
Ciconia nana ( De Vis, 1888 )
|
publication ID |
https://doi.org/ 10.3853/j.0067-1975.57.2005.1440 |
|
persistent identifier |
https://treatment.plazi.org/id/585C1A03-6419-FFDF-11D4-CFEA5281FD79 |
|
treatment provided by |
Felipe |
|
scientific name |
Ciconia nana ( De Vis, 1888 ) |
| status |
|
Fig. 1 View Fig
Xenorhynchus nanus De Vis, 1888 . Proc. Linn. Soc. N.S.W. 3: 1287, Qld: Darling Downs: Condamine River: Chinchilla.
Ciconia nana ( De Vis, 1888) . Rich & van Tets, 1982: 306A; van Tets, 1984: 470; van Tets & Rich, 1990: 166; Vickers-Rich, 1991: 752.
De Vis (1888) based Xenorhynchus nanus on material from the Condamine River, near Chinchilla, in the Darling Downs of Queensland. The material comprised a distal tibiotarsus, collected by J. Daniels, and a proximal ulna, a later acquisition but described at the same time. Subsequently De Vis (1905) reported this species from Wurdulumankula, Cooper Creek, South Australia, based on a distal tibiotarsus, collected by Professor J. Gregory. The original tibiotarsal fragment was designated as the lectotype by Brodkorb (1963). Similarities in size and shape to species of Ciconia View in CoL were noted by Rich & van Tets (1982), who provisionally transferred this form to this genus, where it has been listed in subsequent reviews (e.g., van Tets, 1984; van Tets & Rich, 1990; Vickers-Rich, 1991), although no detailed comparisons had been made.
The Darling Downs, southeastern Queensland, feature deposits of two discrete periods. Pleistocene deposits occur on the east side of the Condamine River (26°48'S 150°41'E), producing the Darling Downs Local Fauna (Molnar & Kurz, 1997). The older, Pliocene-aged assemblage, the Chinchilla Local Fauna, which yielded the holotype of X. nanus , comes from the fluviatile Chinchilla Sands along the western banks of the Condamine River , near Chinchilla. On the basis of closer faunal resemblances of this fauna to the Early Pliocene Bluff Downs Local Fauna than to the Pleistocene Darling Downs Local Fauna, its age has been put at Early to Middle Pliocene ( T. Rich et al., 1991) GoogleMaps .
Many important specimens from Gregory’s trip along Cooper Creek have their locality listed as Wurdulumankula, although no similar place name has been found on Gregory’s maps ( Gregory, 1906), and the exact location of this site is uncertain (Tedford & Wells, 1990). It is considered to be located in the Piranna Soakage of Cooper Creek, in the eastern Lake Eyre basin, South Australia, and to be one of a number of sites from which fossils of the Malkuni Fauna have been recovered, one of two faunas in the fluviatile deposits of the Katipiri Formation (Tedford & Wells, 1990). These assemblages represent fluviatile/lacustrine faunas in the Great Artesian basin that predate the formation of inland dunes and are probably Late Pleistocene in age ( Woodburne et al., 1985). The fossils are found as “float” or in place.
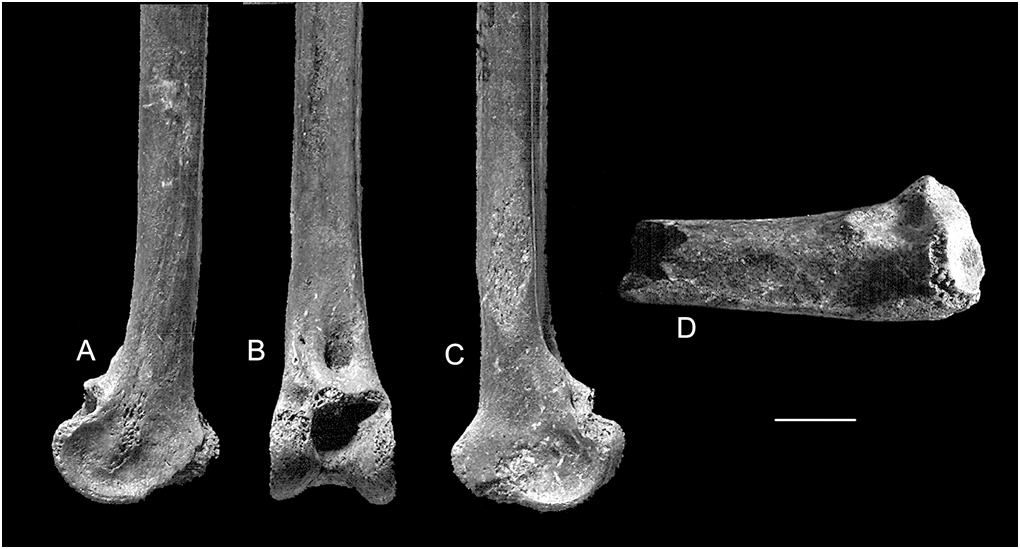
Lectotype. Distal tibiotarsus ( QM F1131 ; Fig. 1a–c View Fig ; Brodkorb, 1963).
Type locality. North bank of the Condamine River , 5 km from Chinchilla, Darling Downs, Queensland .
Paralectotype. Proximal ulna ( QM F5514 , Fig. 1d View Fig ) .
Etymology. Nanus (Latin, a dwarf), in reference to the small size of this bird in comparison with its putative congener, E. (X.) asiaticus .
Diagnosis. Ciconia nana is diagnosed from other living species in the genus by the following combination of characters: the anterior extension of the condylus lateralis relative to the condylus medialis is greater; the ridge from the papilla for M. tibialis cranialis to the condylus medialis is only slightly incised by a furrow; and the tuberculum retinaculi m. fibularis is nearly confluent with the lateral border of the shaft (in anterior view).
Of the fossil forms, few can be compared with C. nana for lack of common elements. Ciconia maltha was much larger ( Table 1), as was the unnamed Mio-Pliocene species from Arizona, which Bickart (1990) characterized as a “giant, equalling in size large individuals of the extant Jabiru mycteria ”. The somewhat younger Ciconia stehlini had a tibiotarsus of comparable proximal width to C. nana . Jánossy (1992) did not provide any characters that serve to separate these species morphologically. Of living species, C. ciconia and C. nigra are similar to C. nana in proximal width of the tibiotarsus but the condyli of C. nana are deeper.
Referred material. Distal tibiotarsus (QM F5513), Wurdulumankula, Cooper Creek, South Australia.
Measurements. Table 1.
Description
De Vis’ (1888) description of the original tibiotarsus (QM F1131) was detailed, and identified diagnostic characters at family, generic and specific levels. Descriptions of the other specimens amounted to just a few adjectives. The second tibiotarsus “adds nothing to our information about the smaller Jabiru than that it attained a rather larger size than the tibia already described”. Its distal width was greater and “all parts of the bone are proportionately larger” ( De Vis, 1905). About the ulna De Vis (1888) remarked only that it was of compatible size with the first tibiotarsus but “unfortunately its worn condition unfits it for description”.
Tibiotarsus. The lectotypical tibiotarsal fragment QM F1131 consists of the distal end with the shaft broken distal to the crista fibularis (length 118.6 mm as preserved). It is abraded on the proximal borders of both condyli, the cristae of trochlea cartilaginis tibialis, the epicondylus medialis and the papilla for M. tibialis cranialis. The anterior face of the shaft is flattened, with the linea extensorius developed into a low ridge along its distal end. The posterior face is strongly rounded. The tuberculum retinaculi m. fibularis is confluent with the lateral border of the anterior face of the shaft; a nutrient foramen is proximal to this. There is a large, broad, elongate scar on the lateral face of the shaft; the distal end is level with the tuberculum. The tuberositas retinaculi extensorius on the linea extensorius is small but obviously elongate. The sulcus extensorius is of uniform depth, not deepening markedly proximal to the pons supratendineus. The distal border of the pons supratendineus is developed anteriorly into a strong ridge. The condyli are similar in shape and size, with the condylus medialis extending slightly further distally and more markedly anteriorly. The depressio epicondylus lateralis is deeper and more extensive than the depressio epicondylus medialis; both have prominent rims anteriorly. Despite abrasion, the cristae of the trochlea cartilaginis tibialis are prominent, extending well away from the shaft. Measurements, Table 1.
The other tibiotarsal fragment, QM F5513 , is slightly larger and has less abrasion of the distal end. It retains about half the length of the shaft, as does the lectotype (length 62.6 mm as preserved) and the medial half is missing for much of this. It agrees closely with the lectotype in morphology other than that the sulcus extensorius is slightly deeper, the tuberositas retinaculi extensorius is more raised, the tuberculum retinaculi m. fibularis is a small distance from the lateral border rather than confluent with it, and the distal opening of the canalis extensorius is somewhat larger and rounder .
Ulna. Specimen QM F5514 consists of the proximal end of a right ulna. It is rather damaged, with the olecranon missing, and moderate to heavy abrasion on the cotyla dorsalis, facies articularis radialis and crista intercotylaris. It is broken distal to the proximalmost papilla. The impressio m. brachialis is long, moderately deep proximally and shallow distally. It is bounded posteriorly by a broad, rounded tuberculum lig. collateralis ventralis and anteriorly by a heavy ridge, which separates it from a prominent incisura radialis. Although, because of the abrasion, measurements of this element cannot be compared directly with those of other taxa, overall the specimen is slightly smaller than the ulna of Ciconia ciconia . The measurements of the specimen as preserved are, length 46.0 mm; proximal width 16.6 mm; proximal depth 11.8 mm.
De Vis (1888) placed this species in the same genus as the living Ephippiorhynchus asiaticus , “noting further its strong resemblance to the Jabiru’s tibia in the massiveness, direction, and sculpture of the bridge traversing the intercondylar space, we cannot but admit congeneric affinity between the two”. A comparison of the distal tibiotarsal fragments with other living species of the Ciconiidae , and E. asiaticus in particular, demonstrates that De Vis’ generic allocation for these specimens is not supported. The comparative material available to De Vis was limited to selected taxa, almost all of Australian origin. Thus, his placement of the fossil specimens in the same genus as the only Australian species is not surprising; it is doubtful that he had access to osteological representatives of any other genera of storks.
De Vis (1888) noted size differences between the fossil tibiotarsus and that of E. asiaticus : “the fossil tibia … is in the mean two-ninths less in its dimensions than the recent bone, indicating a bird but little more than half the bulk of the jabiru of the present day”. The morphological differences he mentioned were that “the rotular channel is shallower; there is considerably less intercondylar space behind the posterior edge of the bridge, the canal under the bridge is relatively much wider, the ectocondylar tubercle is not prominent, and the double flexure inwards and forwards apparent in the living jabiru between the shaft and the articular end is scarcely appreciable”. Most of these are either actually differences between Ciconia and Ephippiorhynchus or have no generic significance.
The condition of the ulnar fragment is not suitable to permit a useful comparison. The shallow, round impressio m. scapulotricipitalis may be of generic significance but it is also likely that the possible slight morphological differences between the fossil and recent specimens are due to abrasion.
Riversleigh stork
The presence of a new species of stork from Oligo-Miocene deposits at Riversleigh, was briefly mentioned by Vickers- Rich (1991). Boles (1991) noted that it “was not close to the living … Xenorhynchus ” without further elaboration, and subsequently ( Boles, 1997) stated that the material was “probably referable to Ciconia ”. This taxon is described below.
The Riversleigh deposits are located 5 km west of the Riversleigh homestead (19°02'S 138°45'E), 200 km north of Mt Isa , northwestern Queensland, where they occur as an outcrop of Tertiary limestone overlying the Cambrian Thorntonia Limestone. There are now over 200 named Oligo-Miocene deposits at Riversleigh. An informal system of grouping has been used (Systems A–C). These systems are “regionally clustered sites that appear to be superpositionally-related (differing in age but not significantly in position) and/or space-related (spatially isolated but approximately contemporaneous)” ( Archer et al., 1989). The principal accumulations are thought to have occurred in several episodes involving large lakes, shallow pools and cave deposits. Undoubted stork material has been recovered from three sites and a referred specimen comes from a fourth GoogleMaps .
White Hunter Site, Hal’s Hill Sequence, D-Site Plateau, is considered to be part of System A, of Late Oligocene/ Early Miocene age ( Creaser, 1997; Myers & Archer, 1997). The White Hunter Local Fauna also contains other birds, including the small casuariid Emuarius gidju (Patterson & Rich, 1987) (Boles, 1992) , the dromornithid Barawertoris tedfordi Rich, 1979, a flightless rail ( Boles, 2005) and several passerines. Wayne’s Wok Site is in the central section of the D-Site Plateau. Its age is still unclear, but may be System A or B (?Early to Middle Miocene) ( Black, 1997; Cooke, 1997a; Creaser, 1997). Birds found here also include Emuarius , dromornithids and passerines. Bitesantennary Site is a cave deposit in the Verdon Creek Sequence, on the northern section of the D-Site Plateau, where it intrudes into the widespread D-Site layer. It is possibly a System B site ( Cooke, 1997a).
Specimens of five skeletal elements were obtained at Bitesanntennary Site in close proximity and are assumed to have been associated. Proximal tarsometatarsal fragments from White Hunter Site and Wayne’s Wok Site allow direct comparison with each other but not with a distal tarsometatarsal fragment from Bitesantennary Site. Because the tarsometatarsal fragments all come from storks of comparable size and morphology, they are referred to the same taxon. A cervical vertebra from Neville’s Garden Site (Early Miocene) is tentatively referred to this species because of its size and morphological similarity to that of living storks.
| T |
Tavera, Department of Geology and Geophysics |
| QM |
Queensland Museum |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Ciconia nana ( De Vis, 1888 )
| Boles, Walter E. 2005 |
Ciconia nana ( De Vis, 1888 )
| Vickers-Rich, P 1991: 752 |
| van Tets, G 1984: 470 |