
Artema nephilit, Aharon & Huber & Gavish-Regev, 2017
|
publication ID |
https://doi.org/ 10.5852/ejt.2017.376 |
|
publication LSID |
lsid:zoobank.org:pub:F67AE00B-28CF-48AF-89D6-31251B22BB7C |
|
DOI |
https://doi.org/10.5281/zenodo.3852326 |
|
persistent identifier |
https://treatment.plazi.org/id/8C0CA94A-B5CF-4585-B51D-FF25711103E5 |
|
taxon LSID |
lsid:zoobank.org:act:8C0CA94A-B5CF-4585-B51D-FF25711103E5 |
|
treatment provided by |
Carolina |
|
scientific name |
Artema nephilit |
| status |
sp. nov. |
Artema nephilit View in CoL sp. nov.
urn:lsid:zoobank.org:act:8C0CA94A-B5CF-4585-B51D-FF25711103E5
Figs 1 View Figs 1–2. 1 , 4–6 View Figs 3–8 , 9–14 View Figs 9–14 , 35–75 View Figs 35–44 View Figs 45–54 View Figs 55–66 View Figs 67–75 , 203 View Figs 201–207 , 210 View Figs 208–214
“ Artema mauriciana View in CoL ” (misidentification) – Bodenheimer 1937: 238 (“ Palestina ”); see Notes below.
“ Artema mauricia ” (misidentification) – Dalmas 1920: 59 (Bodrum, Turkey).
Diagnosis
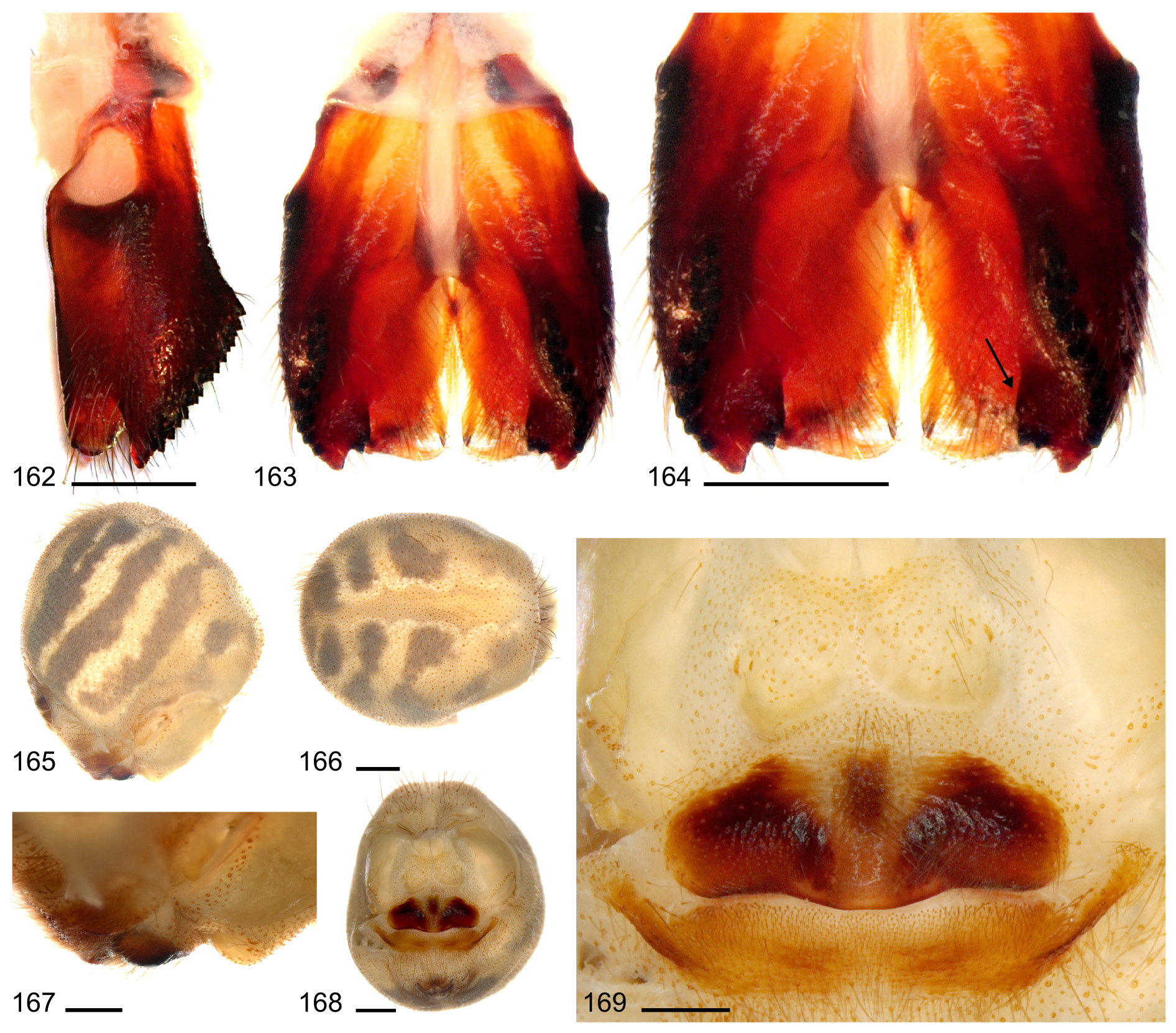
Males can be distinguished from all known congeners by their bulbal processes: process c ( Fig. 40 View Figs 35–44 ) projecting prolaterally, processes d and e absent ( Fig. 39 View Figs 35–44 ) ( A. magna : process c robust, strongly curved prolaterally, process d distinct rounded projection on ventral side of bulb – see Figs 159–160 View Figs 155–161 ; A. doriae and A. transcaspica : process d small, pointed towards ventrodistally) and by unique median projection on each male cheliceral process ( Figs 43–44 View Figs 35–44 , 67 View Figs 67–75 ) (only A. magna with similar median projection but no modified hairs connect to main ridge as in A. nephilit sp. nov. – see Figs 163–164 View Figs 162–169 ). Females with semicircular epigynum ( Figs 45–50 View Figs 45–54 ); differing from A. atlanta by straight posterior epigynal margin; from A. magna by epigynal plate length to width ratio; from A. bunkpurugu by much less prominent anterior epigynal projections (AEP in Fig. 48 View Figs 45–54 ) (cf. Huber & Kwapong 2013: figs 49, 53–54).
Etymology
The species epithet is derived from the feminine singular noun of the biblical name “Nephilim”, the giants who were seen by the twelve people sent by Moses to scout the Land of Canaan. It refers to the large size of the spider. Noun in apposition.
Material examined
Holotype
ISRAEL: ♂, Northern District, Jordan Rift Valley, Mount Berniki, south Teverya ( Tiberias ) (32.777° N, 35.541° E), in cave, - 120 m b.s.l., 16 Sep. 2013, B.A. Huber, S. Aharon and E. Gavish-Regev leg. ( HUJ Ara 16000).
GoogleMapsOther material
ISRAEL: 3 ♀♀, 1 juv. together with holotype, same data ( HUJ Ara 15996–99); 1 ♂, 1 ♀, same data but deep in cave ( ZFMK Ar 15218–19); 1 ♀ ( HUJ Ara 16001), 1 juv. ( HUJ Ara 16002), same data; 1 ♀, HaTanur Waterfall , near Metula (33.269° N, 35.581° E), 15 Mar. 2011, L. Friedman leg. ( CSZ); GoogleMaps 2 ♂♂, 1 ♀, Oren Cave, Mount Karmel (32.714° N, 34.975° E), in cave, 70 m a.s.l., about 3 km from Mediterranean Sea , 17 Sep. 2013, B.A. Huber, E. Gavish-Regev and S. Aharon leg. ( ZFMK Ar 15225– 26); GoogleMaps 1 ♂, 1 ♀, 1 juv., En Perat, Nahal Perat (31.833° N, 35.303° E), in caves and a building, 260–300 m a.s.l., 15 Sep. 2013, B.A. Huber, S. Aharon and E. Gavish-Regev ( ZFMK Ar 15220–21); GoogleMaps 1 ♀, 1 juv., in pure ethanol, same data ( ZFMK Isr 51); 1 ♀, Ein ad-Duyuk, near Jericho (31.895° N, 35.421° E), 25 May 1983, P. Amitai leg. ( HUJ Ara 16003); GoogleMaps 8 juvs, Qumeran Cave (31.741° N, 35.459° E), 7 Nov. 2012, L. Friedman leg. ( SMNH); GoogleMaps 1 ♀, in pure ethanol, Nahal Dawid, En Gedi (31.470° N, 35.390° E), among rocks, -250 to - 300 m b.s.l., 10 Sep. 2013, B.A. Huber, S. Aharon and E. Gavish-Regev leg. ( ZFMK Isr 58); GoogleMaps 1 ♀, same data ( HUJ Ara 16004); 1 ♂, 1 ♀, En Gedi (31.470° N, 35.390° E), 20 Sep. 1975, B. Feldman leg. ( HUJ Ara 16005-6); GoogleMaps 1 juv., same locality, 29 May 1968, P. Amitai leg. ( HUJ Ara 16007); 1 ♀, in pure ethanol, Ma’ale Ya’ir (31.215° N, 35.359° E), under large rocks, - 370 m b.s.l., 9 Sep. 2013, B.A. Huber, S. Aharon and E. Gavish-Regev ( ZFMK Isr 49); GoogleMaps 2 ♀♀, Nahal Temarim (31.640° N, 35.411° E), under large rocks, 3 Sep. 2013, S. Aharon leg. ( HUJ Ara 16008-9); GoogleMaps 2 ♀♀, 1 juv., Sedom Cave (31.087° N, 35.395° E), - 370 m b.s.l., 15 Aug. 1939, A. Shulov leg. ( HUJ Ara 16010-12); GoogleMaps 1 ♀, in pure ethanol, same locality, 10 Sep. 2013, B.A. Huber, S. Aharon and E. Gavish-Regev ( ZFMK Isr 47); 1 ♂ ( HUJ Ara 16013), 1 ♀ ( ZFMK Ar 15222), same data; 2 ♀♀, same locality, 30 Oct. 1941 ( CSZ); 3 juvs, Sedom Spring, 15 Aug. 1957, H. Ginsburg leg. ( HUJ Ara 16014–16); 3 ♀♀, Arubotaim Cave (31.101° N, 35.390° E), - 380 m b.s.l., 10 Sep. 2013, B.A. Huber, S. Aharon and E. Gavish-Regev leg. ( ZFMK Ar 15223); GoogleMaps 1 ♀, 2 juvs, in pure ethanol ( ZFMK Isr 46), 1 juv. ( HUJ Ara 16017), same data; 1 ♀, Malham Cave (31.076° N, 35.397° E), - 370 m b.s.l., 10 Sep. 2013, B.A. Huber, S. Aharon and E. Gavish-Regev leg. ( HUJ Ara 16018); GoogleMaps 1 ♀ ( ZFMK Isr 44), 1 juv. (HUJ 16038), in pure ethanol, same data; 1 ♀, NW of ’En Tamar, Nahal Zin (30.989° N, 35.346° E), in crevices, - 340 m b.s.l., 9 Sep. 2013, S. Aharon, E. Gavish-Regev and B.A. Huber leg. ( HUJ Ara 16019); GoogleMaps 1 ♂, molted to adult 30 Oct. 2013, NW of ’En Tamar, Nahal Zin (30.991° N, 35.347° E), in tube near road tunnel, - 340 m b.s.l., 9 Sep. 2013, B.A. Huber, S. Aharon and E. Gavish-Regev leg. ( ZFMK Ar 15224); GoogleMaps 1 ♀, Ramat Avedat, near Sede Boqer , Negev Desert [~ 30.84° N, 34.77° E], Jan. 1987, V. and B. Roth leg. (MCZ 34068); GoogleMaps 1 ♂, 1 ♀, Kibbutz Samar (29.831° N, 35.022° E), dumping ground, 100 m a.s.l., 11 Sep. 2013, S. Aharon, E. Gavish-Regev and B.A. Huber leg. ( HUJ Ara 16020–21); GoogleMaps 1 ♀, in pure ethanol, same locality, 19 Oct. 2013, E. Gavish-Regev leg. ( HUJ Ara 16022); 1 ♂, 2 ♀♀, in 80% ethanol, same data, in houses ( HUJ Ara 16023–25); 1 ♂, 1 ♀, Eilat Field School buildings (29.509° N, 34.920° E), 12 Oct. 2013, S. Aharon and E. Gavish-Regev leg. ( HUJ Ara 16026-27); GoogleMaps 2 juvs, in pure ethanol, same data ( HUJ Ara 16028–29).
GREECE: 1 ♀, Athens, Acropolis [37.97° N, 23.72° E], collection Roewer , date and collector not given ( SMF RII /3724). GoogleMaps
TURKEY: 1 ♂, 2 penultimate males, Bodrum [37.03° N, 27.42° E], 16 Sep. 1919, R. Varriale leg. ( MSNG); GoogleMaps 1 ♀, Antalya Prov., Kemer District, Çıralı (36.399° N, 30.475° E, 10 m a.s.l.), in building near beach, 3 Aug. 2016, B.A. Huber leg. ( ZFMK, collected as juvenile, molted to adult in Jul. 2017, still alive as of Nov. 2017). GoogleMaps
CYPRUS: 1 ♂, Nicosia [35.16° N, 33.36° E], 1937, F. Werner leg. ( NHMW 19.551); GoogleMaps 1 ♂, 2 juvs, Nicosia, near Agios Sozomenos (35.059° N, 33.421° E), 180 m a.s.l., in cave, 5 Jul. 2016, M. Hadjiconstantis leg. ( CMH); GoogleMaps 1 ♂, 2 ♀♀, same data but 20 Jan. 2016 ( CMH); 3 ♀♀, Nicosia, Geri (35.102° N, 33.417° E), 150 m a.s.l., in house, 15 Sep. 2016, M. Hadjiconstantis leg. ( CMH); GoogleMaps 1 ♂, 2 juvs, Famagusta, Lisi, (35.106° N, 33.680° E), 60 m a.s.l., in Panagia Lysis church, 15 Apr. 2016, M. Hadjiconstantis leg. ( CMH). GoogleMaps
JORDAN: Aqaba Province: 2 ♂♂, Wadi Rām (29.7405° N, 35.4574° E), among rocks, 830 m a.s.l., 12 Sep. 2013, B.A. Huber leg. ( ZFMK Ar 16161-62), collected as juveniles, molted to adults in Dec. 2015 and Sep. 2016. – GoogleMaps Ma’an Province: 2 ♂♂, 2 ♀♀, Petra (30.324° N, 35.447° E), 900–950 m a.s.l., rock-cut tombs, near theater, 13 Sep. 2013, B.A. Huber leg. ( ZFMK Ar 15227); GoogleMaps 1 ♀, 1 juv., in pure ethanol, same data ( ZFMK Isr 36); 1 ♂, same data but among rocks ( ZFMK Ar 15228). – Karak Province: 1 ♀, 1 juv., Wadi Hasa (31.004°– 31.014° N, 35.494 –35.506 ° E), -330 to - 250 m b.s.l., among rocks, 14 Sep. 2013, B.A. Huber leg. ( ZFMK Ar 15229–30); GoogleMaps 1 ♀, in pure ethanol, same data ( ZFMK Isr 63). – Madaba Province: 1 ♂, 1 juv., Wadi Mujib (31.465° N, 35.578° E), - 380 m b.s.l., among rocks, 14 Sep. 2013, B.A. Huber leg. ZFMK ( Ar 15231). GoogleMaps
Material assigned tentatively (see Notes below)
YEMEN: 1 ♀, “ Yar Ju Amlah ” [= Al Ja’milah?, 17.11° N, 43.56° E], 1950, M. Borri and M. Poggesi leg. ( MZUF 128 ). GoogleMaps
UNITED ARAB EMIRATES: 1 ♀, Near Ra’s al-Khaimah (25.46º N, 55.96º E), hand-collected, 8 Mar. 2005, A. van Harten leg. ( ZFMK Ar 15232). GoogleMaps
Description
Male (holotype)
MEASUREMENTS. Total body length 7.0, carapace width 3.1. Leg 1: 48.4 (11.9 + 1.4 + 13.9 + 18.3 + 2.9), tibia 2: 10.2, tibia 3: 8.0, tibia 4: 10.9; tibia 1 L/d: 35. Distance PME–PME 190 μm, diameter PME 190 μm, distance PME–ALE 90 μm, distance AME–AME 50 μm, diameter AME 170 μm.
COLOR. Carapace pale to light beige, with dark gray median band. Clypeus with wide light brown band (as in Figs 4 View Figs 3–8 , 9–14 View Figs 9–14 , 35 View Figs 35–44 ). Legs pale yellow to light ochre with dark rings on femora subdistally, patellae + tibiae proximally, and tibiae subdistally, tips of femora and tibiae whitish; sternum pale, with narrow light brown margins. Abdomen light brown with white and dark dots forming stripes from dorsal to lateral side (as in Figs 51–52 View Figs 45–54 ).
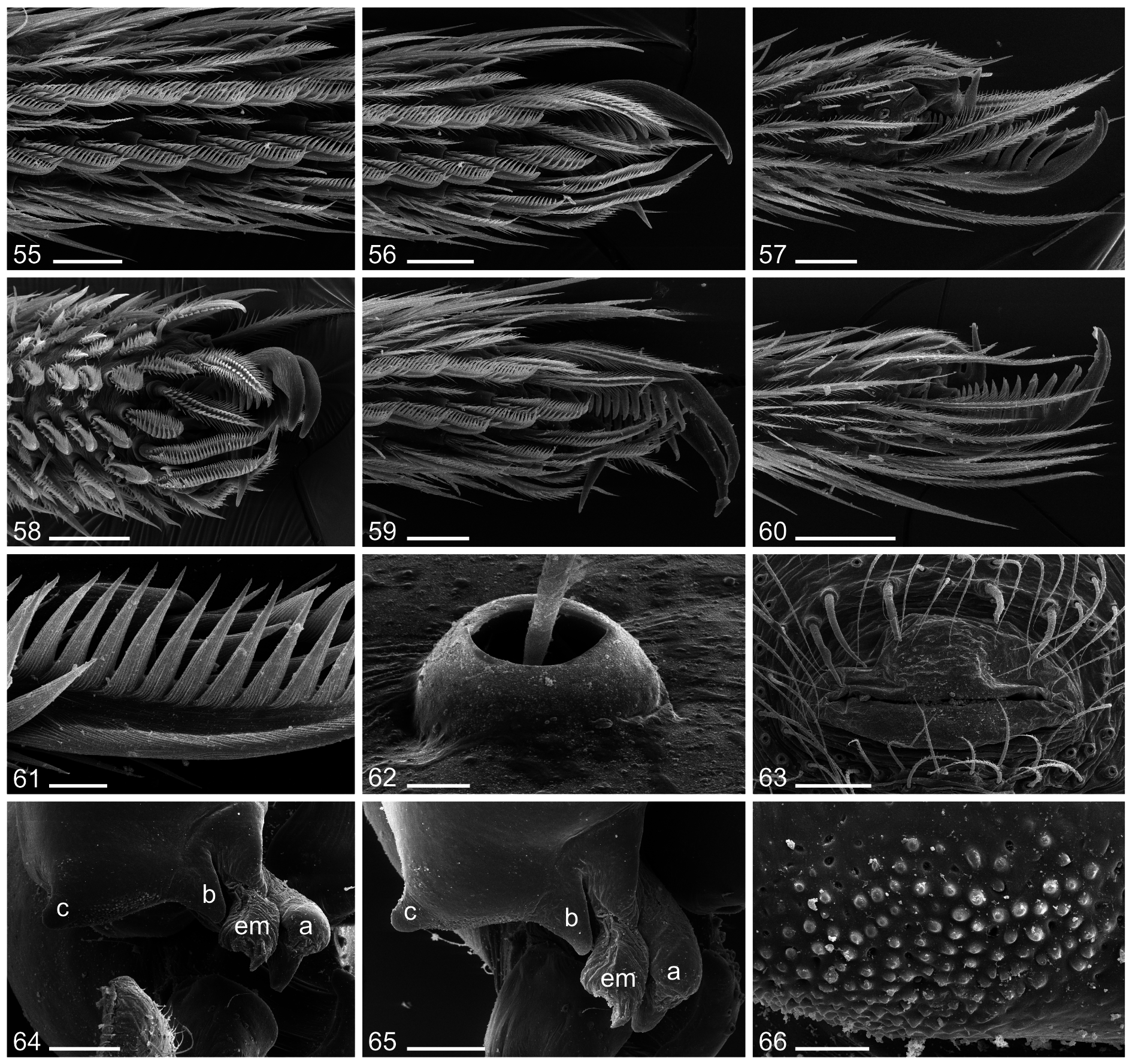
BODY. Ocular area slightly elevated. Carapace with median pit and distinctive posterior furrow. Clypeus unmodified. Sternum wider than long (2.3/1.5). Chelicerae as in Figs 42–44 View Figs 35–44 and as in Figs 67–69 View Figs 67–75 , with frontal row of ~20 modified (cone-shaped) hairs on each side that splits distally, with outer branch ending at tip of process, inner branch ending at tip of median projection; with indistinct small field of stridulatory ridges laterally (as in Fig. 70 View Figs 67–75 ). Abdomen globose and high; gonopore with four epiandrous spigots.
PALPS. As in Figs 36–38 View Figs 35–44 . Coxa unmodified. Trochanter with short ventral projection. Femur with short, somewhat pointed retrolateral process proximally; ventral membranous area proximally bordered on both sides by sclerotized ridges; femur-patella hinges close together dorsally. Patella very short. Procursus with proximal dorsal process and weakly developed ventral pocket; with distal dorsal notch on prolateral margin (as in Fig. 40 View Figs 35–44 ). Bulb with two processes close to membranous embolus (processes a and b, as in Fig. 40 View Figs 35–44 ), process c (as in Fig. 40 View Figs 35–44 ) strongly projecting prolaterally, and indistinct projection on ventral side of bulb (as in Figs 39 View Figs 35–44 , 64–65 View Figs 55–66 ). Area between processes b and c modified with granular surface (as in Fig. 66 View Figs 55–66 ).
LEGS.Without spines;with long curved hairs, especially on tibiae and metatarsi; retrolateral trichobothrium on tibia 1 at 8%; prolateral trichobothrium present on all tibiae; pseudosegmentation not visible.
Male (variation)
Tibia 1 in 14 other males: 9.2–20.6 (mean 14.4). Color pattern on abdomen varies from distinctive stripes consisting of black and white dots on pale to brown-ochre background to pale without any pattern (cf. females, Figs 52–53 View Figs 45–54 ); leg color varies from light brown to ochre; ocular area usually light brown to ochre; median band on carapace sometimes reaches posterior margin of ocular area, forming short
v-shaped mark. Cheliceral modified hairs vary slightly in numbers. Gonopore with variable number of epiandrous spigots (4–6) ( Fig. 63 View Figs 55–66 ).
Female
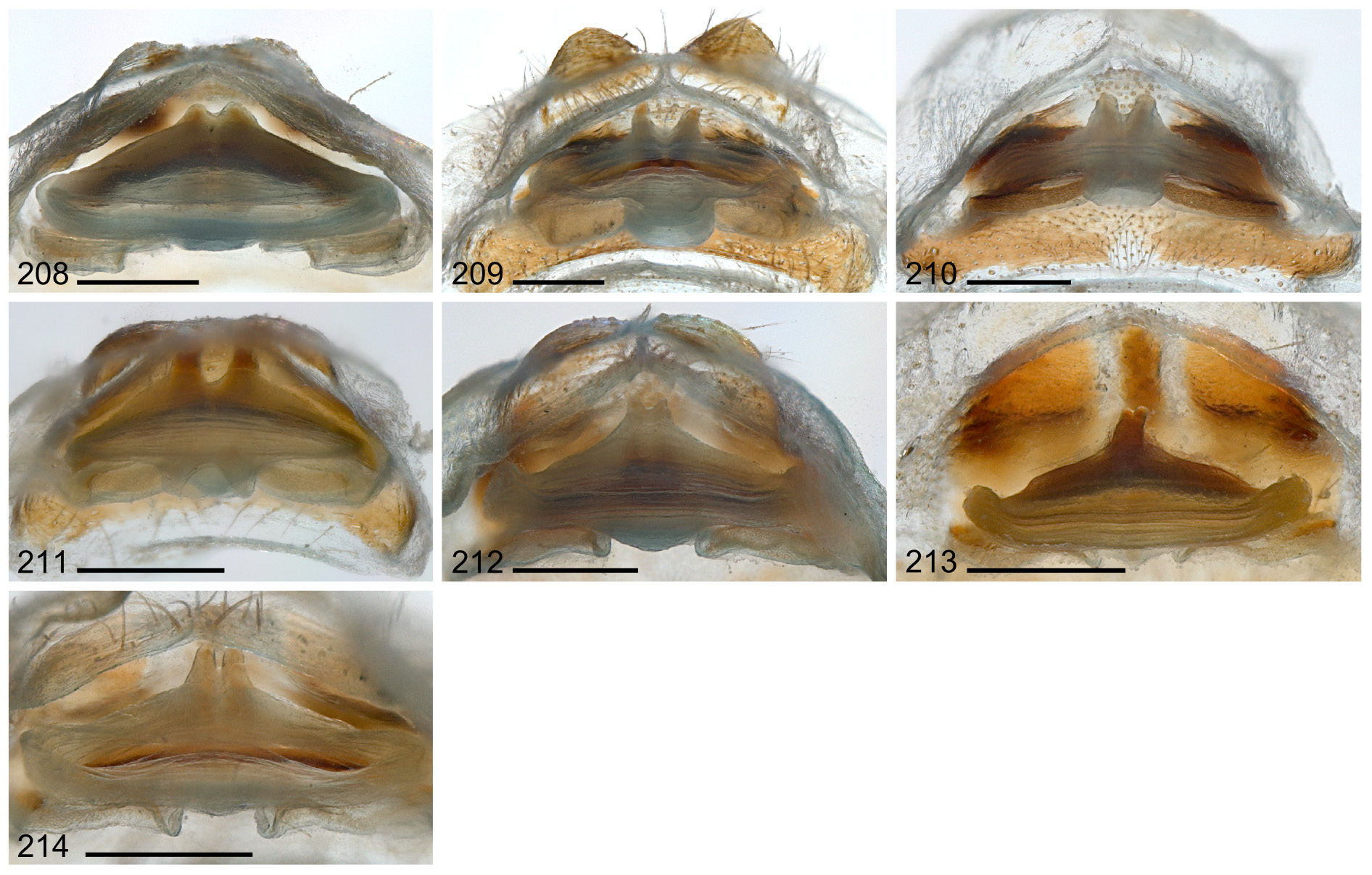
In general similar to male; tibia 1 in 26 females: 8.0–16.5 (mean 12.5); several females with light brown carapace margins and subtle brown stripes on ocular area. Stridulatory files laterally on chelicerae more distinct than in males ( Fig. 71 View Figs 67–75 ). Epigynum almost twice as wide as long, semi-circular, posterior margin almost straight, gently undulating; epigynal plate consisting of two sclerotized lateral areas that appear swollen posteriorly, pale median area, and dark median sclerite anteriorly (variably large and variably distinct) ( Figs 45–50 View Figs 45–54 ); pair of projections anterior to epigynum (AEP) visible ( Fig. 48 View Figs 45–54 ) but not prominent as in A. bunkpurugu (cf. Huber & Kwapong 2013: figs 49, 53–54).
Natural history
The spiders were found both in natural habitats such as caves, deep crevices, and under boulders, as well as in man-made habitats such as buildings, under low culverts, and artificial caves. In caves, the spiders built irregular webs (not clearly dome-shaped webs) near the ground, under stones, in crevices, and on the cave ceiling. The spiders seemed to avoid the deeper, dark parts of the visited caves. In a large cave survey we conducted in Israel ( Aharon 2016), we found A. nephilit sp. nov. in caves especially along the Jordan Rift Valley, in a very hot and arid zone with low mean annual precipitation (50–150 mm). Only a few specimens were found outside the Jordan Rift Valley, in caves in areas with higher mean annual precipitation (400–650 mm) in a more mesic habitat. These findings suggest that the species is more abundant in xeric, hot and dry caves rather than in humid caves nearby. The spiders were always found on the east or south slopes that are dryer than the west and north slopes and thus seem to be adapted to arid conditions ( Aharon 2016).
We examined more than 40 caves in Israel; 19 caves were inhabited by A. nephilit sp. nov., and most of these caves were dry. Some specimens were also found in hot, humid caves, but the species was never found in wet caves. In caves in southern Israel, where food abundance was low, we could not observe prey items. In a cave situated in the Golan Heights, in the north of Israel, where the annual precipitation is higher, we recorded remnants of isopods, tenebrionid beetles, small scorpions, and a spider. In the south of Israel, in a basement of the Eilat Field School, we observed remnants of adult males that had probably been killed by females (that were seen nearby), while in a cave in Nahal Perat, east of Jerusalem, we observed the predation of an adult male A. nephilit sp. nov. by Holocnemus pluchei (Scopoli, 1763) . Penultimate instar juveniles collected from caves in southern Israel (Arubatayim, Sedom, and Malcham) and reared independently in Bonn, Germany and Midreshet Ben-Gurion, Israel, molted to adult almost simultaneously after an unusually long period of 9–10 months (September 2013 to July 2014) (4 ♂♂ and 1 ♀ molted in Midreshet Ben-Gurion; 3 ♂♂ in Bonn). This simultaneous timing suggests that the long period is not an artifact caused by the food regime or differences from natural conditions, but rather reflects an intrinsic growth and maturation period.
A mating sequence was observed on 10 Jan. 2015 (in Bonn, Germany). Male from Israel, Sedom Cave; collected penultimate in September 2013; molted to adult in July 2014. Female from Israel, Mount Berenice; collected penultimate in September 2013; molted to adult in September 2014. Previous trials (with various males and females) performed in daylight had resulted in no male reaction when placed into the female’s web. Trials under red light resulted in male courtship, but the females, who had not recently fed, attacked the males. Two days after being fed with crickets, the female above finally accepted the male. Time specification shown as hr:min.
0:00 Male is gently transferred to female web.
0: 12 Male approaches female, bounces slightly and touches silk lines with front legs; female directs herself towards male, vibrates slightly a few times.
0: 16 Male touches female legs and approaches further ( Fig. 9 View Figs 9–14 ); female remains mostly still.
0:17 Final approach; male lowers palps below body ( Fig. 10 View Figs 9–14 ); searches for about 20 sec for correct position on female, apparently by searching for correct alignment of chelicerae relative to epigynum ( Fig. 11 View Figs 9–14 ); female abdomen is pushed backwards.
0:18 As soon as chelicerae are correctly placed (locked?) at epigynum, palps are rotated 180° around longitudinal axis (procursus moving outward and forward; Fig. 12 View Figs 9–14 ); insertion of palps, start of copulation; female releases legs from silk, pulls all legs above body and is moved by male to perpendicular final mating position.
0:19–1:02 No major movements, only very slight jerks of male body, usually every 6–8 seconds, rarely with shorter (4 sec) or longer (10 sec) intervals; no rhythmic palpal movements; no female movements; no female stridulation; asymmetric insertion of palps, consistent over entire copulation (left palp more deeply inserted than right palp; Fig. 13 View Figs 9–14 ).
1:02 Sudden separation, apparently initiated by male.
1:02–1: 12 Female remains motionless; male cleans palps for about 10 min ( Fig. 14 View Figs 9–14 ), then also motionless.
1:18 End of observation.
About 24 weeks later, this female produced an egg-sac. After another two weeks (i.e., beginning of August 2015), about 40 spiderlings emerged; two weeks later again they molted for the first time. In July 2017, i.e., almost two years after eclosion, most remaining specimens (about 15) molted to adults.
Distribution
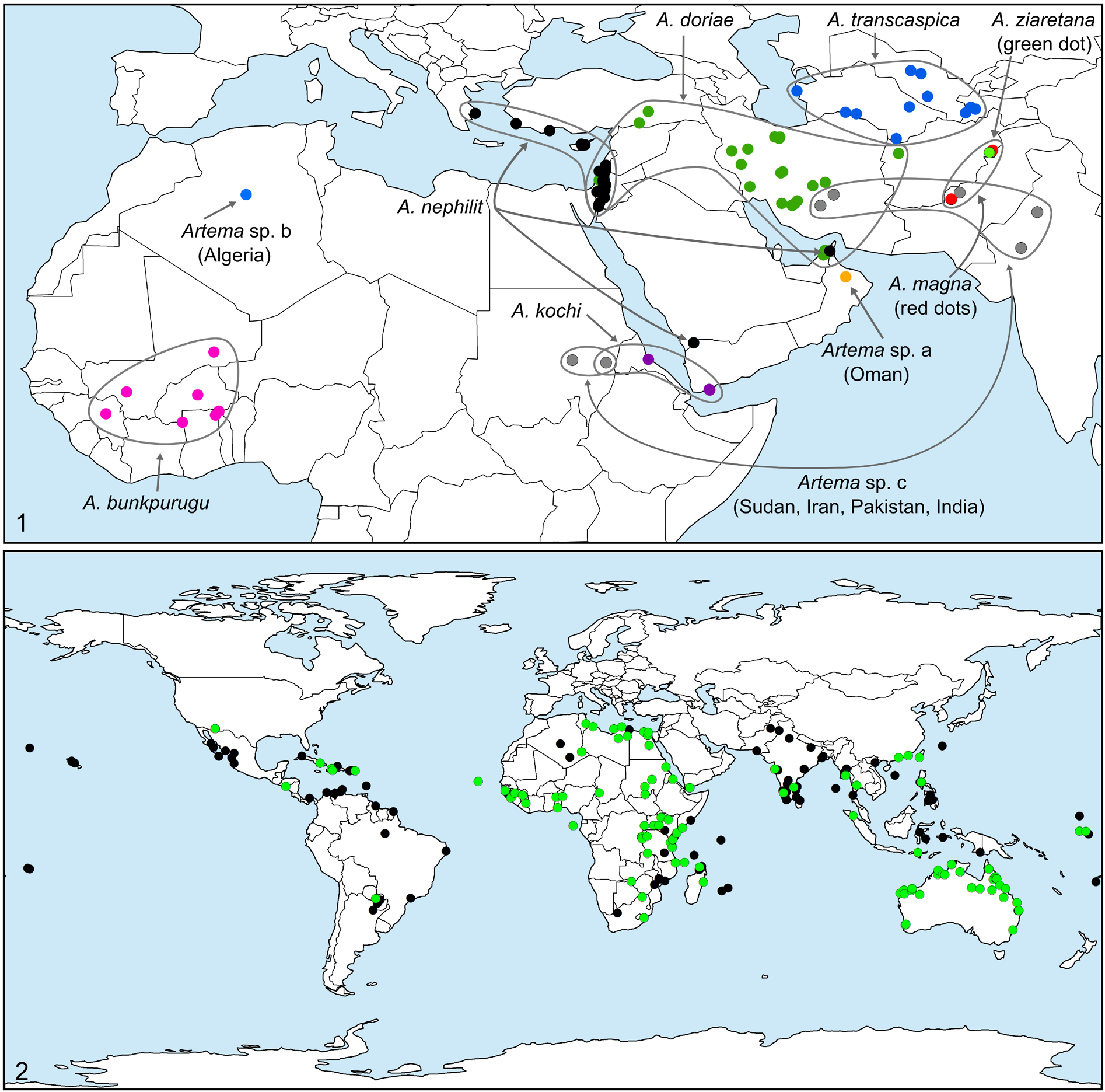
After the ubiquitous Holocnemus pluchei , A. nephilit sp. nov. is the most common pholcid in the Jordan Rift Valley. It has been collected from underground shelters and caves from the Eilat Mountains ( Fig. 8 View Figs 3–8 ) in the south, to Nahal Ayun in the north. Specimens from outside the Rift Valley ( Israel: Ramat Avedat near Sede Boqer; Oren Cave ( Fig. 7 View Figs 3–8 ), Mount Karmel; Jordan: Petra), as well as from Cyprus, Greece, and Turkey, indicate that the species is probably widely distributed in the area. Two females with very similar epigynum were collected from Yemen and the United Arab Emirates ( Fig. 1 View Figs 1–2. 1 ).
Notes
We have not seen the material identified by Bodenheimer (1937) as A. mauriciana . However, A. mauriciana (= A. atlanta ) does not seem to occur in Israel and there is no evidence that it ever existed there. We consider it likely that Bodenheimer was dealing with the common A. nephilit sp. nov. rather than with the much rarer (i.e., in Israel) A. doriae .
The females from Yemen (“Yar Ju Amlah” = Al Ja’milah?) and from the United Arab Emirates have a very similar epigynum, but since no males from these localities are known, they are assigned tentatively.







| ZFMK |
Germany, Bonn, Zoologische Forschungsinstitut und Museum "Alexander Koenig" |
| SMNH |
Canada, Saskatchewan, Regina, Royal Saskatchewan Museum |
| SMF |
Germany, Frankfurt-am-Main, Forschungsinstitut und Naturmuseum Senckenberg |
| NHMW |
Austria, Wien, Naturhistorisches Museum Wien |
| ZFMK |
Zoologisches Forschungsmuseum Alexander Koenig |
| SMNH |
Department of Paleozoology, Swedish Museum of Natural History |
| SMF |
Forschungsinstitut und Natur-Museum Senckenberg |
| MSNG |
Museo Civico di Storia Naturale di Genova 'Giacomo Doria' |
| NHMW |
Naturhistorisches Museum, Wien |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.