
Ciconia louisebolesae, Boles, 2005
|
publication ID |
https://doi.org/ 10.3853/j.0067-1975.57.2005.1440 |
|
persistent identifier |
https://treatment.plazi.org/id/585C1A03-641C-FFD2-1199-CE765467FD21 |
|
treatment provided by |
Felipe |
|
scientific name |
Ciconia louisebolesae |
| status |
sp. nov. |
Ciconia louisebolesae n.sp.
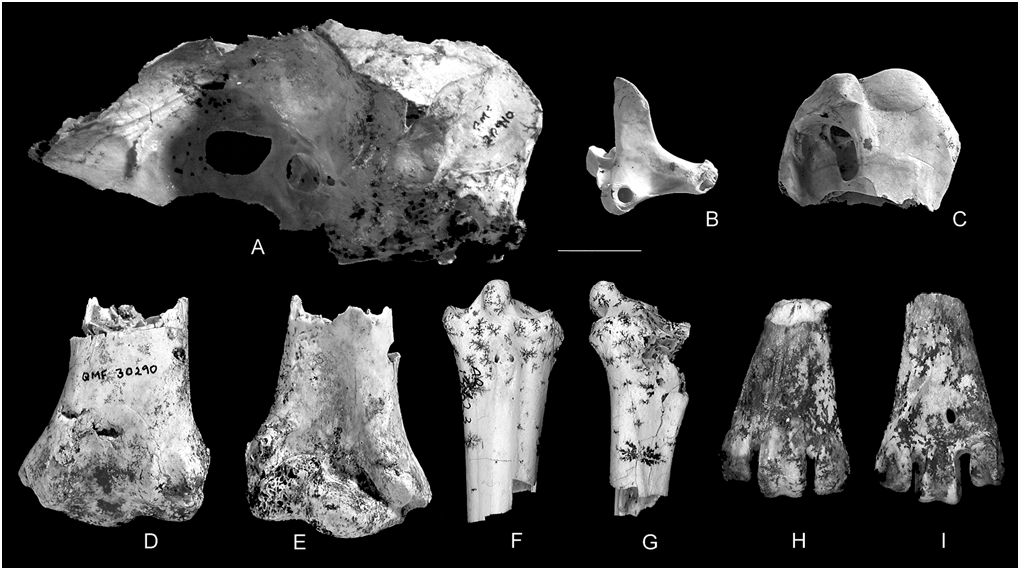
Fig. 2 View Fig
Holotype. QM F30290, right distal humeral fragment with surface damage to the anterior face of the condylus dorsalis, tuberculum supracondylare ventrale and dorsal border of sulcus humerotricipitalis.
Type locality. Bitesantennary Site, Riversleigh, northwestern Queensland, currently considered to be Early Miocene; Bitesantennary Local Fauna.
Paratypes. All from Bitesantennary Site. Cranium — QM F20910, neurocranium, lacking skull roof; quadrate — QM F20893, complete right element; humerus — QM F20911, proximal right element broken through distal to the midpoint of the crista bicipitalis and to the fossa pneumotricipitalis, and missing the tuberculum dorsalis and processus deltopectoralis; tibiotarsus — QM F31350, extreme proximal left element broken through the shaft through proximal end of the foramen interosseum proximale; damage to most projecting features, including both the cristae cnemialis, particularly the crista cnemialis caudalis, the medial edge of the facies articularis medialis, and extensively on the posterior edge along the area of contact between the facies articularis medialis and lateralis; Tarsometatarsus — QM F36446, right distal fragment broken through the shaft proximal to the fossa metatarsi I .
Etymology. Dedicated with love and respect to my mother, Louise Boles, for her guidance in my development as a person and her tolerance of my many transgressions.
Diagnosis. Similar in size to C. ciconia and C. nigra , but sufficiently different to recognize as a new species. It differs by the following suite of characters of the distal humerus: the condylus ventralis humeri is proportionally smaller, not extending as far ventrally; the epicondylus ventralis is closer to the distal end; the muscle scars distal to the epicondylus ventralis are more extensive (in ventral view) and more excavated (in anterior view), although less than in genera such as Ephippiorhynchus ; and the posterodorsal corner is more rounded (in dorsal view). The crista bicipitalis is not short (as in C. maguari ) and is prominently extended. In its size and comparative narrowness of the incisura tibialis, the tibiotarsus more closely resembles that of the small species of Ciconia (e.g., episcopus and abdimii ) rather than C. ciconia or the larger taxa.
Referred material. QM F50428 View Materials (Neville’s Garden Site) cervical vertebra with only minor damage; QM F36445 (Wayne’s Wok Site) left proximal tarsometatarsus with damage to the crista hypotarsi medialis and plantar face of cotyla lateralis; QM F36447 (White Hunter Site) proximal left tarsometatarsus missing most of hypotarsus.
Measurements. Tables 2–4.
Description
Cranium. Measurements as preserved: 31.7 mm width of parietal; 40.1 mm width at midline of orbits (front of specimen as preserved); 40.7 mm width at processes temporalis. The specimen comprises the rear of the skull missing the dome of skull dorsal to the fossa temporalis and in a line just across top of the orbitae, retaining most of the parietal and frontal on a level with the orbital rims; the lamina parasphenoidalis is present but palatal elements are missing; the interorbital septum is present to in front of the fontaculi interorbitales and about half way through the orbits. Many of the external structures are damaged, with the processus postorbitalis lost and the posteroventral border of the processus paraoccipitalis and edges of the lamina parasphenoidalis slightly to strongly abraded; a large opening in the centre of the septum interosseus is a post- mortem artefact. The orbitae are broad and rounded. The fossa temporalis is broad, with the crista temporalis nuchalis strongly developed. The processus zygomaticus is short and blunt. The foramen magnum is squarish oval, with the dorsal border broken. The condylus occipitalis is hemispherical, with the dorsal border abraded. The recessus tympanicus dorsalis is moderately small and round; the fenestra vestibule, foramen pneumaticum caudale and fenestra cochleae are tightly grouped in a small recess. The cotylae quadratica otici and squamosi are of similar circular shape and size. The interior of the cranial cavity is largely intact. The fossa cranii caudalis and fossa tecti mesencephale are broad and circular, with the crista tentorialis prominent.
Quadrate. Viewed anterodorsally, the element is very transversely expanded across the processus mandibularis and less so across the processus oticus; the midbody is comparatively thin between these processes, with the lateral side markedly concave. The crista tympanicum is low. The processus orbitalis is straight on its anterior border; its posterior border is straight until dorsal end, which curves to meet the anterior border at the apex. The capitula squamosum and oticum are elliptical, their long axes converging anteriorly (in posterodorsal view). The incisura intercapitularis is broad but shallow; it extends to a large round foramen pneumaticum located centrally on the posterodorsal surface. The region from the base of the capitulum squamosum, through this foramen, diagonally towards the base of the condylus lateralis is anteroposteriorly compressed compared to the rest of the element. The posterior surface between the processus mandibularis and processus oticus is very slightly concave (in lateral view). The condylus medialis is the most medially projecting part of the element, while the projection of the condylus lateralis is the greatest laterally, exceeding that of the processus oticus. The condyli lateralis/caudalis lie perpendicular to the processus orbitalis. The small, oval condylus pterygoideus stands discrete from the condylus medialis, separated by a small sulcus.
Cervical vertebra. This is very elongate (greatest length 39.7 mm, proximal width 26.2 mm, proximal depth 19.8 mm, distal width 18.6 mm, distal depth 19.5) and agrees with modern Ciconia ciconia in morphology. It is not diagnosed further.
Humerus. In anterior view, the caput humeri is rounded, sloping dorsally from the apex more steeply than on the ventral side; it is moderately broad, becoming expanded at its anteroposterior midpoint. Its distal border on the posterior surface is obscure. The sulcus lig. transversus is broad and deep but short, confined to the ventral third of the anterior face, barely reaching the ventral border of the caput; it ends abruptly at the edge of the anterior face of the element. Its depth is augmented by a bordering ridge on its proximal side and the intumescentia humeri on the distal. The proximal surface of the intumescentia is flat and smooth and (in proximal view) is little inflated; it is tilted dorsally to join smoothly with the shallow, indistinct impressio coracobrachialis.
On the posterior surface, the broad incisura capitis separates the caput humeri from a long, deep and thickened area along the proximoventral border of the element, proximal to the fossa pneumotricipitalis, before merging with the proximal end of the posterior shaft surface without an obvious demarcation. Ventrally the prominent proximal section attenuates towards the crista bicipitalis, but protrudes substantially more posteriorly than the crista. On its ventral end, it supports the elongate scar for M. biceps brachii.
The tuberculum ventrale is relatively large, triangulartrapezoidal in shape and oriented proximodorsally. In proximal view, it sits at the apex of a triangular block bounded along its dorsal base by the sulcus lig. transversus. This triangular block protrudes posteriorly well beyond the rest of the proximal end of the element. The deep but moderately thin crus dorsale fossae runs distally from this, forming the dorsal border of the fossa pneumotricipitalis. The fossa is elliptical and highly pneumatic. The area between it and the crus ventrale fossae is excavated as a broad concave basin. The thick crus has a well marked, elongate scar for the attachment of M. scapulohumeralis caudalis. The crista bicipitalis is not well-developed ventrally.
The ridges bordering the sulcus scapulotricipitalis are short but moderately pronounced; they do not extend to the distal border. The ridge on the ventral side of the sulcus humerotricipitalis is broader and longer but about the same height as those defining the sulcus scapulotricipitalis. The sulcus humerotricipitalis is broad and shallow, with no obvious separation from the flat posterior surface of the shaft proximal to it; it is apparent only because of the bordering ridges. The fossa m. brachialis becomes confluent with the shaft on its dorsal and proximal sides and is poorly demarcated on its ventral side. The ridge along the ventral border supporting the tuberculum supracondylare ventrale is strongly developed and rather broad. In distal view, the condylus dorsalis humeri extends further anteriorly than does this ridge, which is turn projects only slightly more than the condylus ventralis humeri. The distal border of the condylus dorsalis humeri just reaches the dorsodistal corner. The condylus ventralis humeri is a rounded oval without obvious inflation. The incisura intercondylaris is moderately broad but shallow. The scars for M. flexor carpi ulnaris and M. pronator profundus face distally; they are adjacent, separated by only a thin ridge. The scars for M. ectepicondylo-ulnaris and M. extensor digitorum communis are shallower and less extensive.
Tibiotarsus. The specimen is too damaged to permit standard measurements for comparison with other taxa. The following measurements, taken from the specimen as preserved, give an indication of the general size: proximal width 15.5 mm; proximal depth (measured from the facies articularis medialis to the crista cnemialis lateralis in a direct anteroposterior line) 20.2 mm; in both cases, the actual value would be considerably greater, particularly for the latter measurement.
The crista cnemialis cranialis is broken; the remaining base is straight and does not extend far distally, although some distance further than the crista cnemialis lateralis. Their junction, at about a right angle, is marked by a shallow sulcus intercnemialis. The crista patellaris is straight, in both proximal and lateral views. The incisura tibialis is deeply incised, concave and moderately narrow; at its proximal end, it merges with a shallow excavation on the proximal surface. Its posterior border flares abruptly medially as the anterior side of the rather narrow and strongly rounded facies articularis lateralis. Much of the proximal surface of the facies is missing but what remains indicates that this was markedly raised. The circular fossa retropatellaris is deeper and smaller than the flattened and shallowly concave facies articular medialis. The circular attachment for M. femorotibialis medialis is on the end of the crista medialis, facing posteromedially. The facies gastrocnemialis is relatively flat from the medial side of the crista cnemialis cranialis along the medial side of the shaft, before curving smoothly into the posterior face.
General agreement in morphology and the association of the Riversleigh tibiotarsal fragment with other stork material from Bitesantennary Site are used as the basis for allocating it to this taxon.
Tarsometatarsus. The eminentia intercotylaris is directed anteroproximally at about 50° from the horizontal, projecting further anteriorly than the cotylae. The rounded anterior side is elliptical (in anterior view) with its long axis directed proximodistally; from the apex, it slopes posterodistally on its posterior side. The lateral side drops directly to the cotyla lateralis; the medial side extends as a long sloping ridge, confluent with the anterior rim of the cotyla medialis to past its mediolateral midpoint. The apex is slightly broader than the base (in medial view).
The cotylae are roughly equal in both their anterior and posterior extents and thus in overall depth. In proximal view, the cotyla medialis is roughly circular over most of its border, with the anterior side flattened. Posterior to the anterior rim, the cotyla meets medial side of the eminentia intercotylaris in a smooth, gradual slope. The medial rim projects strongly proximally to a narrow edge. In anterior view, the cotyla medialis is situated more proximally than the cotyla lateralis.
The area intercotylaris immediately posterior to the eminentia intercotylaris has a shallow depression between the eminentia intercotylaris and the borders of the cotylae. It then slopes posteriorly to another, much more extensive depression just anterior to the hypotarsus. The medial side of this section is higher than the lateral side.
The hypotarsus has two cristae hypotarsi separated by a broad, open, moderately deep sulcus hypotarsi. There is a very thin, low ridge running through the sulcus parallel to the crista lateralis hypotarsi. On the lateral side of this low ridge, the sulcus slightly excavates the base of the crista lateralis hypotarsi such that this is narrower than the posterior margin. The configuration of the crista medialis hypotarsi cannot be assessed. The main axis of the hypotarsus runs through the eminentia intercotylaris about 20° medial to the anteroposterior midline. The hypotarsus is separated from the ridge of the lateral shaft surface by a moderately deep furrow and from the medial shaft surface by broader and shallower furrow; both furrows contain a small nutrient foramen.
The anterior face is excavated at the proximal end by a deep fossa infracotylaris dorsalis, which continues distally as a wide sulcus extensorius. The deepest part of this basin contains two foramina vasculare proximale at the same level, immediately proximal to the round tuberositas m. tibialis cranialis. On their medial side, the sulcus extensorius is prominent, bordered both medially and laterally by narrow ridges.
Viewed anteriorly, the lateral and medial margins of the proximal fragment of shaft converge distally. Distal to the rim on the medial side, the shaft curves medially to a projection level with the cotyla, before angling proximally into the medial rim of the cotyla. The lateral side also swings outwards to meet the rim of cotyla lateralis, but not as strongly. The curvature of the anterior shaft face to the eminentia intercotylaris is greater on the lateral side because of an anterior projection distal to the cotyla lateralis. A rounded projection lateral to the cotyla lateralis originates just distal to the rim, then collapses to a low, flat, broad ridge forming the lateral shaft surface. There is a thin, low ridge on the lateral shaft surface that starts near the midpoint of the cotyla, trending posterodistally towards the midline of the shaft, then more distally. On the medial side, the projection on the proximal end of the medial shaft face continues distally as the broad, rounded shaft margin.
On the distal end, the small portion of shaft that is retained is narrow before flaring to meet the trochleae. Its lateral margin is straight, the medial more curved to join trochlea metatarsi II (in dorsal view). There is a deep groove running proximodistally proximal to, and level with, the lateral border of the trochlea metatarsi III. It meets the foramen vasculare distale, which is obscured on the dorsal surface. The medial side of the dorsal face slopes to join the medial surface; the junction of the dorsal and lateral faces is rather abrupt.
On the plantar surface, the sulcus supratrochlearis plantaris is moderately shallow. It is not strongly demarcated laterally, but medially is confined by a moderately high, narrow ridge connecting the base of the trochlea metatarsi II and the fossa metatarsi I. This long, elliptical fossa is situated on the proximal end of the ridge. There is a low, very thin ridge running lateral to the midline and meeting the foramen vasculare distale, which is prominent on this side.
This fragment is markedly compressed dorsoplantarly. The dorsal surface is more or less straight (in medial view), with the dorsal side of the trochlea metatarsi III projecting beyond it. The ridge supporting the fossa metatarsi I protrudes further plantarly than does the rest of the plantar shaft surface. The sulcus intertrochlearis lateralis is broader and deeper than the sulcus intertrochlearis medialis.
The trochleae are arranged in a curve, viewed distally. The trochlea metatarsi II lies at an angle of about 10° medial to the dorsoplantar midline; the trochleae metatarsi III and IV are tilted laterally about 10° and 20° from this midline, respectively. The trochlea metatarsi IV projects further dorsally and distally than does the trochlea metatarsi II, but neither projects as far as the trochlea metatarsi III. The trochlea metatarsi II extends further plantarly than the trochlea metatarsi IV. The trochlea metatarsi II is rounded dorsally and distally; its plantar border comprises a triangular projection, with a wide fovea lig. collateralis occupying much of the medial surface. The trochlea metatarsi III is grooved on its dorsal surface, with the lateral side projecting slightly further distally. The other trochleae lack grooves on their dorsal surfaces, although the trochlea metatarsi IV has a shallow groove on its distal surface. The dorsal surface of the trochlea metatarsi IV is tilted laterally. The distoplantar corner forms a triangular projection, projecting beyond the rest of the trochlea, but it and the fovea lig. collateralis are less prominent than on the trochlea metatarsi II.
The two proximal fragments differ in size, but share the same morphology. The magnitude of the morphometric differences are within that exhibited by modern taxa ( Table 4; see also Hancock et al., 1992 for measurements of living species).
Taphonomy
The occurrence of a stork in Wayne’s Wok and White Hunter Sites is not surprising. Both support rich local faunas comprising species of a range of vertebrate groups, including many indicative of aquatic environments. In contrast, Bitesanntennary Site is an initially unlikely source of such a bird. It has also yielded many species (almost all bats with 11 species recorded; Hand, 1997), but on both faunistic and geological bases it is considered to be a cavefill deposit. The other somewhat anomalous occurrence found here is that of the bulungamayine kangaroo Ganguroo bilamina ( Cooke, 1997b) .
There are several explanations for the presence of the non-bats Ciconia louisebolesae and G. bilamina in this cave deposit (see Baird, 1991b). It is possible that they used the cave as a shelter ( Ganguroo ) or there may have been a pool or other suitable foraging habitat within its immediate entrance. Otherwise the cave may have served as a predator’s lair and these species represent prey items, or their carcasses may have been washed into the cave from outside after death. An entrance of the cave may have opened upward through the roof rather than to the side and thus served as a pitfall trap for unwary animals. The skeleton of a Yellowbilled Spoonbill Platalea flavipes , a bird of roughly similar size, shape and habits as Ciconia storks, has been recovered from Weekes Cave, South Australia; van Tets (1974) considered that it “may have become trapped in the sink hole after blundering into it” in search of residual water after a wet period in the usually dry environment.
| QM |
Queensland Museum |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |