
Proctogastrolaelaps subsolanus Joharchi & Marchenko, 2021
|
publication ID |
https://doi.org/ 10.11646/zootaxa.5072.4.5 |
|
publication LSID |
lsid:zoobank.org:pub:14DA1F98-A010-46FC-A657-E1EEDF2069DE |
|
DOI |
https://doi.org/10.5281/zenodo.5748834 |
|
persistent identifier |
https://treatment.plazi.org/id/4F1A3318-FF99-933B-FF3F-6EC5FD3DFC5E |
|
treatment provided by |
Plazi |
|
scientific name |
Proctogastrolaelaps subsolanus Joharchi & Marchenko |
| status |
sp. nov. |
Proctogastrolaelaps subsolanus Joharchi & Marchenko sp. nov.
( Figures 1–18 View FIGURES 1–5 View FIGURES 6–14 View FIGURES 15–18 )
Type material: Holotype, female, Tunaycha Lake , Sakhalin Island, Russia, 46°51’22.4”N 143°07’01.4”E, 10 August 2021, A.A. Khaustov, O. Joharchi, I. Döker and V. A. Khaustov coll., from soil-litter on the shore of the Tunaycha Lake (in TSUMZ) GoogleMaps . Paratypes, four females same data as holotype and GoogleMaps five females, Sakhalin Island , Russia, 47°26’22.4”N 142°45’42.0”E, 12 August 2021, A.A. Khaustov, O. Joharchi, I. Döker and V. A. Khaustov coll. GoogleMaps , from soil-litter on the shore of the Sea of Okhotsk (in TUMZ & ISEA) .
Additional material: one female, Sakhalin Island, environs of Tunaicha Lake , mixed forest, in litter, 8 July 1991, I.I. Volonikhina (= Marchenko) ; one female, Sakhalin Island, Korsakov District, environs of Lesnoe village , mixed forest, in litter, 18 October 1993, A. Basarukin coll. (in ISEA) .
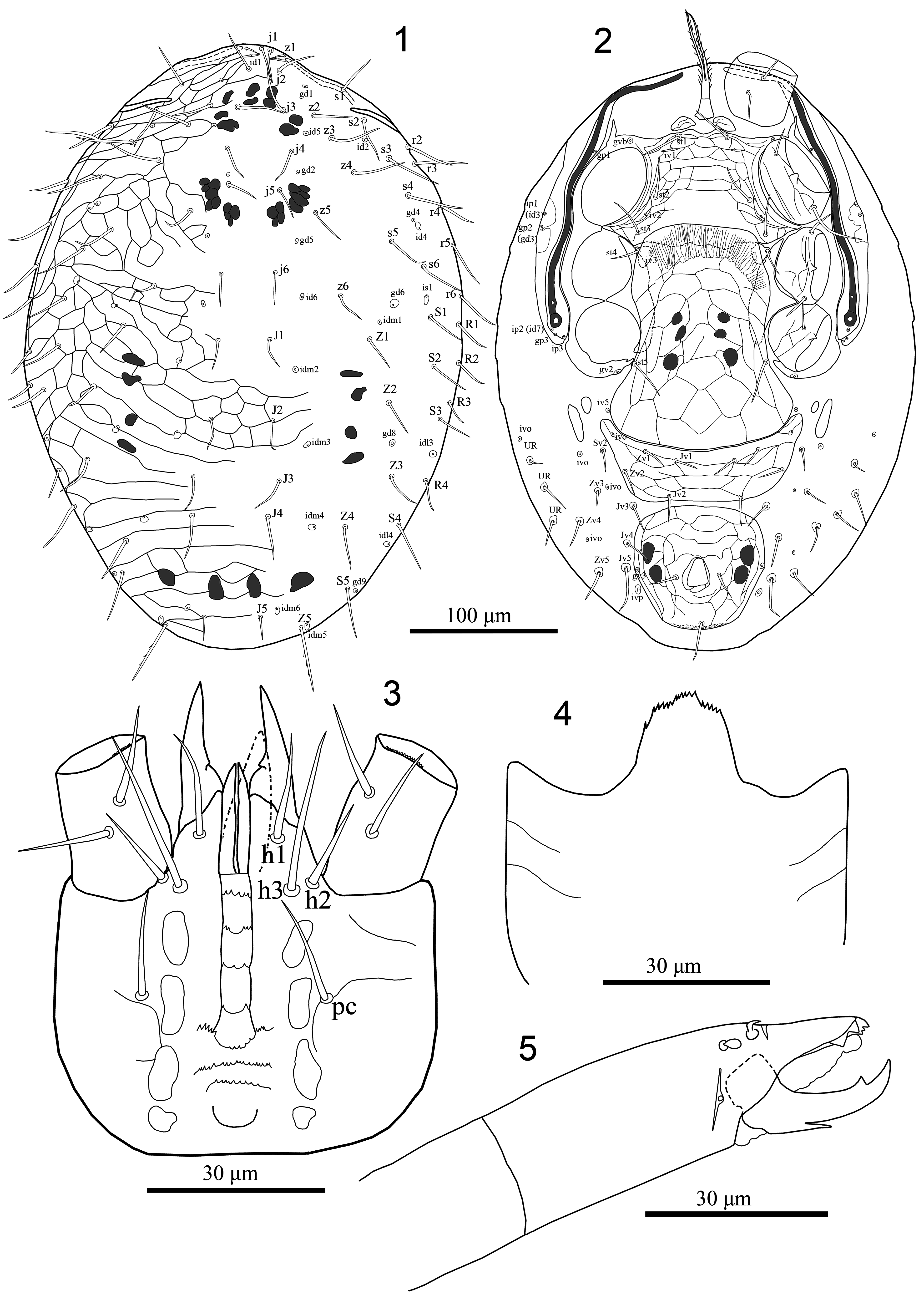
Diagnosis (adult female). Dorsal shield with distinct reticulate ornamentation over whole surface, more distinct in the lateral and opisthonotal sections, with delineated strip along lateral margins and 42 pairs of setae (23 pairs of podonotal setae and 19 pairs of opisthonotal setae, including r6 and R1–4); pre-sternal area comprises with two spindle shaped platelets; sternal shield completely reticulated except posterior area where overlapped by hyaline flap of genital shield; genital shield axe-shaped, expansive, broadened posteriorly, angled posterolaterally behind coxae IV and posteriorly slightly convex; opisthogaster with 14 pairs of setae (including three pairs of UR setae) mostly inserted on small sclerotised rings around basal sockets; anal shield with para-anal setae inserted mid-length of anal opening, post-anal seta longer than para-anals; anal opening more or less enlarged, about 1/4 of shield length, in shield center; peritrematal shield well developed, free from exopodal shield beside coxa IV; epistome with about 20 closely approximated middle-sized denticles; anterior four rows of hypostomal groove each bearing two lateral and 1–3 medial denticles, 5 th row widened, with 13–15 denticles, 6 th and 7 th rows multidentate; corniculi with paraxial process near base, some leg setae thickened and inserted on small tubercles.
Description. Female (n=10) ( Figures 1–18 View FIGURES 1–5 View FIGURES 6–14 View FIGURES 15–18 )
Dorsal idiosoma. Dorsal shield 400–410 long, 222–246 wide, covering most of dorsal idiosoma; shield with distinct reticulate ornamentation over whole surface, more distinct in the lateral and opisthonotal sections, with delineated strip along lateral margins ( Figs 1 View FIGURES 1–5 , 6 View FIGURES 6–14 ). Shield with 42 pairs of setae [23 pairs of podonotal setae (j1–j6, z1–z6, s1–s6, r2–r6) and 19 pairs of opisthonotal setae (J1–J5, Z1–Z5, S1–S5, R1–R4 ))], most dorsal setae similar lengths (21–41), at most as long as distance to base of subsequent setae, central setae shorter, all setae acicular and smooth, excepting shorter z1 (10–12), J5 (16–19); Z5 (45–48) longer than other setae and sparsely barbed ( Figs 1 View FIGURES 1–5 , 6 View FIGURES 6–14 ). Shield with about 21 pairs of discernible pore-like structures, including 14 poroids (id1, id2, id4–id6, idm1–idm6, is1, idl3, idl4) and seven gland openings (gd1, gd2, gd4, gd5, gd6, gd8, gd9), others indistinct ( Figs 1 View FIGURES 1–5 , 6 View FIGURES 6–14 ).
Ventral idiosoma ( Figures 2 View FIGURES 1–5 , 7–10, 12 View FIGURES 6–14 ). Tritosternum with paired pilose laciniae (50–54), fused basally (14–17), columnar base 13–15 × 12–14 wide; pre-sternal area comprises two spindle shaped platelets. Sternal shield length 80–85, maximum width 133–140, narrowest between coxae II (73–78), with distinct reticulate ornamentation over whole surface, except posterior area between setae st2–st3, where overlapped by hyaline flap of genital shield smooth or faintly reticulated ( Figs 2 View FIGURES 1–5 , 7, 9 View FIGURES 6–14 ), posterior margin almost straight but anterior margin undulating, bearing three pairs of smooth setae (st1 22–25, st2 26–29, st3 26–29), st2 closer to st3 than to st1; reaching base of it, poroids iv1–2 slit-like, iv1 aligned transversely adjacent to setae st1, iv2 oblique between st2 and st3. Metasternal setae st4 (19–21) and metasternal poroids located on small metasternal platelets, mostly overlapped by anterior margin of genital shield ( Figs 2 View FIGURES 1–5 , 7, 9 View FIGURES 6–14 ). Endopodal element between coxae I–II fused to sternal shield, bearing gland pore gvb, and fused anteriorly to exopodal strip that flanks coxae I–IV; endopodal platelets between coxae III–IV well-developed, partly overlapped by anterior margin of genital shield, obviously free from sternal shield ( Figs 2 View FIGURES 1–5 , 7 View FIGURES 6–14 ). Genital shield somewhat axe-shaped, length 147–165, width at level of setae st5 and broadest level 92–95 and 124–128, respectively, overlapping posterior margin of sternal shield, extending to level of setae st3, expansive, broadened posteriorly, angled posterolaterally behind coxae IV, posterior margin slightly convex, surface well reticulated with irregular cells; genital setae st5 (28–31) and poroids iv5 inserted on soft cuticle laterad genital shield at posterior level of parapodal platelets behind coxa IV; postgenital platelets absent ( Figs 2 View FIGURES 1–5 , 7, 8 View FIGURES 6–14 ). Ventral shield somewhat trapezoid, shield (31–35) long, with greatest width in anterior corners, at level of poroids (ivo) (138–147), width at mid-level of shield (setae Zv2) (115–121) and at the posterior margin, at level of setae Jv2 (98–105); bearing four pairs of smooth setae (16–20) (Jv1–2, Zv1–2) and a pair of poroid (ivo) at anterior corner of shield, surface well reticulated with irregular cells ( Figs 2 View FIGURES 1–5 , 7, 8 View FIGURES 6–14 ). Anal shield relatively rounded, slightly wider (88–92) than long (81–85), almost whole surface lineate-reticulate; Anal opening somewhat enlarged, located mainly in center region of shield, anal opening 19–22 × 13–15 wide; post-anal seta (25–29) longer than para-anal setae (14–17), cribrum wide, consisting of a terminal tuft with 3–4 irregular rows of spicules, reaching posterolateral corners of shield, limited to region posterior to post-anal seta, gland pores gv3 on lateral margins of shield slightly anteriad para-anal setae level ( Figs 2 View FIGURES 1–5 , 7, 10 View FIGURES 6–14 ). Soft opisthogastric cuticle with two pairs of metapodal plates, the anterior small, ovoid (8–12 long × 4–6 wide) and the posterior larger, fusiform (27–30 long × 6–8 wide), ten pairs of smooth and same length setae (14–22) (JV3–JV5, ZV3–ZV5, SV2, three setae of UR -series), except Jv5 (30–33) longest, mostly inserted on small sclerotised rings around basal sockets and five pairs of poroids (iv5, three pairs of ivo; ivp) ( Figs 2 View FIGURES 1–5 , 7 View FIGURES 6–14 ). Peritrematal shield well developed laterad peritremes, bearing one gland pore and one poroid at level between coxae II–III; poststigmatic extension of shield relatively small, free from exopodal shield beside coxa IV, with two pairs of poroids and one pair of gland pores; anterior region of peritrematal shield fused to anterior margin of dorsal shield anteriad seta s2 ( Figs 2 View FIGURES 1–5 , 7, 12 View FIGURES 6–14 ). Peritremes long, extending anteriorly to anterior level of coxae I (level of seta z1). Parapodal element behind coxae IV relatively thick and bearing gland pore gv2, fused to exopodals flanks coxae I–IV. Spermathecal apparatus not distinguishable.
Gnathosoma ( Figs 3–5 View FIGURES 1–5 , 11, 13–14 View FIGURES 6–14 ). Epistome semi-triangular, with about 20 closely approximated middlesized denticles ( Fig. 4 View FIGURES 1–5 ). Corniculi robust and horn-like, with paraxial process near base, smooth internal malae and acuminate labrum obviously shorter than corniculi; salivary stylets narrow, apically blunt, longer than corniculi; hypostomal and capitular setae smooth, h3 (27–29)> pc (16–19)> h1 (14–17)> h2 (13–15); hypostomal groove with seven rows of denticles, rows 1–5 connected by lateral lines, anterior four rows each bearing two lateral and 1–3 medial denticles, 5 th row widened, with 13–15 denticles, 6 th and 7 th rows free, multidentate ( Figs 3 View FIGURES 1–5 , 11 View FIGURES 6–14 ). Supralabral process not distinguishable. Chaetotaxy of palps: 2–5–6–14–15; palpfemoral seta al and palpgenual setae al2 bacilliform flattened apically and spatulate, respectively, other setae simple, palptarsal claw two-tined ( Fig. 13 View FIGURES 6–14 ). Second cheliceral segment length (69–73), movable digit length (30–33) and basal segment length (42– 45). Cheliceral dorsal and antiaxial lyrifissures distinct; hyaline rim on paraxial surface of fixed digit absent. Fixed digit of chelicera with short and prostrate dorsal mucro near short and subspatulate dorsal seta, five teeth of various sizes (including two large subterminal teeth not aligned with others), and expanded membranous lobe instead of setiform pilus dentilis; movable digit with one tooth in addition to apical tooth and sharp proximal mucro ventrally ( Figs 5 View FIGURES 1–5 , 14 View FIGURES 6–14 ).
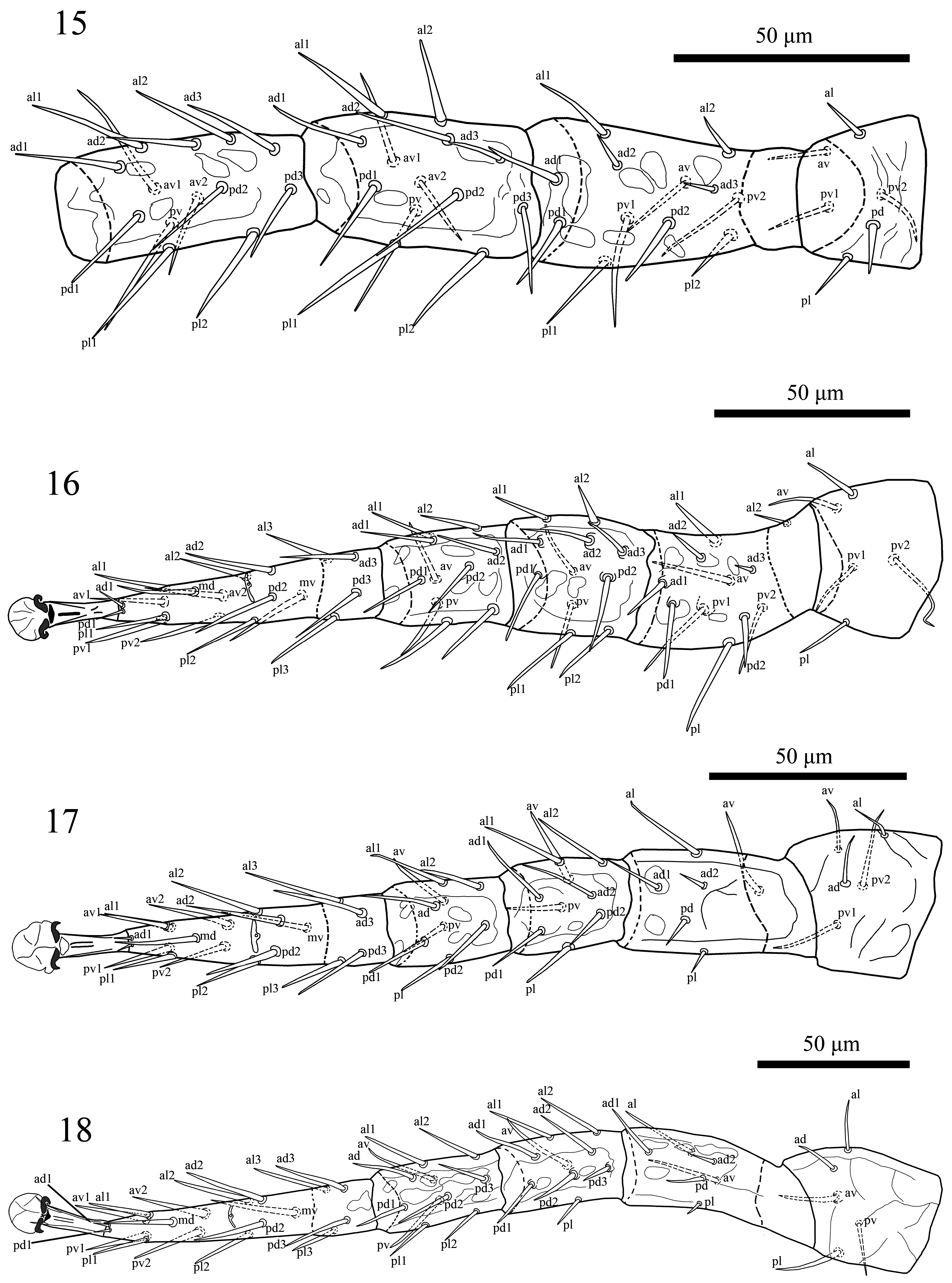
Legs ( Figs 15–18 View FIGURES 15–18 ). Legs II and III (236–247) short, legs I (308–313) and IV (310–318) longer. Chaetotaxy: Leg I ( Fig. 15 View FIGURES 15–18 ): coxa 0 0/1, 0/1 0, trochanter 1 0/1, 1/2 1 (al and pd thickened), femur 2 3/1, 2/2 2 (ad1, pd1, pd2 thickened), genu 2 3/2, 3/1 2 (al1, al2 thickened), tibia 2 3/2, 3/1 2. Leg II ( Fig. 16 View FIGURES 15–18 ): coxa 0 0/1, 0/1 0, trochanter 1 0/1, 0/2 1 (al slightly thickened), femur 2 3/1, 2/2 1 (ad1, pd1, pv1 inserted on small tubercles), genu 2 3/1, 2/1 2 (all dorsal setae inserted on small tubercles), tibia 2 2/1, 2/1 2. Leg III ( Fig. 17 View FIGURES 15–18 ): coxa 0 0/1, 0/1 0, trochanter 1 1/1, 0/2 0, femur 1 2/1, 1/0 1, genu 2 2/1, 2/1 1, tibia: 2 1/1, 2/1 1. Leg IV ( Fig. 18 View FIGURES 15–18 ): coxa 0 0/1, 0/0 0, trochanter 1 1/1, 0/1 1 (av inserted on small tubercles), femur 1 2/1, 1/0 1, genu 2 2/1, 3/0 1 (posterior dorsal setae inserted on small tubercles), tibia 2 1/1, 3/1 2. Tarsi II–IV with 18 setae (3 3/2, 3/2 3 + mv, md); with thickened some setae, see Figure 16–18 View FIGURES 15–18 . All legs with elongate pretarsi, bearing small paired claws, short paradactyli and rounded pulvilli.
Male and immatures. Unknown.
Etymology. The specific name of the new species is derived from the Latin adjective subsolanus , sub- (“under, below”) + sôlânus (“of the sun”), and is meaning eastern. This name refers to the species occurring in the easternmost part of Russia.
Remarks. McGraw & Farrier (1969) erected the genus Proctogastrolaelaps to accommodate a new species P. libris , which was described associated with the bark beetles in Virginia and North Carolina, USA. Farrier & Hennessey (1993) also included this species in their checklist of soil-inhabiting and free-living Mesostigmata (Acari: Parasitiformes) from North America. Since that time, there has been no other formal occurrence or recording of Proctogastrolaelaps in the world, and is now recorded in Far East of Russia for the first time, from soil-litter. The original description of the genus is brief and its illustrations are incomplete, lacking some important details, especially those concerning leg chaetotaxy and gnathosoma. Moraes et al. (2016) in the world catalogue of the families Ascidae , Blattisociidae and Melicharidae clarified the definitions of all melicharids genera but a satisfactory clear concept of the genus Proctogastrolaelaps has still not been achieved. The discovery of the second species of Proctogastrolaelaps allowed us to develop a concept of the genus as above and clarified significant morphological attributes in the genus by the inclusion of the new species.
The new species can be easily distinguished from P. libris by the following characteristics: dorsal shield with 42 pairs of setae, including 23 pairs of podonotal setae and 19 pairs of opisthonotal setae, R5 and R6 absent (vs. dorsal shield with 44 pairs of setae, including 21 pairs of podonotal setae, both R5 and R6 present in P. libris ); (2) pre-sternal area comprises with two spindle shaped platelets (vs. absent in P. libris ); (3) peritrematal shield well developed, fused anteriorly to dorsal shield, posteriorly free (vs. peritrematal shield posteriorly fused to exopodal shield beside coxa IV in P. libris ); (4) genital shield expansive, broadened posteriorly, angled posterolaterally behind coxae IV (vs. genital shield not expansive and angled posterolaterally behind coxae IV in P. libris ); (5) Fixed digit of chelicera with five various sizes of teeth (including two large subterminal teeth not aligned with others) (vs. fixed cheliceral digit with two teeth in addition to apical tooth in P. libris , two large subterminal teeth absent); (6) soft opisthogastric cuticle with ten pairs of smooth setae inserted on small sclerotised rings around basal sockets (vs. opisthogastric cuticle setae without small sclerotised rings around basal sockets in P. libris ). Although, this genus has not been recorded from any part of the world except the Far East of Russia (this study) and USA ( McGraw & Farrier, 1969; Farrier & Hennessey, 1993), possibly as a result of a poor sampling effort for melicharid mites in other countries. But according to the collecting habitat of the new species, we can expand the occurrence habitat of this genus from galleries of bark beetles to soil-litter, however still scarce the knowledge about the feeding patterns or any other aspects of these mites behavior.
Nothing is known about the feeding behaviour of these mites or any other aspects of their biology. According to its normal morphology (like other free-living species of melicharids) (e.g. dentate chelicerae and sclerotised and horn-like corniculi, etc.), suspect that P. subsolanus is probably a predator of small invertebrates. We stress that further comprehensive field and experimental works are needed to find more species of this genus to establish the true role of this mite in its respective ecosystems.

| V |
Royal British Columbia Museum - Herbarium |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |