
Age: Middle Miocene, Astaracian
|
publication ID |
https://doi.org/ 10.1111/j.1096-3642.2007.00327.x |
|
persistent identifier |
https://treatment.plazi.org/id/55432810-A032-1E39-FF27-FD63FAC1538B |
|
treatment provided by |
Felipe |
|
scientific name |
Age: Middle Miocene, Astaracian |
| status |
|
Age: Middle Miocene, Astaracian View in CoL (MN 6).
Why a new lectotypification? Schaefer & Zapfe (1971) selected a lectotype for Anisodon grande among remains from Sansan. This specimen was a left maxilla (MNHN Sa 9340, formerly A.C. 4233) with P 4 –M 3. Unfortunately, this specimen is not part of the type series as it was not described by de Blainville (1849), so it could not be chosen as the lectotype (Article 74.1, ICZN, 1999: 82). Moreover, according to Article 74.2 (ICZN, 1999: 82), ‘If it is demonstrated that a specimen designated as a lectotype was not a syntype, it loses its status of lectotype’.
Schaefer & Zapfe (1971) stated that the left maxilla MNHN Sa 9340 was illustrated on plate 8 of the Ostéographie ( de Blainville, 1849). In fact, although there is a tooth row on plate 8, it is that of the right maxilla MNHN Sa 9339. This error is the result of two factors. Firstly, the illustrations published by de Blainville are reversed, as a result of the engraving techniques used at that time. Secondly, Schaefer & Zapfe (1971) were confused by the presence of the P 4 on the illustration of the tooth row ( Fig. 2B View Figure 2 ), as this tooth is present in MNHN Sa 9340 but not in MNHN Sa 9339. Conversely, the illustration of MNHN Sa 9339 is accurate in that the M 2 is unworn, and that the distal alveolus and the distal part of the mesial alveolus of P 3 are present (compare Figs 1 View Figure 1 and 2B View Figure 2 ). Hence, the P 4 was probably added to the illustration, maximizing the information provided. As there are other isolated P 4 s in the collection, we do not know whether de Blainville used the premolar of MNHN Sa 9340 when adding the missing P 4 to the illustration of the tooth row of MNHN Sa 9339.
Then, among the syntypes, the right maxilla MNHN Sa 9339 appears to be the best choice for the lectotype. It is well preserved and was illustrated by de Blainville (1849), thus following ICZN Recommendation 74B (ICZN, 1999: 83). Moreover, it clearly has some relation to Schaefer & Zapfe’s (1971) reasoning. In the following section, we provide a detailed description of the cranium (MNHN Sa 15670) found by Lartet and described briefly by de Blainville (1849). We also present the first description of the skull that belongs to the articulated skeleton found by Filhol (1890) at Sansan (MNHN Sa 15671).
Cranium MNHN Sa 15670
This specimen represents the first ever known chalicothere skull. It is obvious that Filhol (1890) examined the specimen, as he gave measurements missing from the original description by de Blainville (1849). Since that time, however, there has been no published evidence that any subsequent authors have studied this skull. De Bonis et al. (1995) were unable to locate it. Subsequently, this cranium was identified in the collection of the MNHN by one of us (PT), and Geraads et al. (2001) subsequently published a brief comment, but did not provide a detailed description. Furthermore, their comment is surprising with respect to the specimen (see discussion below). Consequently, a complete description of this specimen is provided with as much detail as possible.
Comparing the illustrations by de Blainville (1849: plate 8; see Fig. 2 View Figure 2 ) with the cranium today ( Fig. 3 View Figure 3 ), it is obvious that it was subsequently damaged, probably after 1890, as Filhol (1890: 296) provided measurements of the condyles, which are now lost. The two M 3 s were originally complete, but are now broken. De Blainville’s illustrations show the anterior part of the left zygomatic arch, which is absent today. Finally, there is a large vertical fracture on the posterior part of the cranium showing that the occipital region was dissociated from the remaining skull (the condyles were probably lost at that time).
Dorsal view: Sutures between the different bones are not apparent. The frontal crests are low, especially anteriorly. They converge posteriorly, but do not fuse to form a sagittal crest. Following this, the crests diverge to define the ‘interparietal triangle’ ( Barone, 1966; see also de Bonis et al., 1995), but because the superior part of the occiput is broken we cannot determine its orientation. The braincase is tubular.
Lateral view: The anterior part of the snout is broken, especially on the right side. On the left side, the orbital area is distorted: the inferior part of the maxilla and the anterior part of the orbit are separated and joined to the remainder of the skull in a more posterior and lower position than would have occurred in life. This probably explains de Blainville’s (1849: 68) error when he described the orbit as ‘very small, round and quite low’ (our translation from the French original). Indeed, the anterodorsal margin of the orbit is present on the skull roof clearly anterior to its suggested position.
The foramen infraorbitale lies above the distal bor- der of M1. The alveolar border is convex in lateral view. There is an outgrowth of the maxilla just above the mesial root of M2. Such an outgrowth is unknown in other chalicotheriines. Sutures between the maxilla and jugal, and between the maxilla and lacrimal, are marked by a shallow vertical groove. The jugal extends anteriorly to a point situated above the parastyle of M2. The orbitosphenoidal area is badly damaged, but it is still possible to observe the deep groove accommodating the optic nerve on the orbit floor. The postglenoid process is strong and flattened anteroposteriorly. It is separated from the external auditory meatus by a narrow furrow. Above the meatus, there is a strong tuber that develops from the lambdoid crest. The meatus is bordered posteriorly by the posttympanic process of the squamosal. The ventral margin of the meatus displays a blunt, rounded process. The squamosal is slightly domed, so the braincase is swollen in transverse section. The occipital face is vertical.
Occipital view: Superficially, the occipital face seems to be low, but this results from the lack of its lower part (including the condyles, foramen magnum, and paroccipital processes). Its upper edge is rounded. The occipital tubercle is high on the occiput and continues ventrally as a vertical ridge. The paroccipital process continues dorsally on the occipital face as a vertical ridge.
Ventral view: The skull is strongly distorted and the anterior part is translated laterally. Moreover, the anteroposterior axis of the basicranium is not exactly the same as that of the snout. A large part of the guttural area is missing.
The snout is broken in front of the P 3, but the distal end of the left P 2 alveolus is preserved. The tooth rows converge strongly anteriorly. The zygomatic arch extends as far forward as the distal part of M 2. The anterior part of the ventral edge of the zygomatic arch displays a strong elongate tuberosity. Between this tuberosity and M 3 there is a deep anteroposterior groove. There is no retromolar space. The palate is broken displaying numerous fractures, and the choanal opening is missing.
What is preserved of the guttural fossa shows that it was divided sagittally by a high crest that issued from the basisphenoid, anterior to the basioccipital crest. The postglenoid processes are anteroposteriorly flattened and their anterior faces display a subvertical, ovoid articular facet. The tympanic bulla is elbowshaped and flattened ventrally. On both sides, the anterior borders of the bullae are broken, but it is obvious that they extended more anteriorly than the postglenoid processes. The auditory duct is pressed against the posterior face of the postglenoid process. The vaginal crest and stylohyoid fossa are only partially visible, but it is still obvious that they were positioned as far posteriorly as the external auditory meatus. Although the paroccipital processes are broken at their base, it is clear that they were orientated ventrally and externally. The foramen ovale opens in front of the anterior wall of the bulla. The foramen lacerum medium cannot be located. Anterior to the foramen ovale, there is no trace of a canalis alisphenoideus. Although its absence cannot be confirmed in MNHN Sa 15670 because the sphenoid area is badly damaged, specimen MNHN Sa 9398 (see below) displays no canalis alisphenoideus. The foramen caroticum perforates the bulla on its posterior ventral margin, whereas the foramen metoticum and canalis nervi hypoglossi cannot be observed.
Teeth are badly preserved on this specimen, and yield no useful new information. What is visible does not differ from the other remains from Sansan.
Skull of the complete individual discovered by Filhol (MNHN Sa 15671)
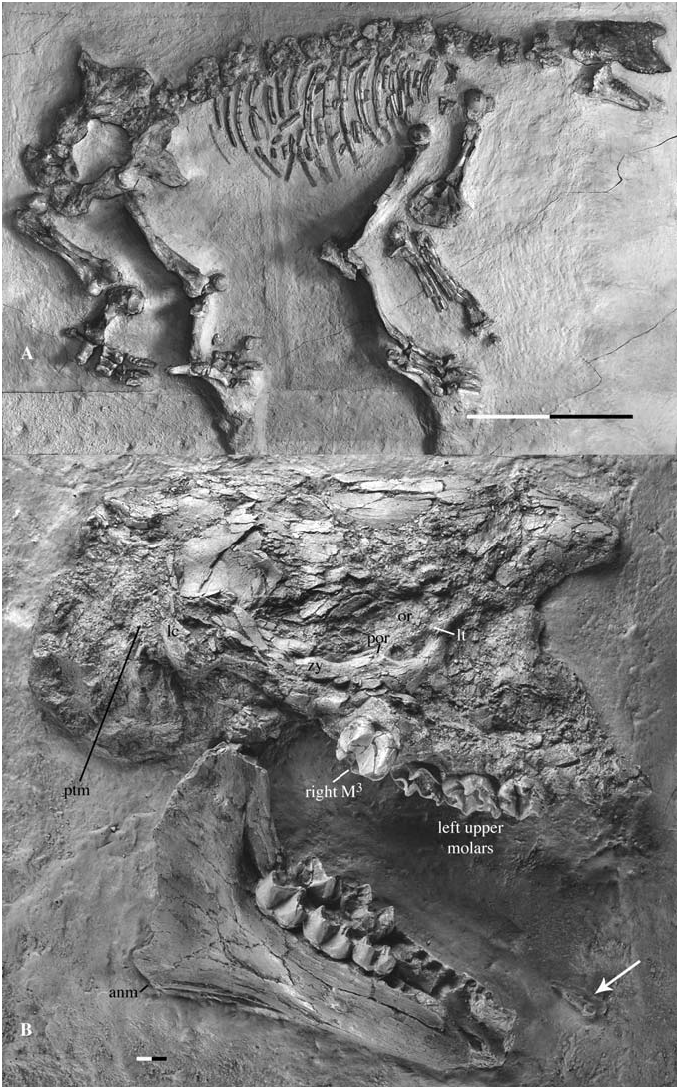
When Filhol (1890) announced the discovery of a complete, articulated skeleton, he emphasized the association between a ‘ Macrotherium ’ postcranium (which was thought to be an ‘edentate’ at this time) and a skull with a Chalicotherium - type dentition, but he did not describe the cranial anatomy. In fact, nobody has done this to date. This omission may be because this skull is quite crushed and fragmented, and taking its distortion into account is therefore difficult. The specimen ( Fig. 4A View Figure 4 ) lies on its left side and is embedded in a large slab of lacustrine limestone from the lower levels of the Sansan deposits ( Filhol, 1890). A cast of the cranium was made to facilitate study. Although badly crushed, this skull yields a number of important observations.
Cranium: Viewed as a whole ( Fig. 4B View Figure 4 ), the cranium seems very high throughout its length. The upper profile of the skull roof seems to be horizontal and the occiput is exceptionally bulged posteriorly and ventrally. Finally, the lengthening of the skull appears to be too great for what is traditionally considered as a ‘macrothere’. However, these first impressions do not reflect the original morphology.
In fact, the degree of distortion misrepresents the original morphology and only a few features can be confidently identified on this specimen. Anteriorly, the two maxillae are closely pressed against one another. The right maxilla is represented only by the M 3 and a part of its associated alveolar margin. The rest of the right maxilla consists of small bony fragments lying inside the nasal cavity. The left maxilla is better preserved: the three molars are present and it is possible to follow the alveolar margin up to the alveolus of P 2. The anterior border of the left maxilla seems to indicate the position of the nasal opening, but it is difficult to be sure whether it is damaged or not, so the shape of the nares remains conjectural. Dorsally, fragments of the nasal(s) and frontal(s) are present. This area is so badly crushed and distorted that it cannot be concluded that the nasals are as far anteriorly extended as they appear to be, in a way that somehow resembles extant tapirs. Posteriorly, the skull roof is better preserved. However, deformation of the originally horizontal skull roof into a two-dimensional, vertical plane gives the misleading impression that the skull is very deep. There is no sagittal crest. The lower part of the lambdoid crest can be observed as can the posttympanic process of the squamosal, which lies slightly posterior to the lambdoid crest. The preserved part of the lambdoid crest indicates that the occiput did not project posteriorly. All of the occiput, and what is visible of the basicranium in lateral view, is crushed, giving the misleading impression that the skull bulges posteriorly and ventrally.
The principal features of interest in this skull are the preservation of the zygomatic arch and anterior border of the orbit. Despite its bad condition, this specimen is the only one from Sansan that displays these areas. The rounded anterior border of the orbit shows a strong lacrimal tubercle. The zygomatic arch is shattered, but the remaining fragments are easily distinguishable. The jugal bears a moderate, but obvious, postorbital process just anterior to the jugalsquamosal suture. The zygomatic arch is slightly bowed posterodorsally, but in any case is not as high as the superior border of the orbit.
Mandible: The two hemimandibles are pressed against one another ( Fig. 4B View Figure 4 ). Only one observation can be made concerning the symphysis: on the left hemimandible, a dorsal part of the symphysis is present and extends posteriorly at least up to the P 2, but the right hemimandible hides its complete extent. The corpus increases in height posteriorly. Its ventral border is straight. The angulus mandibulae expands ventrally. In lateral view, the tooth rows are slightly concave. The slope of the anterior border of the ramus is orientated at about 90 degrees relative to the corpus. There is no retromolar space. The coronoid process is partly broken and the crest present on its anterolabial border is low.
Upper dentition: Only the left M 1 –M 3 and right M 3 are present. This specimen was probably a young adult when it died as tooth wear is minimal. On each molar the protoloph only reaches the paraconule, and continues as a low crest until the protocone summit. The protocone is receded but its position is still relatively anterior, leaving a large, U-shaped distal valley behind it. A vertical crest descends from the summit on both the labial and lingual walls of the paracone. The mesial cingulum is strong and not reduced in front of the protocone. On M 1 and M 2 the metacone is more labial than the paracone.The metaloph is shorter on M 3 than on M 2 (because of the contraction of the distal part of M 3). On M 3 the metacone is labially displaced. The external wall of the metacone and metastyle is nearly parallel to the mesiodistal length. The mesiolingual face of the hypocone of M 3 is very abrupt and the distal cingular crest does not reach the summit of this cusp. Finally, the postfossette (sensu Butler, 1965) is not more pinched on M 3 than on the two other molars (see below). The lingual cingulum is weakly developed.
Lower dentition: Right M 1 –M 3 are undamaged, whereas left M 2 –M 3 are broken. The left hemimandible displays the alveoli of P 2 –M 1, whereas the right is broken anterior to the alveoli of P 4. On the molars, the paraconid is low and the metaconid is the highest cusp. The ‘metastylid’ [perissodactyls lost the metastylid early in their evolution, this pseudometastylid is the distal cusp of a twinned metaconid ( Hooker, 1994)] is observable, although it is not particularly distinct from the metaconid. It shows a well-developed metacristid. The talonid is slightly more elongated than the trigonid and both are V-shaped. The mesial wall of the hypolophid and the distal walls of the protolophid and entolophid show a weak vertical, medial incisure (see description of MHNT VAL-3, below). The cingulum is thickened on the distal face and weak elsewhere.
The other teeth are represented by their alveoli, and it is notable that P 2 has two alveoli. An isolated tooth ( Fig. 4B View Figure 4 ) is embedded in the sediment several centimetres in front of the mandible. This tooth is complete and has only one root. It is either a lower canine or a lower incisor. Unfortunately, little is known of such teeth in Anisodon grande . Zapfe (1979) tentatively assigned some isolated teeth from Neudorf (MN 6, Slovak Republic) to incisors of A. grande , but in the absence of any association. These teeth are slightly flattened labiolingually. The labial face is flat, whereas the lingual one shows two enamel crests descending toward the lateral border of the tooth and joining a basal, lingual point. They are either sharp or rounded, probably as a result of differential attrition by the tongue and food. The incisors of the holotype of Anisodon macedonicus (UT DKO 234) are much reduced, rounded and button-shaped ( de Bonis et al., 1995). The isolated tooth associated with MNHN Sa 15671 does not correspond to any of these descriptions, and is more compatible with lower canine morphology: a triangular crown with a distal crest showing a weak lingual, vertical groove (see description of the symphysis MHNT VAL-4 from Saint-Gaudens, below). For the moment, this tooth is provisionally identified as a right lower canine.
Additional observations
Other cranial remains from Sansan (Appendix 4) provide some additional character information that is not seen in the two skulls described above.
MNHN Sa 9398 includes several cranial fragments that belong to the cerebral region. The occipital condyle is relatively small, oval, low, and narrow. The ‘interparietal triangle’ is displaced on the dorsal face of the skull. A fragment of the left basicranium confirms the absence of the canalis alisphenoideus.
The crowns of P 2 and P 3 are unknown at Sansan. On P 4, the protoloph does not reach the protocone and ends at the paraconule. On very worn teeth it only reaches the base of the protocone. On molars, the crest extending the protoloph continues until either the protocone summit or the paraconule base (intraspecific variation). The development of the crest on the lingual wall of the paracone is also variable. Sometimes, the distal part of M 3 is less reduced and the metacone is less displaced labially (e.g. MNHN Sa 9373).
The symphysis is never completely preserved in any specimen found at Sansan, but several remains suggest that it extends posteriorly up to the level of the P 2 –P 3 boundary. There is no retromolar space on the mandible. The angle between the anterior border of the ramus and the corpus is variable. When it is preserved, the angulus area always expands ventrally.
The P 2 is similar to that of UT DKO 234. Only one complete P 4 is known from Sansan. It is molar-shaped, but the talonid is lower than the trigonid. The entoconid is weak.
The awkward symphysis described by de Blainville
De Blainville (1849: 68–69, plates 8 and 9) described and illustrated the right hemimandible MNHN Sa 9376. According to him, three incisors were present in a rather long symphysis that expands laterally forming a palette-shaped structure ( Fig. 2A View Figure 2 ). There is no doubt that de Blainville illustrated the specimen MNHN Sa 9376 – which, nevertheless, now lacks the symphysis. With the exception of the symphysis, the outline of the specimen is exactly the same and P 2 is drawn in its conspicuous pathological position (note that only M 2 and M 3 are present in MNHN Sa 9376, but de Blainville completed the dentition on plate 8 probably for the needs of illustration as he did for MNHN Sa 9339). The symphysis illustrated by de Blainville is somewhat reminiscent of that of a hippopotamus, because of its great dilatation. Yet, neither MNHN Sa 9376 nor any other remains from Sansan [nor mandibles from Neudorf described by Zapfe (1979)] display evidence for such a morphology. From his description, it is obvious that de Blainville observed a symphysis. Was he misled in his identification, assembling two isolated pieces erroneously, or has this material been misinterpreted by de Blainville and lost since? We tentatively interpret this specimen somewhat differently. Comparison with other known chalicotheriine symphyses (especially MHNT VAL-4 from Saint-Gaudens, see below) reveals that the original remains observed by de Blainville would have been distorted. The anteroposterior axis of the symphysis seems to sit at an angle of 20–30° to that of the corpus. If this interpretation is correct, the symphysis would not be more expanded than in other chalicotheres. More importantly, if this symphysis is genuine, there would have been two incisors instead of three as de Blainville thought (see below). Such a hypothesis can only be confirmed by study of this symphysis, but no such specimen could be located in the MNHN collection.
Note concerning remains from Neudorf
Zapfe (1979) described chalicotheriine remains from Neudorf an der March, now Devínska Nová Ves ( Slovak Republic), and assigned them to Chalicotherium grande . These remains were found in fissure fills referred to the Middle Miocene (MN 6). The chalicotheriines from Neudorf are represented by at least 60 individuals, including both cranial and postcranial remains ( Zapfe, 1979).
Geraads et al. (2001: 600) raised doubts concerning the conspecific status of the Neudorf and Sansan remains. Our study confirms this suspicion as some specimens from Neudorf differ from their equivalents at Sansan. This means that perhaps two taxa are associated at Neudorf (probably Anisodon grande and a Chalicotherium ). The Neudorf remains need to be reviewed to establish their taxonomic status.


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |