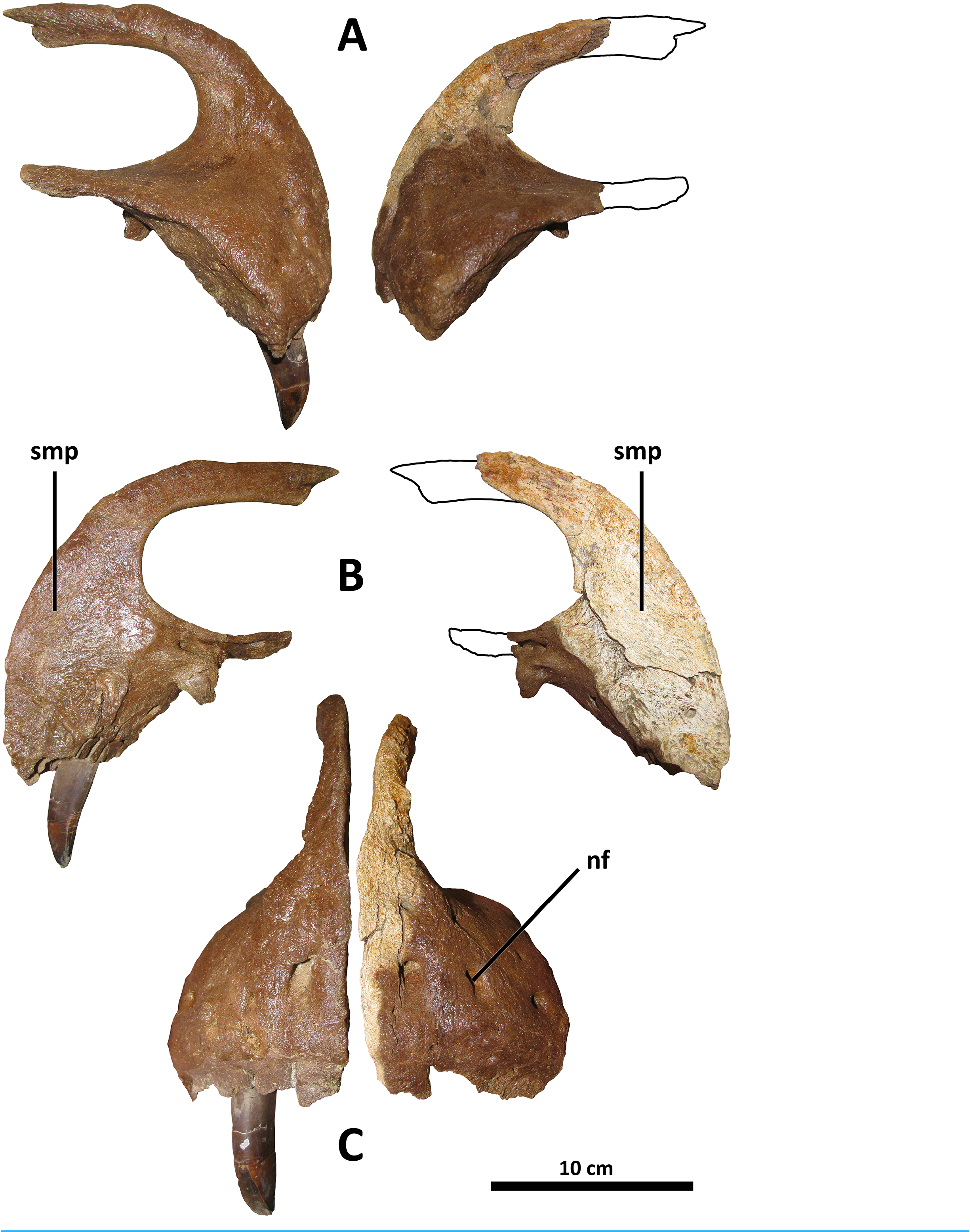
The lacrimal antorbital recess differs in morphology from D. torosus , but is similar to that of D. horneri , Tarbosaurus , and Tyrannosaurus in that the rostrodorsal ala joining the rostral and ventral rami of the lacrimal is inflated into a cylindrical bar that is elevated in relief relative to the rest of the recess (this ala is inflated in D. torosus , but to a lesser degree such that no discrete bar is formed between the rostral and ventral rami; Carr et al., 2017, Fig. S2F View Figure 2 ) (E. A. Warshaw, 2022, personal observations). This feature is also present in the Dinosaur Park taxon (TMP 2001.36.1, Voris et al., 2019, Fig. 6 View Figure 6 ). Also distinguishing the lacrimal of D. wilsoni from D. torosus is a ventrally directed antorbital fossa in the latter. The lacrimal antorbital fossa is laterally directed in other tyrannosaurids, including D. wilsoni , the Dinosaur Park taxon (TMP 2001.36.1, Voris et al., 2019, Fig. 6 View Figure 6 ), D. horneri (MOR 590 and MOR 1130, Carr et al., 2017, Fig. 3 View Figure 3 ), Tyrannosaurus , Tarbosaurus , Albertosaurus , and Gorgosaurus ( Carr & Williamson, 2004, Fig. 10 View Figure 10 ).
Rostrally, the ventral process of the lacrimal rostral ramus is unique in D. wilsoni in having a rounded distal end; this process comes to a pronounced tip in most tyrannosaurids ( Carr, Williamson & Schwimmer, 2005, Fig. 8 View Figure 8 ; Loewen et al., 2013, Fig. 3 View Figure 3 ), with the possible exception of D. horneri , in which the holotype specimen MOR 590 has a pointed ventral process and that of the paratype MOR 1130 is rounded (E. A. Warshaw, 2022, personal observations; Carr et al., 2017, Figs. 2C View Figure 2 and 3 View Figure 3 ). Given the eminent possibility of taphonomic alteration of this feature ( i.e., “rounding down” of a pointed ventral process into a rounded one by abrasion prior to burial), exaggerated by the small size of the ventral process of the lacrimal, this feature is excluded from consideration either as an autapomorphy of D. wilsoni or as uniting this species with D. horneri .
Caudodorsally, the prefrontal articular surface of the lacrimal can be used to determine the orientation of the long axis of the prefrontal. In D. wilsoni and D. torosus , this element is oriented rostrocaudally ( Carr & Williamson, 2004, Fig. 8 View Figure 8 ). This condition is shared with the Dinosaur Park taxon (TMP 2001.36.1, Paulina Carabajal et al., 2021, Fig. 2D View Figure 2 ), and is also present in Gorgosaurus (UALVP 10, Voris et al., 2022), Teratophoneus (UMNH VP 16690, Loewen et al., 2013, Fig. 3 View Figure 3 ), and Qianzhousaurus (GM F10004, Foster et al., 2021, Fig. 3 View Figure 3 ). Conversely, the prefrontal is oriented rostromedially or mediolaterally in D. horneri (MOR 590, E. A. Warshaw, 2022, personal observations; Carr et al., 2017, Fig. 1 View Figure 1 ), Tarbosaurus (ZPAL MgD-I/4, Hurum & Sabath, 2003, Fig. 1 View Figure 1 ), and Tyrannosaurus (AMNH 5027, E. A. Warshaw, 2022, personal observations; Carr & Williamson, 2004, Fig. 8 View Figure 8 ), as well as at least one specimen of Albertosaurus (TMP 1981.10.1, Carr & Williamson, 2004, Fig. 8 View Figure 8 ).
Postorbital
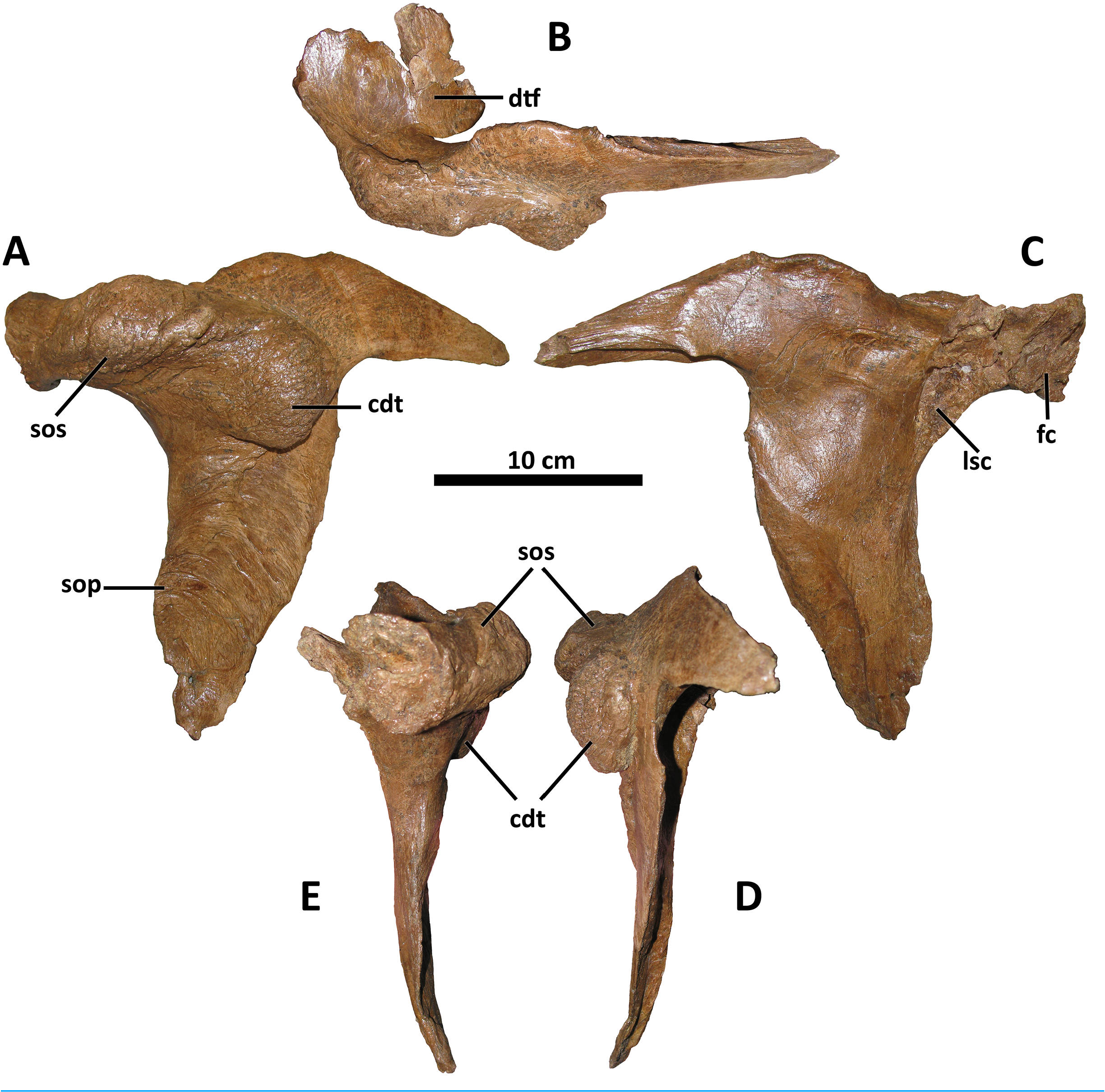
The postorbital of D. wilsoni is most similar to that of D. torosus and the Dinosaur Park taxon in bearing a massive cornual process that approaches the rostral margin of the laterotemporal fenestra caudally ( Fig. 6 View Figure 6 ; Carr et al., 2017; D. torosus CMN 8506, Voris et al., 2019, Fig. 6 View Figure 6 ; TMP 2001.36.1, Voris et al., 2019, Fig. 4 View Figure 4 ). Carr et al. (2017) proposed a cornual process of the postorbital approaching the laterotemporal fenestra as a synapomorphy of Daspletosaurus ; however, the cornual process of the postorbital does not approach the laterotemporal fenestra in the holotype of D. horneri (MOR 590, E. A. Warshaw, 2022, personal observations), and is instead broadly separated from it as in Tyrannosaurus (MOR 980, MOR 1125, MOR 555, E. A. Warshaw, 2022, personal observations) and Tarbosaurus (ZPAL MgD-I/4, Hurum & Sabath, 2003, Fig. 1 View Figure 1 ).
Also shared between D. wilsoni , D. torosus , and the Dinosaur Park taxon is the subdivision of the postorbital cornual process into two discrete processes: a supraorbital shelf protruding from the dorsal margin of the orbit and a caudodorsal tuberosity emerging more caudoventrally ( Fig. 6 View Figure 6 ; Voris et al., 2019, Fig. 4D View Figure 4 ), creating a sinusoidal relief when the postorbital is viewed rostrally or caudally. Both the supraorbital shelf and the caudodorsal tuberosity are situated upon a more ‘typical’ tyrannosaurine cornual process; that is, they lie lateral to a gross swelling of the postorbital similar to that present in other tyrannosaurines ( e.g., Tyrannosaurus, MOR 1125 , MOR 980, MOR 555, MOR 008, E. A. Warshaw, 2022, personal observations). The caudodorsal tuberosity overhangs its caudoventral base, creating a crease between this process and the underlying body of the postorbital; a similar condition is present in the postorbital cornual processes of Gorgosaurus , Teratophoneus , and Bistahieversor ( Voris et al., 2022; J. T. Voris, 2022, personal communication), but not in D. horneri (MOR 590, Carr et al., 2017, Fig. 1 View Figure 1 ), Tyrannosaurus (MOR 1125, MOR 980, MOR 555, MOR 008, E. A. Warshaw, 2022, personal observations), or Tarbosaurus (ZPAL MgD-I/4, Hurum & Sabath, 2003, Fig. 8 View Figure 8 ). A similar crease forms between the body of the postorbital and the cornual process of Tyrannosaurus in specimens with an epipostorbital ( sensu Carr, 2020; AMNH 5027, Molnar, 1991; Carr, 2020); however, no such element is present in the holotype of D. wilsoni (or any other Daspletosaurus specimens; E. A. Warshaw, 2022, personal observations).
The ventral ramus of the postorbital tapers ventrally to a point in D. wilsoni , as in other Daspletosaurus ( Carr, 1999) , including the Dinosaur Park taxon ( Voris et al., 2019, Fig. 4 View Figure 4 ), and in contrast to the enormous subocular process of the postorbital that projects rostrally in Tyrannosaurus ( Carr, 2020) , Tarbosaurus ( Hurum & Sabath, 2003, Fig. 8 View Figure 8 ), Gorgosaurus ( Voris et al., 2022) , Teratophoneus ( Loewen et al., 2013, Fig. 3 View Figure 3 ), and Albertosaurus ( Currie, 2003) . Although the subocular process is present in D. wilsoni (and other Daspletosaurus ), it is small relative to those of other tyrannosaurids ( Fig. 6 View Figure 6 ).
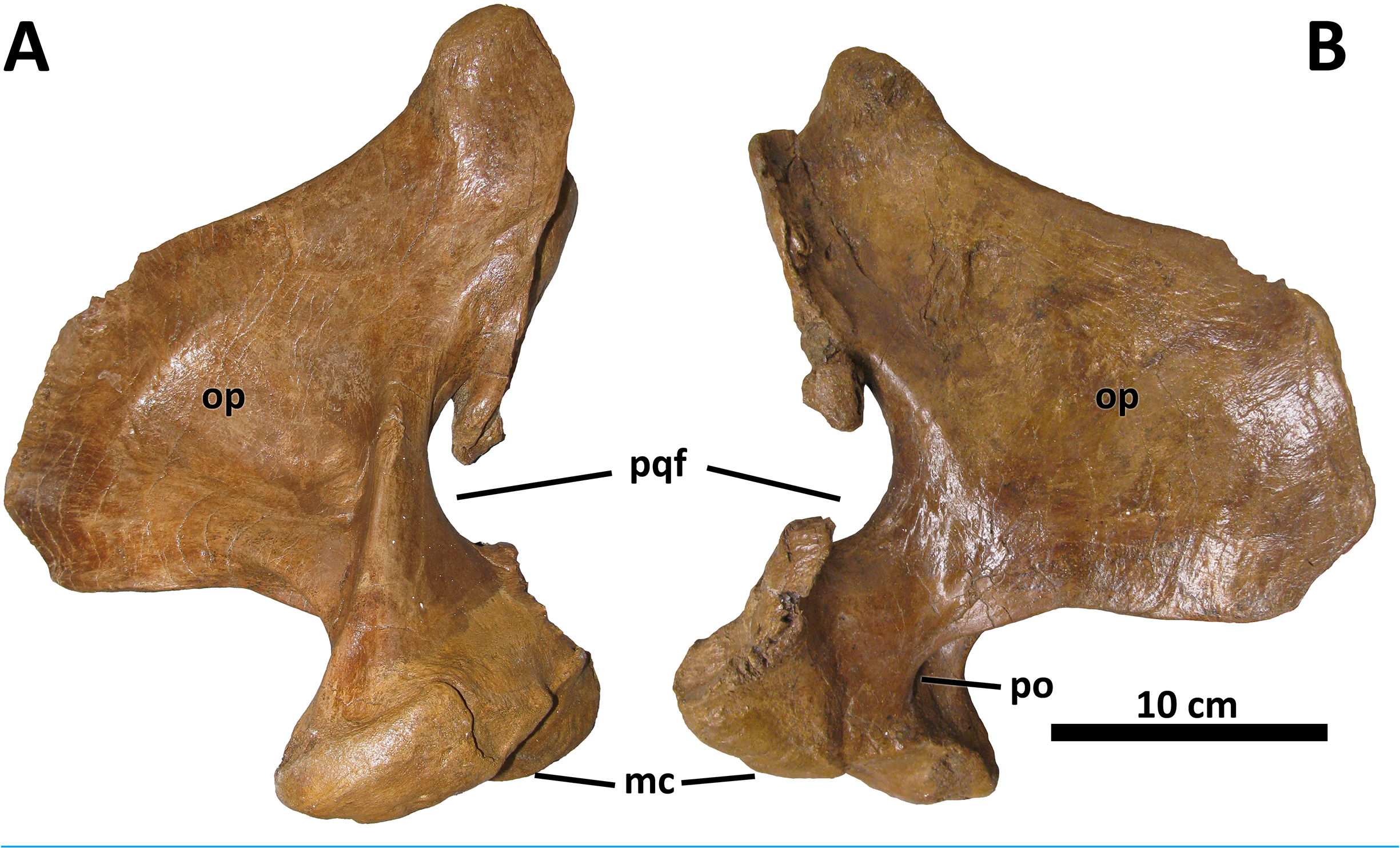
Squamosal
The squamosal of D. wilsoni is indistinguishable from that of D. torosus in that the rostralmost extent of the postorbital contact surface terminates caudal to the rostral margin of the laterotemporal fenestra (also in D. horneri ; Carr et al., 2017), the rostromedial margin of the pneumatic recess on the ventral surface is not undercut ( Fig. 7 View Figure 7 ), and the caudal process is pneumatized (as evidenced by pneumatic foramina in the process’ s rostromedial surface; Carr et al., 2017). No characteristics or combinations of characteristics unique to D. wilsoni are observable on this element.
Quadratojugal
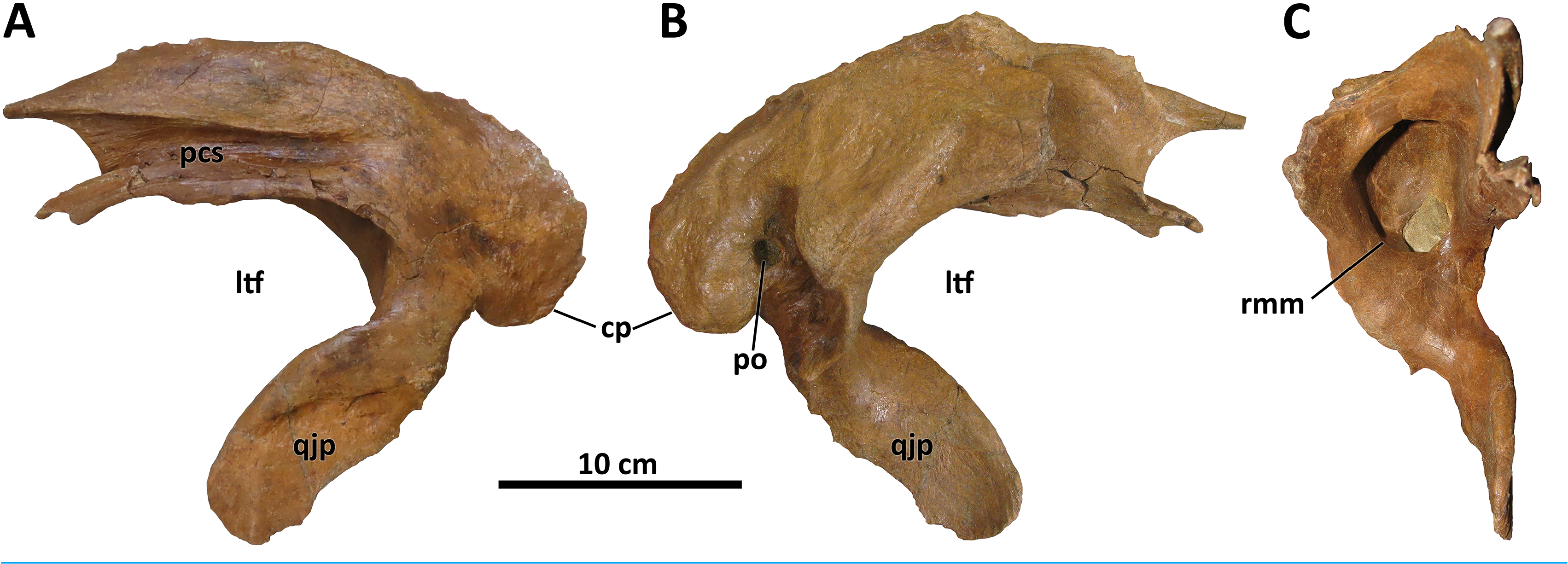
The quadratojugal is conservative morphologically across tyrannosaurids ( Loewen et al., 2013). However, a single characteristic of the quadratojugal of D. wilsoni unites it with D. horneri and at least one specimen of the Dinosaur Park taxon (TMP 2001.36.1), and differs from the condition in D. torosus and less derived tyrannosaurids: a dorsal quadrate contact that is broadly visible in lateral view. In most tyrannosauroids, the dorsal quadrate contact of the quadratojugal is directed medially or rostromedially such that it is obscured by the body of the quadratojugal in lateral view. In D. wilsoni , D. horneri (MOR 590, Carr et al., 2017, Fig. 1 View Figure 1 ), and TMP 2001.36.1 ( Voris et al., 2019, Fig. 6 View Figure 6 ), however, this process is directed caudomedially, exposing it laterally ( Fig. 8 View Figure 8 ).
The dorsal quadrate contact is marginally visible laterally in the holotype of D. torosus, CMN 8506 ( Voris et al., 2019, Fig. 6 View Figure 6 ; J. T. Voris, 2022, personal communication), but not nearly to the extent observable in the aforementioned taxa. The condition in D. torosus may therefore represent individual variation on the caudomedial orientation of most tyrannosaurids, or a structural antecedent to the condition present in other species of Daspletosaurus .
The caudomedial orientation of the dorsal quadrate contact is reversed in the paratype specimen of D. horneri , in which this process is hidden in lateral view (MOR 1130, Carr et al., 2017, Fig. S2K View Figure 2 ). Given that this specimen is younger stratigraphically than the holotype (MOR 590; Carr et al., 2017), this reversal may represent a phylogenetic signal (although it may instead represent intraspecific variation). Tarbosaurus and Tyrannosaurus share this feature with MOR 1130 (E. A. Warshaw, 2022, personal observations).
Quadrate
No discrete morphological characters distinguish the quadrate of D. wilsoni from those of its closest relatives. As in other derived tyrannosaurines, the quadrate is massive, with a shallow fossa on its medial surface and a pronounced pneumatic foramen (and surrounding fossa) at the rostral confluence of the mandibular condyles and the orbital process ( Fig. 9 View Figure 9 ; Carr et al., 2017). The paraquadrate foramen, bounded medially by the quadrate and laterally by the quadratojugal, is small and teardrop-shaped; only its lateral margin is made up by the quadratojugal, as the quadrate forms the dorsal and ventral borders of the foramen.
Although no palatal elements are known, the medial deflection of the quadrate’ s pterygoid wing allows an approximation of the position of the pterygoids relative to the facial skeleton, and suggests a broad orbitotemporal region, as in other tyrannosaurines. Dentary
The dentary of D. wilsoni is deep, with a relatively straight ventral margin and a dorsal (alveolar) margin that trends caudodorsally, increasing the depth of the mandible caudally ( Fig. 10 View Figure 10 ). As in other Daspletosaurus , the texturing of the dentary symphysis is more exaggerated in D. wilsoni than non- Daspletosaurus tyrannosaurines ( e.g., Tyrannosaurus , Thanatotheristes ; Voris et al., 2020), and is composed of several interlocking (presumably, as only the left dentary is known) ridges and cusps. There are 17 dental alveoli, as in D. horneri ( Carr et al., 2017) , and a sharp, narrow Meckelian groove with a rugose knob caudoventral to its rostral end. This knob is present in both other species of Daspletosaurus , as well as Tyrannosaurus , Tarbosaurus , and Zhuchengtyrannus magnus , but not Thanatotheristes or more basal tyrannosaurids ( Carr et al., 2017; Voris et al., 2020).
The lateral surface of the dentary of BDM 107 bears two intersecting grooves caudoventral to the caudal termination of the alveolar margin ( Fig. 10 View Figure 10 ); the edges of these grooves are beveled and are likely pathological. They may represent bite marks, as have been described on the craniofacial bones of other tyrannosaurids ( Voris et al., 2020; Brown, Currie & Therrien, 2022).
Splenial
The splenial of BDM 107 is typical of Daspletosaurus except in the size and form of the mylohyoid foramen ( Fig. 11 View Figure 11 ), an autapomorphy of this taxon. In most derived tyrannosaurines, including D. torosus and D. horneri , this foramen is extremely large, roughly the same dorsoventral depth as the rostral process of the splenial ( Carr et al., 2017). In D. wilsoni , however, the foramen is dorsoventrally shallow, and rostrocaudally elongate, such that it is ellipsoid in form and roughly half the dorsoventral depth of the splenial’ s rostral process. This is most similar to the condition in alioramins ( Brusatte, Carr & Norell, 2012) and Appalachiosaurus ( Carr, Williamson & Schwimmer, 2005) .
Cervical vertebrae
Four cervical vertebrae are preserved in BDM 107 from the cranial-middle portion of the series. No atlas or axis were found. As in all tyrannosaurids, the spinous processes of the cervical vertebrae are subequal in dorsoventral height to their corresponding centra. Both the spinous processes and the centra are craniocaudally short, similar to and most exaggerated in the cervical vertebrae of Tyrannosaurus (see Brochu, 2003, and figures therein). As in Tyrannosaurus (and other large tyrannosaurids), the cranial and caudal faces of the cervical centra in BDM 107 are dorsoventrally displaced from one another in order to create the characteristic ‘S-curve’ of the neck, and the cranial cervical centra are extremely foreshortened craniocaudally ( i.e., much taller than long). This indicates a robustly built cranial portion of the neck, presumably in order to support the weight of the head.
Sacral vertebrae
The spinous processes of two sacral vertebrae are preserved. Both are sub-rectangular in form and bear rugose knobs near their apices, presumably the ossified bases of sacral ligaments.






No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |