
Aselliscus dongbacana, Tu & Csorba & Görföl & Arai & Son & Thanh & Hasanin, 2015
|
publication ID |
https://doi.org/10.3161/15081109ACC2015.17.2.002 |
|
DOI |
https://doi.org/10.5281/zenodo.4323736 |
|
persistent identifier |
https://treatment.plazi.org/id/4F2D9C7D-6108-FFFE-4BDE-FB4AA958FD35 |
|
treatment provided by |
Carolina |
|
scientific name |
Aselliscus dongbacana |
| status |
sp. nov. |
Aselliscus dongbacana sp. n.
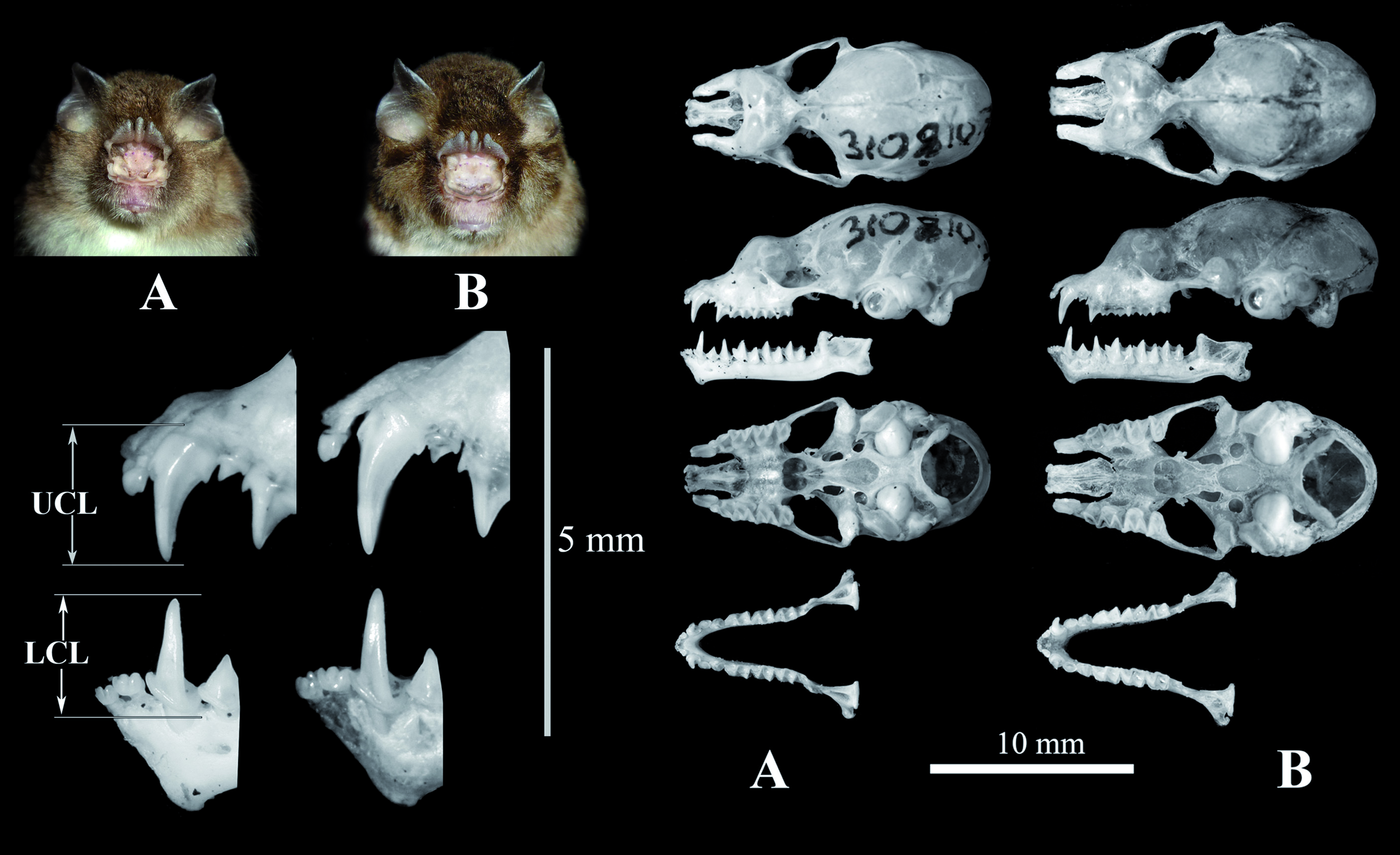
( Fig. 5B View FIG )
Holotype
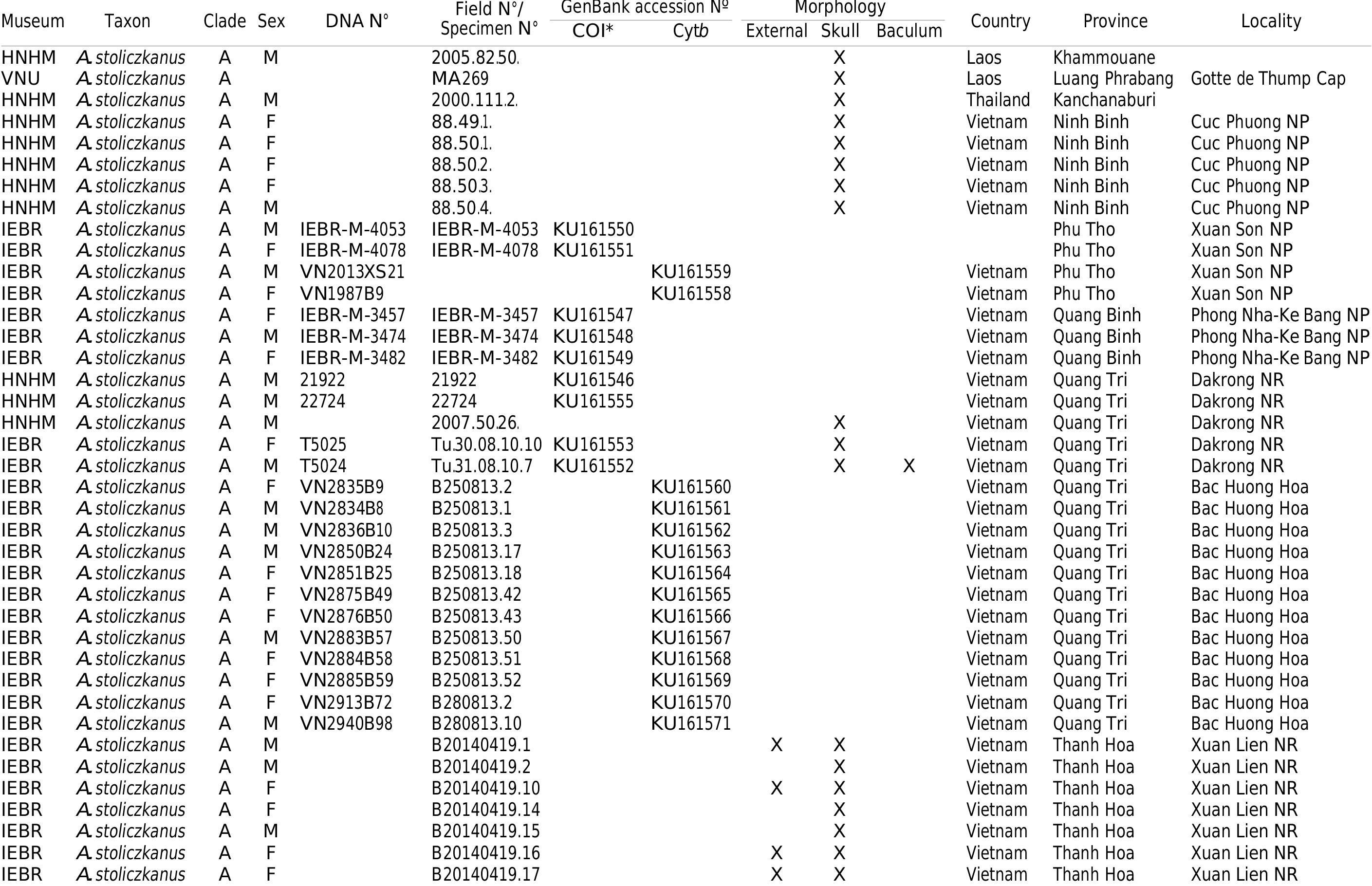
IEBR-VN11-0143 (Field no.: Tu.230511.1, tissue code: VN11-0143), adult ♂, body in alcohol, skull and baculum removed, collected by V. T. Tu on 23 May 2011. Mass: 4.5 g. Measurements (in mm) are as follows: FA: 43.8; Head and body length: 40.5; Tail: 39.5; Ear length: 12.2; Tibia: 19.7; 3 rdmt: 32.5; 3rd 1: 15.7; 4 thmt: 31.5; 4th 1: 13.2, cartilage: bifurcate; and 5 thmt: 27.9, 5th 1: 13.1, cartilage: bifurcate. GLS: 14.94; CCL: 13.01; C 1 C 1: 3.57; M 3 M 3: 5.55; ZB: 7.61; MB: 7.29; BW: 6.05; CM 3: 5.28; ML: 9.42; CM 3: 5.66; UCL: 1.51; and LCL: 2.01. The sequence of COI has been deposited in the EMBL/GenBank/DDBJ nucleotide databases with accession no. KU161543 View Materials . GoogleMaps
Type locality
Na Phong cave GoogleMaps , Ba Be National Park, Bac Kan province, Vietnam ( 22°23’N, 105°36’E; entrance altitude: 280 m a.s.l.).
Paratypes
IEBR-VN 11-0124 (Field no.: Tu.20.05.11.2; adult ♂; accession no. of COI sequence: KU161541 View Materials ) ; IEBR-VN 11-0125 (Field no.: Tu.20.05.11.3; adult ♂; accession no. of COI sequence: KU161542 View Materials ) ; IEBR-VN 11-0146 (Field no.: Tu.23.05.11.4; adult ♂; accession no. of COI sequence: KU161545 View Materials ) ; bodies in ethanol, skulls extracted; IEBR-VN 11-0115 (Field no.: Tu.19.05.11.2; adult ♀; accession no. of COI sequence: KU161539 View Materials ), IEBR-VN 11-0118 (Field no.: Tu.19.05.11.5, adult ♀; accession no. of COI sequence: KU161540 View Materials ), IEBR- VN11-0144 (Field no.: Tu.23.05.11.2; adult ♂; accession no. of COI sequence: KU161544 View Materials ), bodies in ethanol, collected from same location as holotype. HNHM 2007.27.9., adult ♂, body in ethanol, skull removed, accession no. of COI sequence: KU161556 View Materials , collected in Ba Be National Park by N. M. Furey and G. Csorba on 02 May 2007 .
Referred material
A series of other specimens identified as clade B collected from Na Hang Nature Reserve, Tuyen Quang province, Vietnam, Khau Ca Nature Reserve, Ha Giang province, and Phia Oac-Phia Den Nature Reserve, Cao Bang province, Vietnam are also referred to this species ( Appendix I View APPENDIX ). All of these specimens are deposited in the IEBR and in the HNHM. Bats identified as A. stoliczkanus were previously recorded at Kim Hy Nature Reserve, Bac Kan province ( Furey et al., 2009, 2010, 2011); these specimens should be allocated to A. dongbacana because this area is situated in the distribution range and just ca. 50 km away from the type locality (Ba Be National Park) of the new species.
Etymology
The specific epithet refers to the restricted distribution range of the new species, called ‘Đông Bắc’ in Vietnamese. Its proposed English name is ‘Dong Bac’s trident bat’ and Vietnamese name is ‘DƠi mũi ba lá Đông Bắc’.
Diagnosis
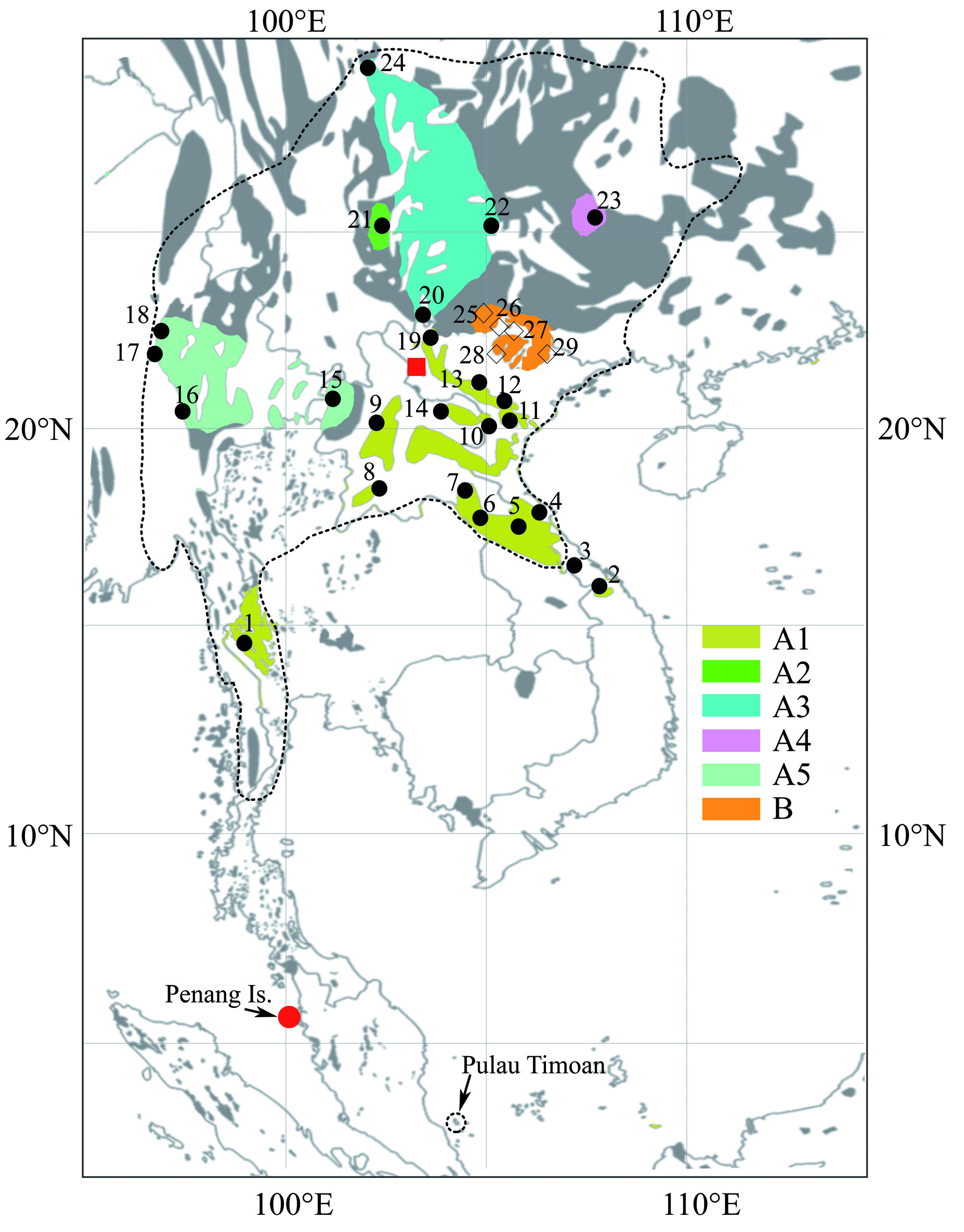
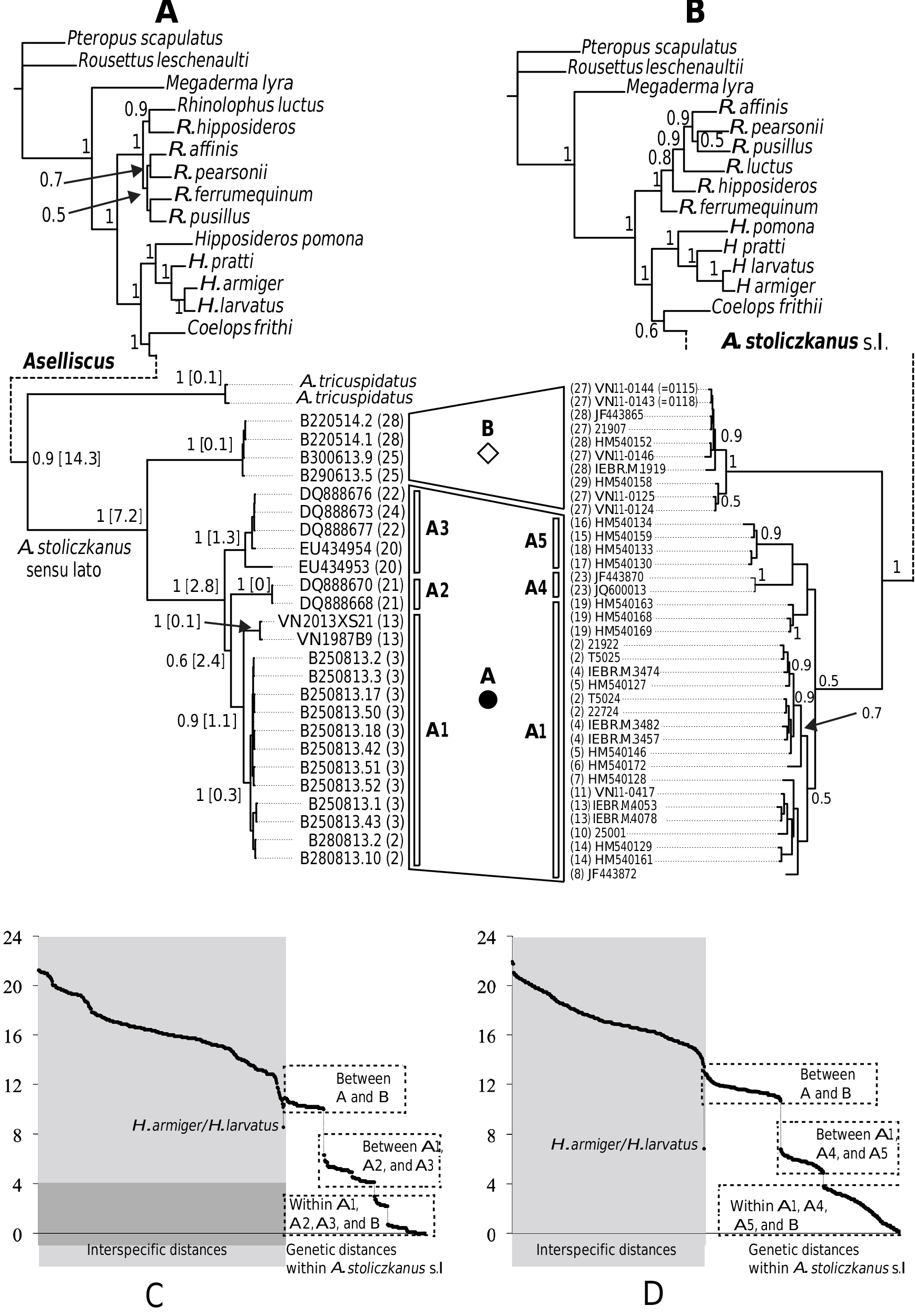
A member of the A. stoliczkanus complex comprising all specimens found in northeastern Vietnam ( Fig. 1 View FIG ) with a FA of ca. 42.8 mm, a GLS of ca. 15.2 mm ( Table 1 View TABLE ). The noseleaf is characterized by an upper margin divided into three points, and three lateral leaflets ( Fig. 5 View FIG ). The pelage is characterized by long and soft hairs, brown or reddish brown on the dorsum and grey or white-grey on the belly. The ears are small and pointed ( Fig. 5 View FIG ). The rostrum is sloping and elongated. The sagittal crest is relatively developed. The upper toothrows are convergent anteriorly. The upper incisors are bilobed. The upper and lower canines have low posterior cusps and are relatively robust with a length of ca. 1.95 mm and ca. 1.51 mm, respectively. The upper anterior premolar (PM 2) is compressed. The M 3 is scarcely reduced ( Fig. 5 View FIG ). COI and Cyt b sequences differ from the other species of the genus Aselliscus by> 10%.
Description
Externally, this is a small species with a FA of ca. 42.8 mm. The upperparts are buffy brown to greyish-brown; the underparts are pale to buffy white. The noseleaf structure is characterized by an upper margin divided into three points, and three lateral leaflets. The ears are small and pointed. ( Fig. 5 View FIG ). The cartilage of the fourth and fifth metacarpal is bifurcate.
The skull of the new species is small with a GLS of ca. 15.2 mm. The rostrum is sloping and elongated. The sagittal crest is relatively developed. The anteriors of the zygoma have a well-developed jugal projection. The upper toothrows are convergent anteriorly. The upper incisors are bilobed. The upper and lower canines have low posterior cusps; the upper anterior premolar (PM 2) is compressed. The M 3 is scarcely reduced ( Fig. 5 View FIG ).
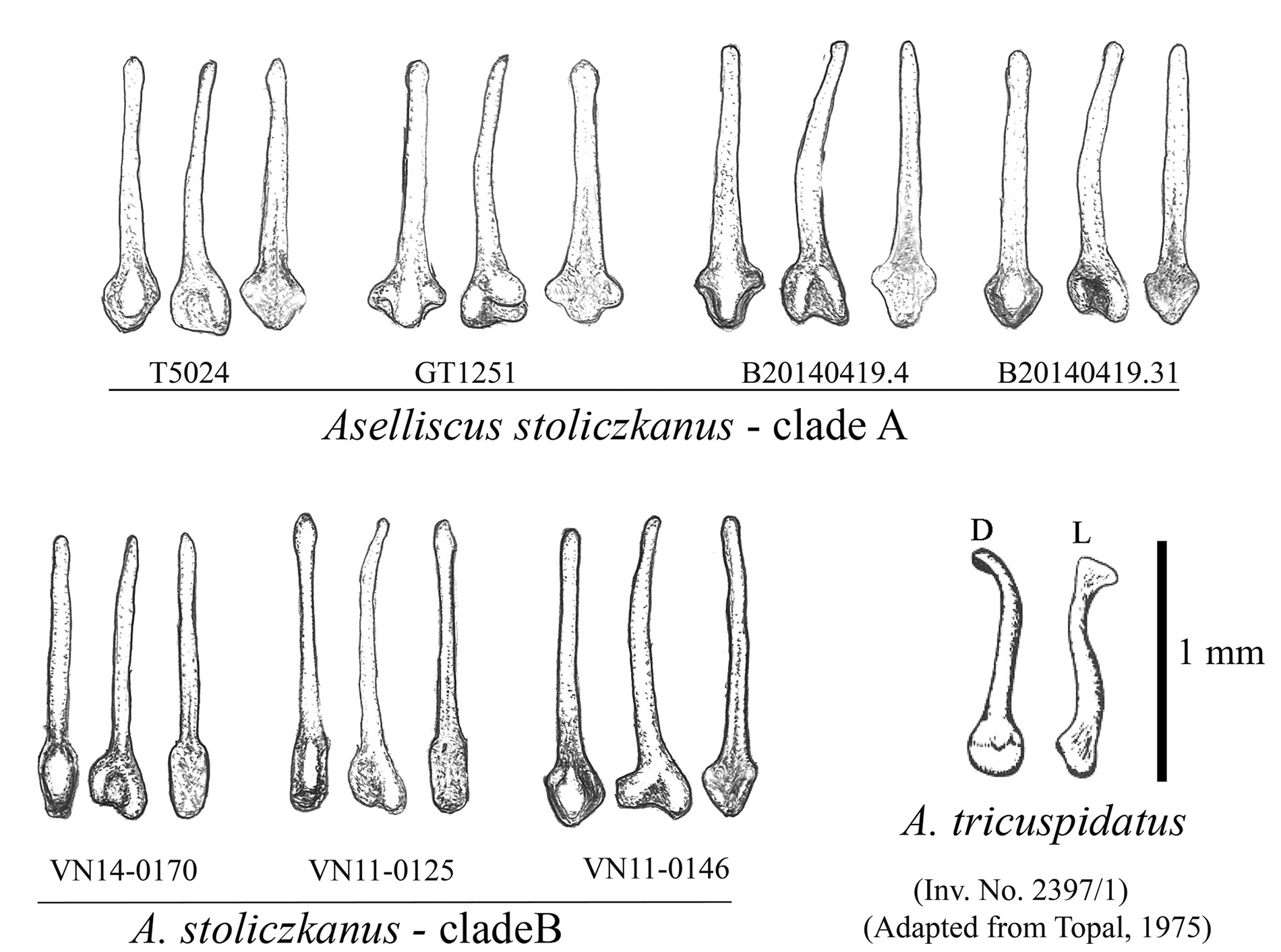
The baculum of the new species is bow-shaped or relatively straight in lateral view. The basal portion is widened with two lateral lobes. The shaft tapers slightly from the basal portion to the blunt tip ( Fig. 3 View FIG ).
Comparisons with other species
In its morphological characters, A. dongbacana differs significantly from A. tricuspidatus by external, craniodental, and baculum features as well as its disjunct geographical distribution. As compared to A. stoliczkanus s.l., the new species is significantly different in size from the holotypes of A. stoliczkanus and A. trifidus ( Table 1 View TABLE , Figs. 1 View FIG , 4A, and 4 View FIG C– 4E). The external and bacular characters of A. dongbacana greatly overlap with those of A. stoliczkanus s.l. found in Indochina and Southern Thailand (including the type series of A. wheleeri ), but the average of most craniodental measurements of the new species are significantly larger than those of the latter. The upper and lower canines of A. dongbacana are also significantly longer and more robust than those of the others ( Table 1 View TABLE and Fig. 5 View FIG ).
As for the acoustic characters, Furey et al. (2009) reported that the echolocation calls of A. dongbacana found at Kim Hy Nature Reserve, Bac Kan province are characterized by a typical constant frequency followed by frequency modulated (CF/FM) signal, with a frequency of maximum energy (FmaxE) of 127.5±2.6 kHz ( n = 5). Li et al. (2007) found that Chinese A. stoliczkanus s.l. emits calls with a relatively low FmaxE, e.g. in Sichuan and Guizhou the average frequency is 120.3±0.3 kHz ( n = 10) and the range of values in Yunnan is 118.4–119.3 kHz. In Myanmar, Khin (2012) recorded an FmaxE of 126.68 ± 4.36 kHz for A. stoliczkanus s.l., whereas the FmaxE of the A. tricuspidatus ssp. collected in YUS Conservation Area, Papua New Guinea is around 115 kHz ( Robson et al., 2012).
Genetics
The Cyt b and COI sequences of A. dongbacana sp. n. differ from those of A. stoliczkanus s.l. and A. tricuspidatus by> 10.0% ( Fig. 2 View FIG and Appendix III).
Distribution
The species is currently known only from karst areas in Northeastern Vietnam ( Fig. 1 View FIG ).
Ecology and habitat
Like other Aselliscus species, A. dongbacana sp. n. is also associated with karst areas, and use caves as roosts both in heavily degraded and intact limestone habitats. So far, nothing is known on the diet of A. dongbacana sp. n., but they might forage on small nocturnal insects in dense environments like A. stoliczkanus sensu stricto (s.s.) does ( Li et al., 2007). However, the differences in skull size and especially in canine length suggest that their food sources may be different. Further studies on the diet of the two taxa is essential for a better understanding of whether food sources are important factors in their diversification. During our surveys, several pregnant females of A. dongbacana sp. n. were captured in May, while lactating females were found in June. These observations confirm that March–July is the primary reproductive period for the new species and also for other insectivorous bats in North Vietnam ( Furey et al., 2011).
Conservation status
To date, A. stoliczkanus s.l. has been classified as Least Concern in the IUCN Red List ( Bates et al., 2008). However, A. dongbacana sp. n. is endemic to northeast Vietnam and little is known about the current population trends of the species. Unfortunately, like many other regional plants and animals, A. dongbacana sp. n. might be at risk due to various types of roost and habitat destruction, i.e. mining, timber harvesting or cave tourism ( Day and Urich, 2000; Clements et al., 2006; Furey et al., 2010). Further studies are needed to assess the impacts of habitat changes on A. dongbacana sp. n. to identify high priority conservation areas to protect the species ( Hutson et al., 2001; Furey et al., 2010; Kingston, 2010).
The speciation of Aselliscus in mainland Southeast Asia: when and how?
Our molecular dating based on Cyt b sequences indicates that the separation between A. dongbacana sp. n. and A. stoliczkanus s.s. took place during the late Miocene (ca. 7.2 Mya), much earlier than the diversification among subclades of A. stoliczkanus s.s. around the Plio-Pleistocene boundary (ca. 2.8– 2.4 Mya — Fig. 2 View FIG and Appendix IV). The period of interspecific divergence seems therefore to coincide with the hypothetic climatic and associated vegetation changes in the region during the late Miocene. Indeed, at the beginning of the late Miocene (ca. 10–8 Mya or more recently), the extent and uplift of the Himalayan mountains and the Tibetan Plateau, linked to the development of the Northern Hemispheric ice sheets played an important role in driving the Asian aridification ( An, 2000; An et al., 2001; Zhang Y. G. et al., 2009). As a consequence, the cool, dry climate caused the vegetation to change from mixed coniferous and broad-leaved forests to grasslands in Asia, and rainforests of the region were thought to be compressed into different refugia ( Morley, 2000; An et al., 2001). At the end of the late Miocene and until the early Pliocene epochs, Southeast Asia was a single block of rainforest, as a consequence of the warm and humid climatic conditions. However, the uplift of Himalaya- Tibetan plateau about 3.6–2.6 Mya and the onset of extensive glaciations on the Northern Hemisphere during the late Pliocene and Pleistocene epochs, led to the development of more open vegetation types and the contraction of the rainforest into several isolated refugia ( Morley, 2000; An et al., 2001; Meijaard and Groves, 2006). With this in mind, the current distribution of Aselliscus spp. in Mainland Southeast Asia ( Fig. 1 View FIG ) suggests that their separation probably occurred in different glacial refugia across the region during two major phases of aridification in Asia since the late Miocene. Aselliscus bats are very small (body mass ca. 5 g), fly at low speeds and are usually associated with karst areas and forage in cluttered habitats ( Li et al., 2007; Francis, 2008). These morphological and ecological features indicate that they might have poor dispersal capacities and high natal philopatry that could prevent gene flow among different isolated populations and facilitate speciation events. Despite their long separation, these taxa were found to have similar morphology and echolocation call features; whereas previous studies indicated that different species of hipposiderid bats are usually recognizable by their call features (i.e., Kingston et al., 2001; Thong et al., 2012). However, given that Aselliscus spp. are associated with karst areas, we hypothesize that their ecological evolution might be under stabilizing selection imposed by the special environmental conditions of karst habitats (i.e., forests and caves) ( Bickford et al., 2007) and consequently reduces morphological and acoustic variation between different taxa.

| HNHM |
Hungary, Budapest, Hungarian Natural History Museum |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |