
Micronycteris brosseti, Simmons & Voss, 1998
|
publication ID |
https://doi.org/ 10.5281/zenodo.4545052 |
|
DOI |
https://doi.org/10.5281/zenodo.4546461 |
|
persistent identifier |
https://treatment.plazi.org/id/4F19FC10-FFAD-FFE5-FD23-27BFFB978886 |
|
treatment provided by |
Plazi |
|
scientific name |
Micronycteris brosseti |
| status |
sp. nov. |
Micronycteris brosseti View in CoL , new species
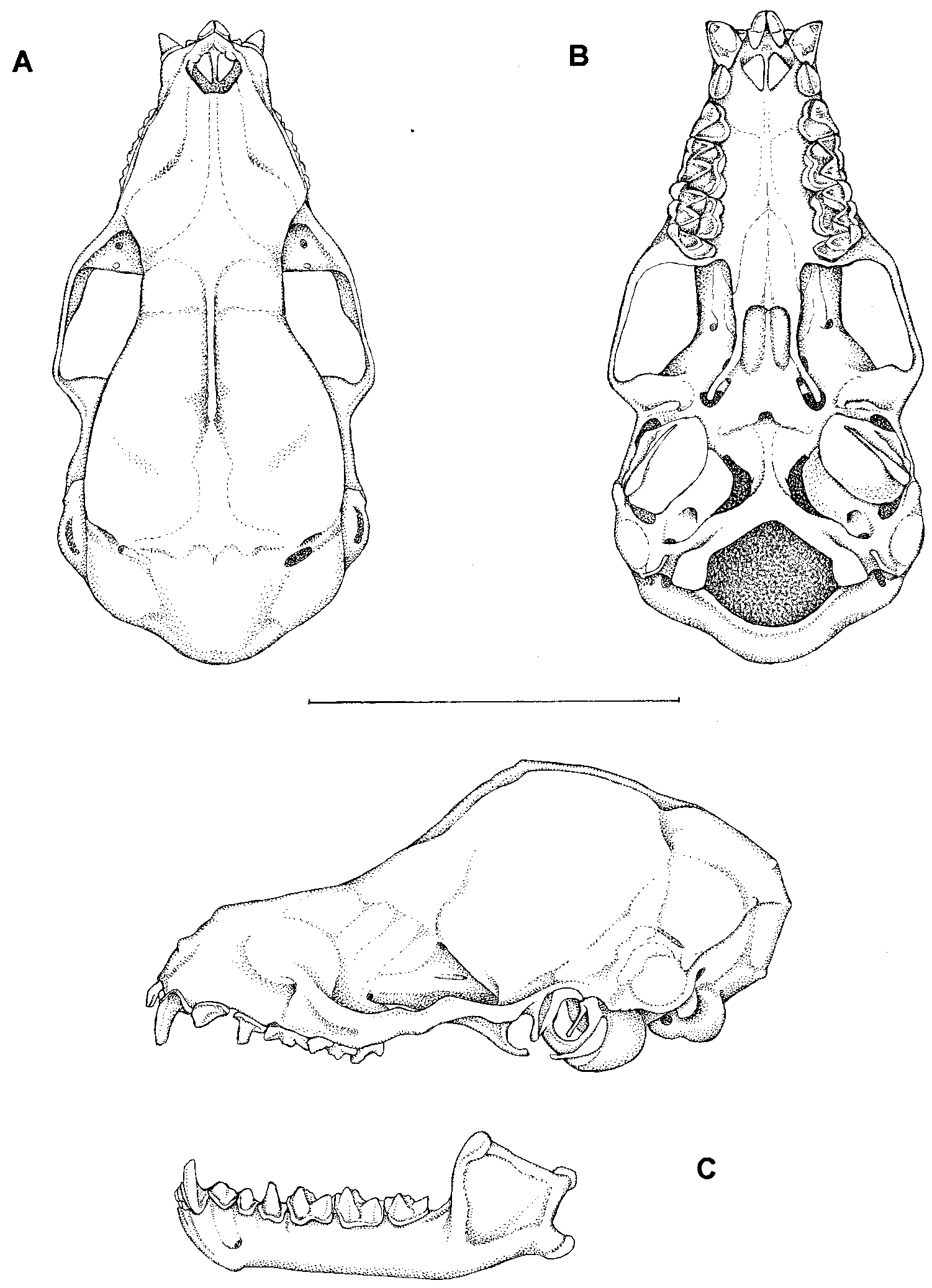
Figure 28 View Fig
TYPE MATERIAL: The holotype ( MNHN 1995.1030 ), an adult male preserved in alcohol with the skull removed and cleaned, was collected at Paracou by N. B. Simmons and R. S. Voss (original number: NBS 372 ) on 20 August 1991. Paratypes include 3 females ( AMNH *266033 , *266038 ; MNHN *1995.1031 ) and 4 males ( AMNH *266032 , *266034 , *266035 , *267419 ), all of which were likewise collected at Paracou . See tables 22 and 23 for measurements.
REFERRED MATERIAL: In addition to the type series from Paracou, we also refer nine specimens to Micronycteris brosseti from four other South American localities as follows: PERU, Loreto, Puerto Indiana, Río Amazonas near mouth of Río Napo (AMNH 73495, 73496, 73498, 73499); Peru, Loreto, Jenaro Herrera (MUSM 5528); Peru, Loreto, Río Yavarı´ Mirı´m, Quebrada Esperanza (FMNH 89100, 89101, 89102); BRAZIL, São Paulo, Rio Juquia´, Barra (FMNH 92997). One of these specimens (MUSM 5528) was previously identified as M. schmidtorum by Ascorra et al. (1991a).
DISTRIBUTION: Although specimens of Micronycteris brosseti are currently known only from eastern Peru, French Guiana, and southeastern Brazil, the species could reasonably be expected to occur throughout Amazonia and the Brazilian Atlantic forest.
ETYMOLOGY: We name this species to hon or André Brosset in recognition of his many important contributions to knowledge of tropical bat faunas in India, Africa, and South America. In particular, Brosset and CharlesDominique (1990) provided an indispensable guide to the bat fauna of French Guiana, as the tattered covers, creased pages, and annotated margins of our wellthumbed field copy eloquently attest.
DIAGNOSIS: A small Micronycteris with dark brown dorsal fur and pale (gray or buff) ventral fur; dorsal hairs bicolored, 7–8 mm long in shoulder region, with white base comprising ⅓–½ of each hair; fur on outside of medial third of pinna short (≤ 3 mm) and dense; pinnae large with rounded tips, connected across crown of head by low interauricular band with shallow midline notch; thumb small; second phalanx of wing digit IV shorter than first; calcar longer than foot; mastoid breadth less than zygomatic breadth; crown height of upper incisors not reduced; no gap present between outer upper incisor and canine; P3 and P4 subequal in anteroposterior length, height of P3 slighly less than that of P4; P4 with small lingual heel and poorly developed lingual cusp; M1 narrower than M2; no noticeable gap between posterior edge of cingulum of M2 and anterolingual edge of M3 when toothrow seen in occlusal view; lower incisors small and bilobed; p2 and p4 subequal in anteroposterior length and height, p3 smaller in both dimensions; coronoid process high, upper margin of ascending process with steep slope (25–308).
Of the characters listed above, four are particularly useful for field identification of M. brosseti : color of the ventral fur (pale gray or buff), length of the fur on the leading edge of the pinna (short, ≤ 3 mm), relative length of the first and second phalanges of wing digit IV (second shorter than first), and the calcar (longer than foot). Within the genus Micronycteris , this combination of traits is unique to M. brosseti .
MEASUREMENTS: A summary of measurements of Micronycteris brosseti , including those of the type series and of all other referred specimens, is provided in table 23.
DESCRIPTION AND COMPARISONS: Micronycteris brosseti requires close comparison with six other likesize congeners with which it might be confused, namely M. homezi , M. megalotis , M. microtis , M. minuta , M. sanborni , and M. schmidtorum . By contrast, M. hirsuta stands out from all other members of the genus by its larger size and unique lower incisor morphology (see table 24 and species account for hirsuta below). The following paragraphs therefore omit hirsuta from further consideration and focus only on the remaining seven species.
Micronycteris brosseti is one of the smallest members of the genus, with mean values for most of its measurements falling well below those of other species found at Paracou (tables 23–25). Despite some morphometric overlap between larger individuals of brosseti and a few smaller specimens in the congeneric series measured by Simmons (1996b: table 2), measurements are clearly useful for distinguishing brosseti from most other Micronycteris species where they occur sympatrically. At Paracou, the only species from which brosseti is indistinguishable in univariate morphometric comparisons (tables 23–25) is microtis .
At Paracou, brosseti is consistently small er than megalotis in body weight (females only), ear length, thumb length, greatest length of skull (males only), braincase breadth (females only), mastoid breadth, zygomatic breadth (females only), maxillary toothrow length, and breadth across molars (males only). Similarly, brosseti is smaller than homezi (known only from one male; see species account below) in body weight, ear length, forearm length, tibia length, and in all craniodental measurements. In comparison with minuta , brosseti is consistently smaller in body weight, ear length, greatest length of skull (females only), condyloincisive length (females only), postorbital breadth, braincase breadth (females only), mastoid breadth, maxillary toothrow length (females only), and breadth across molars (females only). Likewise, brosseti is smaller than schmidtorum in many measurements, including body weight, ear length, tibia length, thumb length, and all craniodental measurements.
The entire dorsal pelage of brosseti is composed of bicolored hairs with white bases and brown tips. In the upper back region, the white bases comprises approximately onethird to onehalf of the length of each hair. This is comparable to the pattern seen in microtis , megalotis , schmidtorum , and homezi , in which the white bases comprise onefourth to onehalf of each hair on the upper back. In contrast, the white bases are much more extensive in minuta (onehalf to twothirds of each hair) and sanborni (twothirds to threefourths of each hair).
The length of the dorsal fur over the upper back in brosseti is approximately 7–8 mm, comparable to that of microtis and homezi . In contrast, minuta and sanborni have shorter fur over the shoulders (5–7 mm), while megalotis and schmidtorum have longer fur (8–11 mm).
The ventral fur of brosseti (including that covering the neck and throat) is either pale gray or pale buff, whereas that of megalotis and microtis is brown (approximately the same shade as the dorsal fur). The underparts of schmidtorum and minuta are also pale (gray or buff), like brosseti , but sanborni has bright white ventral fur.
All of the Paracou specimens of brosseti have a pale gray venter, as do some specimens from Peru (e.g., MUSM 5528). However, some older and somewhat faded Peruvian specimens of brosseti have pale buff venters (e.g., AMNH 73495–73499, collect ed in 1926). It is not clear if the buff color is the result of postmortem alteration or withinspecies variation. Both ventral furcolor variants can be observed among museum series of other species (e.g., minuta and schmidtorum ), so this variation (whatever its cause) is evidently not peculiar to brosseti .
The fur on the outside of the medial third of the ear pinna is short (≤ 3 mm) and dense in brosseti , similar to the condition seen in microtis , minuta , and sanborni . By contrast, the auricular fur of megalotis , schmidtorum , and homezi is much longer (5–8 mm).
As in all other species of Micronycteris , the ears of brosseti are connected across the crown of the head by an interauricular band of skin. In brosseti , this band is low and nearly horizontal in profile, and is marked at the midline by a shallow notch; a similar condition is seen in megalotis and microtis . This morphology contrasts sharply with the interauricular band in minuta , sanborni , and homezi , which is much higher and divided by a deep median notch into two roughly triangular flaps (see species account for homezi below). The interauricular band of schimidtorum is of intermediate height with a moderately deep median notch.
The second phalanx of wing digit IV is shorter than the first in brosseti , as it is also in megalotis , microtis , schmidtorum , and homezi . By contrast, the first and second phalanges of digit IV are subequal in minuta and sanborni .
The calcar is distinctly longer than the hindfoot in brosseti , resembling the condition seen in megalotis , microtis , and schmidtorum . The calcar is approximately equal in length to the hindfoot in sanborni , however, and the calcar is shorter than the foot in minuta and homezi .
As noted by Simmons (1996b), cranial morphology is remarkably similar in all species of Micronycteris . Like other members of the genus, brosseti has a relatively long rostrum, no anteorbital inflation, a large braincase, and shallow basisphenoid pits. Mastoid breadth is less than zygomatic breadth in brosseti , as it is likewise in megalotis , microtis , and schmidtorum ; mastoid breadth is greater than zygomatic breadth in minuta , sanborni , and homezi .
Like all other congeneric species, brosseti has a dental formula of I 2/2, C 1/1, P 2/3, M 3/3 x 2 = 34. The crown height of the upper incisors is not reduced, and no gap is present between the outer upper incisor and the canine. In this respect, brosseti resembles most species of Micronycteris ; the contrasting morphology is seen only in sanborni .
Although differences in the upper premolar dentition are subtle, variation in this region provides an important means of distinguishing species of Micronycteris (see Simmons, 1996b). In brosseti , P3 and P4 are subequal in anteroposterior length, and the height of P3 is very slighly less than that of P4. This is similar to the condition seen in megalotis , microtis , schmidtorum , and homezi . Some variation is known in the former three species, with P3 and P4 subequal in height in some individuals and P3 slightly smaller than P4 in others. Although we did not observe such variation in brosseti (or homezi , known from only one specimen), it would not be unexpected. In contrast to the conditions described above, P3 is noticeably smaller than P4 in both length and height in minuta and sanborni .
The degree of development of the posterolingual heel and lingual cusp on P4 varies among and within species of Micronycteris . In brosseti , P4 has a small lingual heel and a poorly developed lingual cusp. The lingual heel of P4 in brosseti is similar in size to those of megalotis , microtis , schmidtorum , and homezi , but the lingual heel is generally smaller in minuta and larger in sanborni . The lingual cusp of P4, which is formed from the raised edge of the tooth, is poorly developed and lacks a sharp point in brosseti , homezi , minuta , and sanborni , and it is entirely lacking in many specimens of schmidtorum . By contrast, this cusp is better developed and tapers to a sharp point in most specimens of microtis and megalotis ; however, some individuals of both species have a poorly developed cusp, while this cusp may be lacking entirely in others.
The first upper molar is noticeably narrow er than M2 in brosseti as it is in most other species of Micronycteris . Uniquely, the lingual portion of M1 is expanded so that M1 and M2 are subequal in width in sanborni , which also differs from all other species in having a large gap visible between the posterior edge of the cingulum of M2 and the anterolingual edge of M3 when the toothrow is seen in occlusal view.
The lower dentition of brosseti closely resembles that of most other species, with minor differences apparent only in the premolars. In brosseti and schmidtorum , p2 and p4 are subequal in anteroposterior length and in height, while p3 is slightly smaller in both dimensions. In minuta , sanborni , and homezi , p3 is further reduced relative to p2 and p4, which are large and subequal. By contrast, p3 is a larger tooth (approximately the same size as p4) in microtis and megalotis ; in these taxa, p2 is often slightly larger than either p3 or p4, although there is some withinspecies variation in this trait and all three lower premolars are subequal in some individuals of both species.
The coronoid process in brosseti is high, and the upper margin of the ascending process has a steep slope (25–308), similar to the mandibular morphology of megalotis , microtis , and schmidtorum . In contrast, the coronoid process is comparatively low and the upper margin of the ascending ramus has a shallow slope (16–188) in minuta , sanborni , and homezi .
FIELD OBSERVATIONS: Of the eight specimens of Micronycteris brosseti that we collected at Paracou, one was taken in a groundlevel mistnet and the remaining seven from a single roost in a hollow tree; both capture sites were in welldrained primary forest. The roost tree, about 1 m in diameter at breast height, had a single entrance hole (about 20 cm in diameter) 1 m above the ground. A mistnet enclosure around the base of the tree captured seven of the eight bats that emerged (between 18:45 and 19:15 hours) on 19 August 1991; the captured animals consisted of four adult males and three adult females. In addition, the mistnet enclosure captured two Phyllostomus elongatus on the outside as they came swooping toward the roost opening, obviously intending to ambush the much smaller emerging bats. We were unable to revisit this roost in subsequent years because the site was cleared for agricultural research in 1992.
| MNHN |
France, Paris, Museum National d'Histoire Naturelle |
| AMNH |
USA, New York, New York, American Museum of Natural History |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Phyllostominae |
|
Genus |