
Hypacrosaurus altispinus, BROWN, 1913
|
publication ID |
https://doi.org/10.1111/j.1096-3642.2009.00611.x |
|
persistent identifier |
https://treatment.plazi.org/id/5B5087D2-FFEE-EF6E-FC98-D0EBFB8B3A5C |
|
treatment provided by |
Valdenar |
|
scientific name |
Hypacrosaurus altispinus |
| status |
|
HYPACROSAURUS ALTISPINUS BROWN, 1913
Hypacrosaurus altispinus Brown 1913: 395 (original description).
Cheneosaurus tolmanensis Lambe 1917: 18 (plates VI and VII)
Hypacrosaurus altispinus Gilmore 1924a:50 (plates
XI and XII)
Synonymy: Cheneosaurus tolmanensis Lambe, 1917
Holotype: AMNH 5204 About AMNH ; incomplete postcranial skeleton, with vertebral column consisting of the last eight dorsal vertebrae and two proximal caudal vertebrae, all neural spines incomplete distally, both ilia, right ischium, incomplete right pubis, several ribs.
Locality and horizon: The location of the holotype quarry is approximately 15 km east of the town of Trochu, Alberta (Fig. 1), on the east side of the Red Deer River Valley, about 6 km north of Tolman Bridge; The precise location of the quarry is unknown, but its general location was believed to be relocated by L. S. Russell (unpublished manuscript, ROM archives), who gives the Legal Land Description as: Legal Subdivision 8, Sec. 2, Tp. 34, Rge. 22, W of 4th Meridian. The specimen was collected from the Horseshoe Canyon Formation, probably from just above the base of unit 4 ( sensu Eberth, 2004). The quarry is stratigraphically high within the Horseshoe Canyon Formation, above the Drumheller Marine Tongue ( Eberth, 1997), and is of early Maastrichtian age.
Paratypes: AMNH 5206 About AMNH , AMN 5204, AMNH 5217 About AMNH , AMNH 5272 About AMNH .
Plesiotype: CMN 8501 ( Fig. 4 View Figure 4 ).
Referred material: Virtually all of the known cranial material of H. altispinus was examined for this study (see Appendix S1 for the hypodigm). Particularly important specimens include: CMN 2246, CMN 2247, CMN 8501, CMN 8675, ROM 702, ROM 789, AMNH 5248, TMP 82.10.01, and TMP 2006.15.01.
Amended diagnosis: In the most recent diagnosis by Lull & Wright (1942), the vast majority of characters cited as diagnostic for H. altispinus in fact characterize most lambeosaurines. Four characters are retained as valid autapomorphies for H. altispinus : ‘external naris opening far forward’, ‘no nasal vacuity on side of crest’, ‘angle of profile of crest and muzzle 140 degrees’, and ‘Ischium long, with large, terminal foot-like expansion’. The revised diagnosis provided here is as follows. Lambeosaurine hadrosaurid of the genus Hypacrosaurus that has the following autapomorphies: constricted external naris bound caudally by the enlarged dorsal margin of the premaxilla, postorbital with an unbranched squamosal process, jugal with an angular ventral flange, and the premaxilla–nasal fontanelle absent throughout ontogeny. Hypacrosaurus altispinus has one autapomorphy in the postcranial skeleton, a relatively enlarged ischial foot in which the depth of the foot is over 26% the length of the ischial shaft (convergently evolved in Parasaurolophus cyrtocristatus, Ostrom 1961a ).
Biostratigraphy: Hypacrosaurus altispinus is known from only a restricted stratigraphic interval within the Horseshoe Canyon Formation (Fig. 1) that spans informal stratigraphic units 2–4 of Eberth ( Eberth, 2004) or Edmonton Member B ( Russell & Chamney, 1967). Recent 40 Ar/ 39 Ar dates from the HC-15 bentonite (within the Drumheller Marine Tongue) and bentonites within the Knee Hills Tuff (within the Battle Formation) constrain the Hypacrosaurus specimens to between 70.44 ± 0.17 and 67 Mya, an approximately 3-million-year interval, although the temporal span of units 2–4 themselves, not including overlying unit 5 and the Whitemud Formation, is probably significantly shorter ( Eberth, 1997; Eberth, 2004).
Remarks: Gilmore (1914) reported the discovery of material that he referred to Hypacrosaurus from the Two Medicine Formation of Montana one year after the type description by Brown (1913). He subsequently described the material, which included a jugal and a dentary associated with a large part of the postcranium ( Gilmore, 1914, 1917). In both publications, Gilmore emphasized his inability to distinguish the material from the Albertan H. altispinus material, and therefore he tentatively assigned the specimen to? H. altispinus . Work by Morris (1978) demonstrated that this specimen is a taxonomically indeterminate lambeosaurine ( Morris, 1978). Published descriptions ( Gilmore, 1917; Morris, 1978) confirm that this material does not exhibit any autapomorphies of H. altispinus . However, tall craniodorsal neural spines suggest assignment to the genus Hypacrosaurus (and not Lambeosaurus as suggested by Morris, 1978), and its stratigraphic occurrence is consistent with referral to H. stebingeri , as suggested by Horner & Currie (1994); however, this cannot be confirmed in the absence of more complete cranial material.
DESCRIPTION
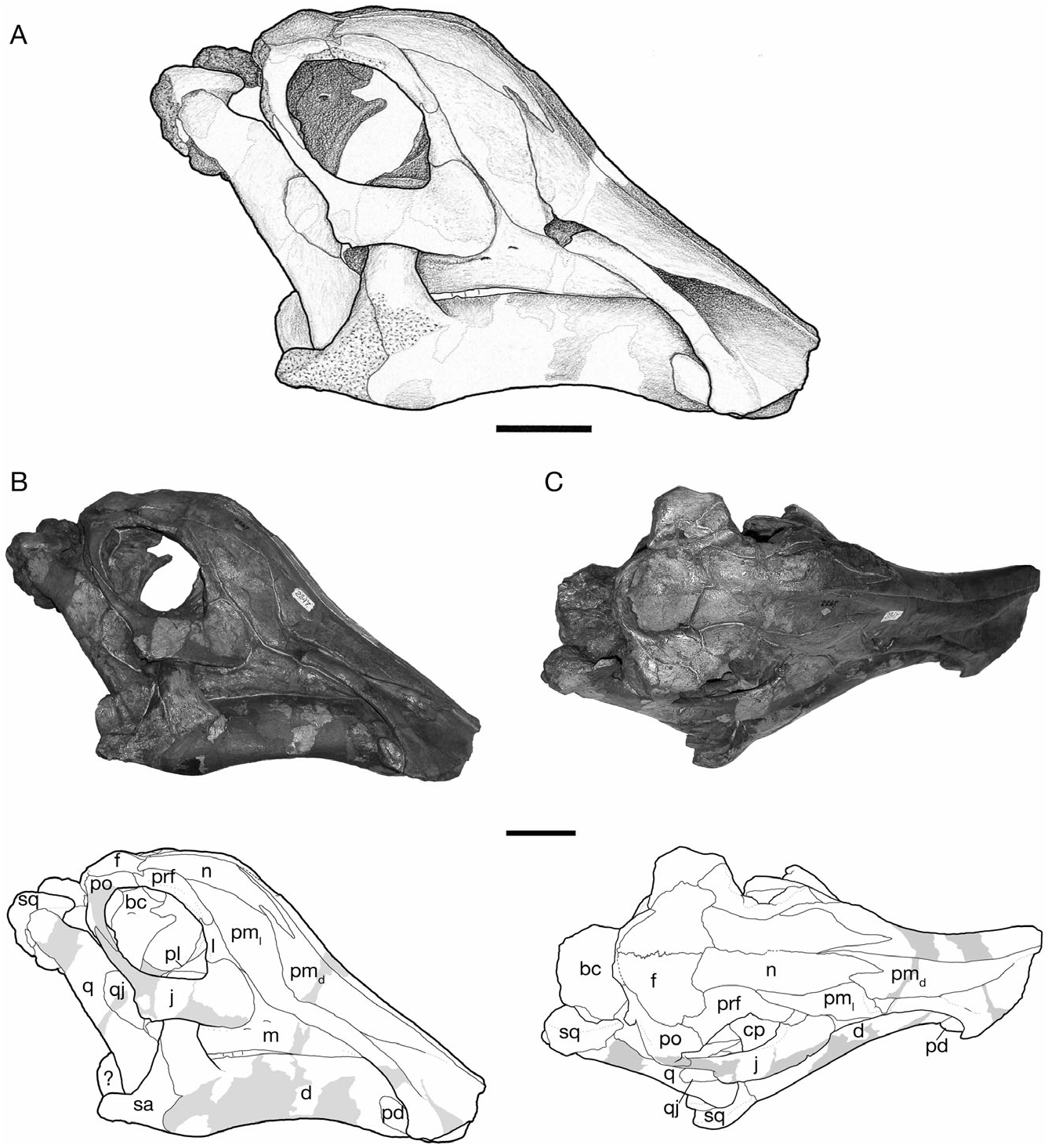
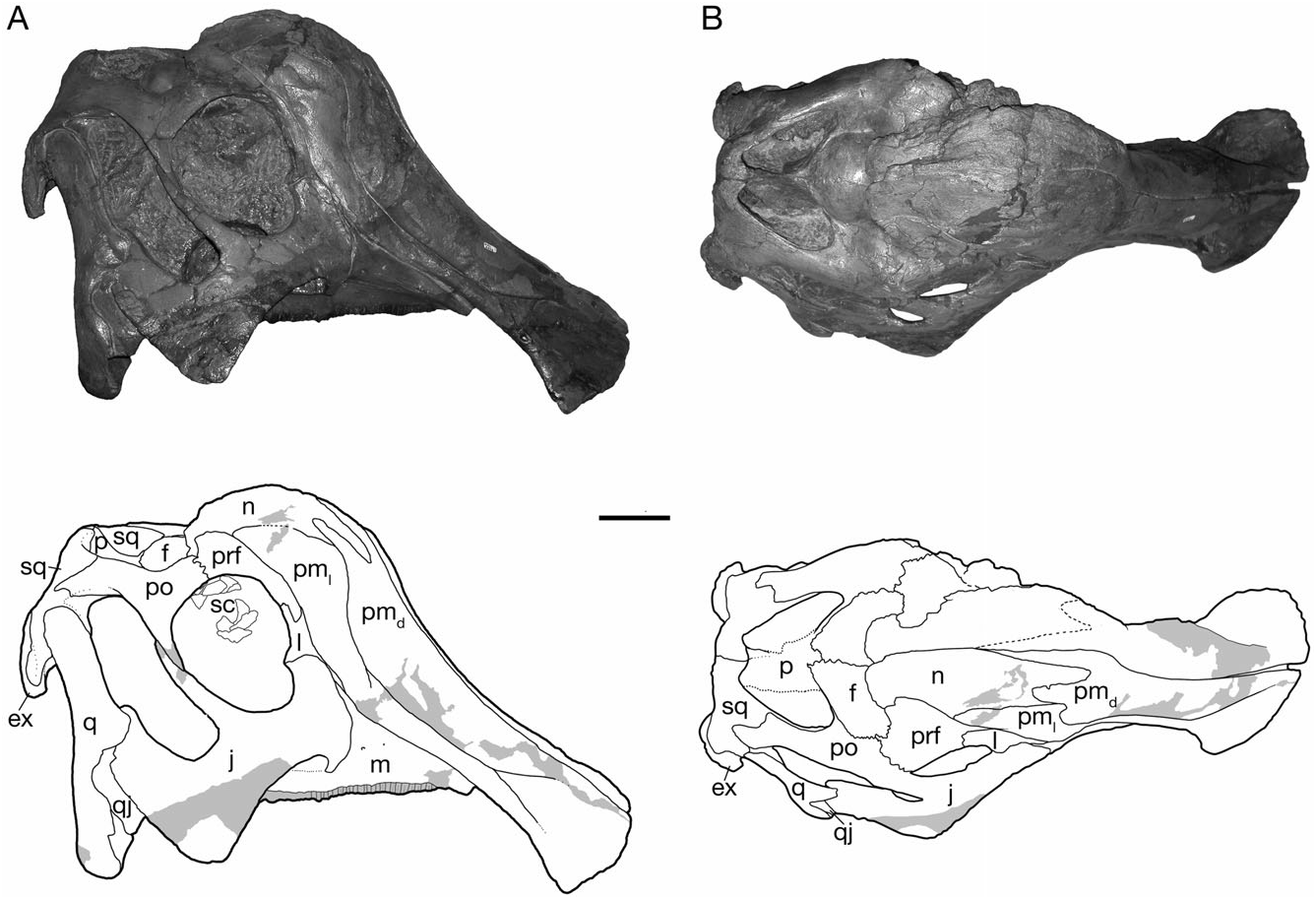
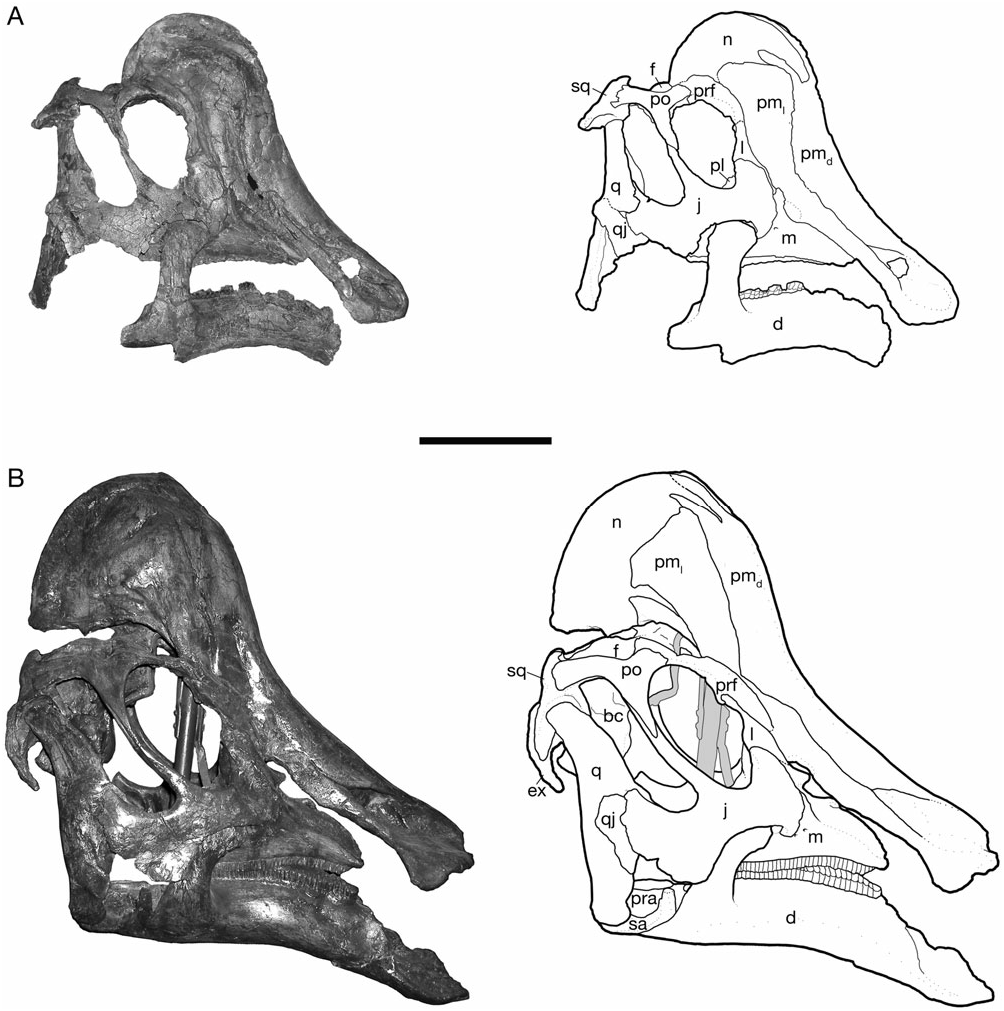
Gilmore (1924a) and Lambe (1917, 1920) described the general form of the skull and the relationships of its constituent bones in H. altispinus , which follows closely that of Corythosaurus described in detail by Ostrom (1961). The cranial growth series of H. altispinus spans the active phase of cranial crest development. The smallest specimen, CMN 2247 ( Fig. 2 View Figure 2 ), is under 45% of the total length of the largest skull: it has an approximate skull length of 365 mm, with a complete maxilla that measures 149 mm in total length. CMN 8675 is the largest known individual, with an estimated skull length of 830 mm, and a maxilla that is 356 mm in length ( Fig. 5 View Figure 5 ).
Cranial crest
The crest is incipient in small individuals, and becomes progressively larger through ontogeny, with the nasal and caudolateral lateral processes undergoing the greatest relative increase in size ( Figs 2–5 View Figure 2 View Figure 3 View Figure 4 View Figure 5 ). The joint between the nasal and the caudodorsal process of the premaxilla migrates from an antorbital to a supraorbital position. In large, presumably adult individuals the overall shape of the laterally compressed cranial crest is variable, ranging from approximately as tall as long (CMN 8501; Fig. 4 View Figure 4 ) to considerably longer than tall (CMN 8675; Fig. 5C View Figure 5 ), the latter condition being similar to H. stebingeri (MOR 455 and MOR 549). The crest closely resembles that of Corythosaurus and H. stebingeri in its semicircular profile, but differs from all lambeosaurines in a number of aspects of its internal construction: the caudolateral process of the premaxilla forms a large, arched contribution to the central external crest surface. The nasal, although of the same general morphology as Corythosaurus , is reduced. Of particular interest, the joint between the nasal and the dorsal process of the premaxilla is similar to Corythosaurus and H. stebingeri in the presence of an interfingering joint, in which a long process of the nasal laps onto the caudodorsal process of the premaxilla in the rostral region of the crest ( Gilmore, 1924b; Evans et al., 2005). However, unlike Corythosaurus and H. stebingeri , H. altispinus has an extremely small, or potentially lacks, the lower branch of the anterior end of the nasal. All known species of Lambeosaurus have an unbranched nasal that does not overlap the dorsal process of the premaxilla ( Evans et al., 2005). In addition, H. altispinus lacks lateral fontanelles in the crest (premaxilla–nasal fontanelles; Maryañska & Osmólska, 1979) that are characteristic of Corythosaurus and Lambeosaurus .
Crest cavities (nasal passages): Based on inspection of broken and/or partially disarticulated specimens, Weishampel (1981) reconstructed the airway of H. altispinus as a relatively straight tube from the external naris to a supraorbital ‘common median chamber’ within the crest, which in turn exits ventrally via a single opening into the interorbital space ( Weishampel, 1981). Evans, Ridgely & Witmer (2009) recently described the internal nasal passage anatomy of H. altipinus (ROM 702), as well as that of the brain cavity endocast, in detail on the basis of new CT scan data. This data reveals that the internal passages of H. altispinus differ dramatically from the previous reconstruction of Weishampel (1981), which is also corroborated by breakage of the articulated premaxilla and nasal of CMN 8675 and ROM 702 ( Fig. 5 View Figure 5 ). Of note, contrary to the previous reconstruction, an s-loop is indeed present, but is modified compared with the Corythosaurus condition ( Weishampel, 1981); the rostral (lower) half of the s-loop nasal passage is greatly elongated, extending to the level of the skull roof. Lateral diverticula are similar to Corythosaurus , but do not extend beyond the small common median chamber ( Evans et al., 2009).
Premaxilla
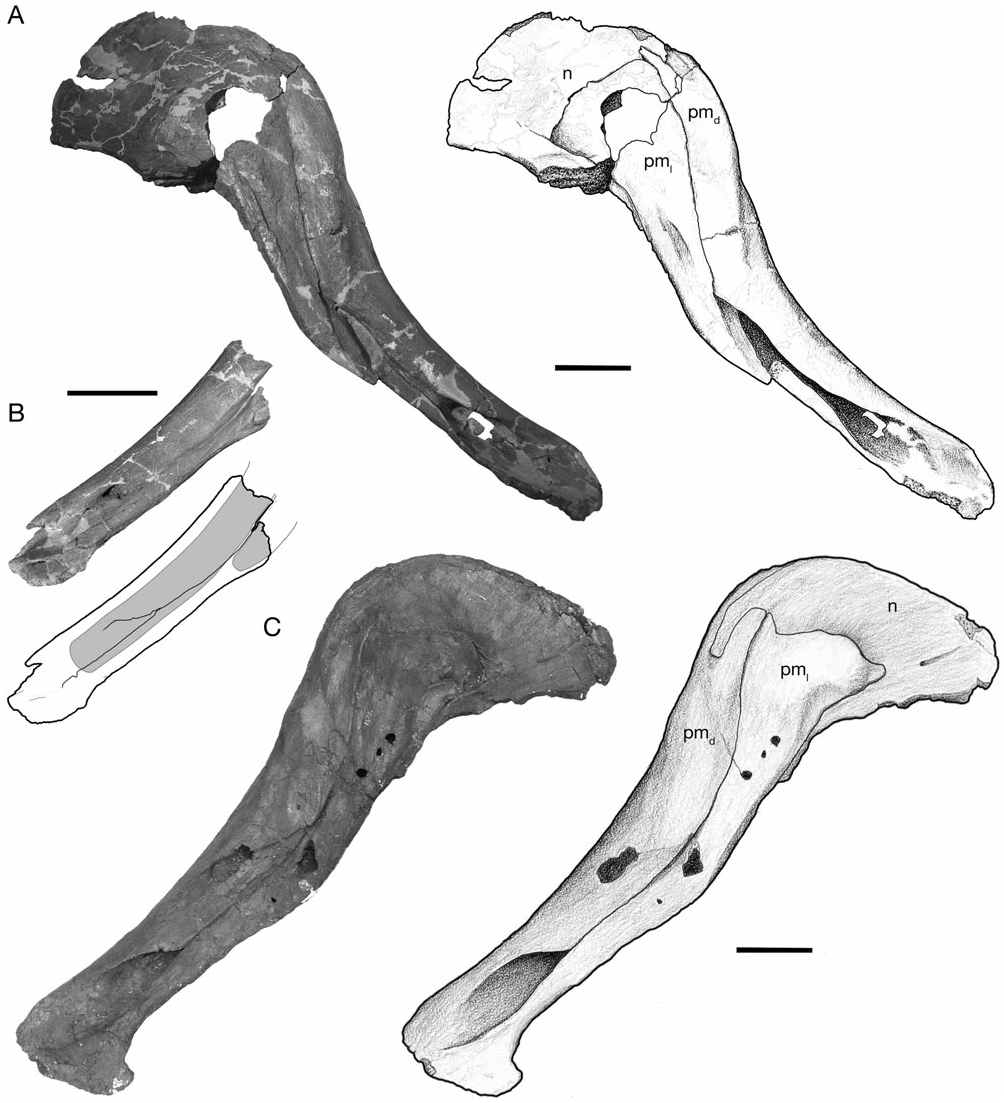
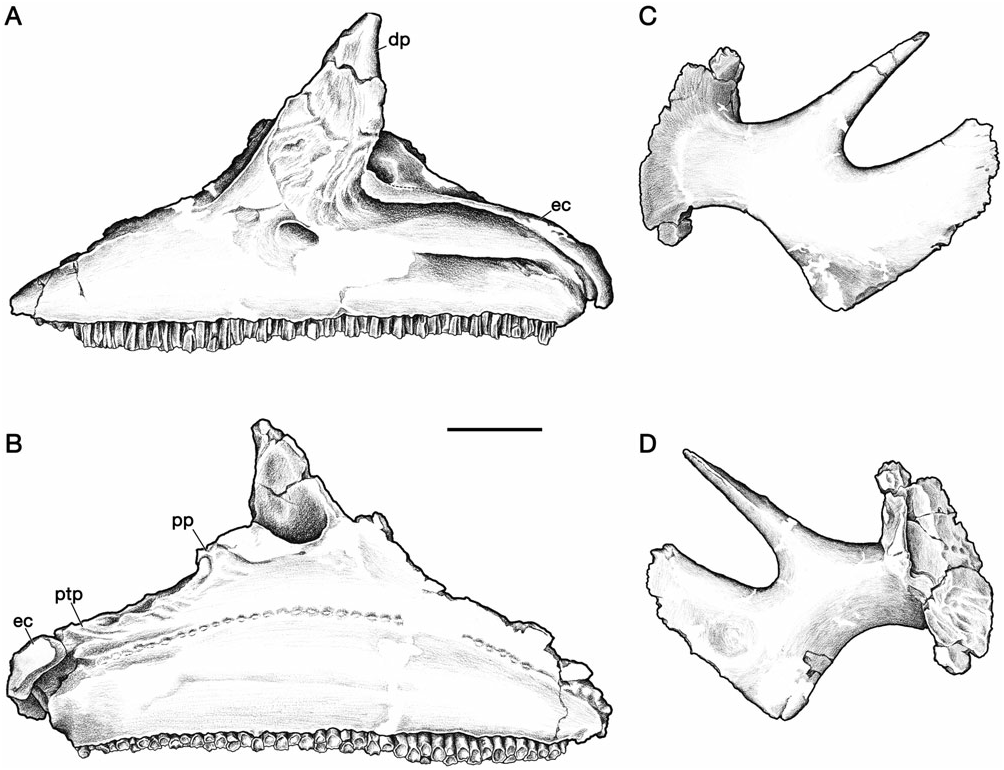
The premaxilla is a large, complex bone that forms the edentulous rostral ‘bill’, as well as an increasingly larger portion of the cranial crest through ontogeny ( Figs 2–5 View Figure 2 View Figure 3 View Figure 4 View Figure 5 ). As described by Gilmore (1924a), and typical of lambeosaurines, the premaxilla contacts the nasal caudally and the maxilla, lacrimal, and prefrontal along its caudodorsally inclined ventrolateral ramus. The straight interpremaxillary suture extends along the midline of the skull to a point caudal to the apex of the crest, but it does not reach the terminal margin of the crest as in Lambeosaurus (e.g. CMN 2869 and TMP 81.37.01). Rostrally, the premaxillae form a broadly arcuate oral margin in dorsal view that reaches a maximum breadth of slightly less than twice the width of the postoral constriction. In ventral view, the rostral bill has a peripheral denticulate margin that is separated from a thickened palatal region by an arcuate transverse groove that mirrors the curvature of the rostrum, as in other hadrosaurids ( Horner et al., 2004). The left and right nasal passages are completely separated in the snout region, and the external naris is surrounded entirely by the premaxilla ( Gilmore, 1924b).
The construction of the external naris in H. altispinus is unique among lambeosaurines ( Morris, 1978). The dorsal margin of the premaxilla abruptly deepens along the caudal margin of the naris to contact the caudolateral process lateral to the vestibule ( Figs 5B, C View Figure 5 ). The lateroventral margin of the naris is identical to that of Corythosaurus , Lambeosaurus , and H. stebingeri . The enlargement of the dorsal narial margin relative to the condition in the latter taxa results in a lacriform naris that is relatively smaller (or ‘constricted’), and more rostrally positioned than in other corythosaurins. This distinctive narial morphology is present in the smallest known juvenile specimen (CMN 2247), and throughout the known ontogenetic series. In H. stebingeri (MOR 549), Corythosaurus , L. lambei , and L. magnicristatus , the naris is elongate, and the general features of its morphology are also consistent through the known ontogenetic series of these taxa ( Evans et al., 2005). Parasaurolophus walkeri also has a constricted naris, but in this case it is the lateroventral rim of the premaxilla that is enlarged relative to the dorsal margin (Evans & Reisz, 2007).
The shallowly concave rostral margin of the premaxilla between the snout and the ascending caudodorsal process forms an angle that is not less than 140°. The crest–snout angle is largest in the smallest specimen (CMN 2247, 163°), but it does not change drastically with growth as in Corythosaurus and Lambeosaurus ( Dodson, 1975) . Variation in this angle ranges from 141° (ROM 702) to 159° (CMN 8675) in adults. The open angle and shallowly concave facial profile of H. altispinus overlaps only with H. stebingeri (MOR 549, 140°; MOR 455, 147°), and possibly with Olorotitan ( Godefroit et al., 2003) . In Corythosaurus and Lambeosaurus , juveniles have facial profiles similar to Hypacrosaurus , but the crest–snout angle becomes progressively smaller through ontogeny, and the crest–snout angle in adults does not exceed 135° in Corythosaurus and 116° in Lambeosaurus ( Dodson, 1975) .
The caudodorsal and caudolateral processes expand through ontogeny, and form a large forward part of the cranial crest in adults. The caudodorsal process extends caudally beyond the lateral process, where it is shrouded laterally by the nasal, but it does not separate the nasals along the caudal margin of the crest, as in Lambeosaurus ( Horner et al., 2004) . The caudodorsal process unites with the nasal in an interlapping and interfingering joint in which a small process of the premaxilla slightly overlaps the nasal, and the nasal has a similar, but larger process that overlaps the lateral surface of the premaxilla. The caudolateral process is a mediolaterally thin sheet of bone that in adults forms a lancet (ROM 702 and CMN 8501) to quarter-circle shaped (CMN 8675) contribution to the center of the crest when viewed in lateral aspect. The lancet shape of the caudolateral process makes it difficult to identify distinct rostral and caudal ‘lobes’ of this process, evident in Lambeosaurus and Corythosaurus ( Horner et al., 2004) . In adults, the caudolateral process forms a relatively larger part of the central region of the crest (in lateral view) than in Corythosaurus , and in this regard more closely resembles Lambeosaurus and H. stebingeri , in which the rostral lobe is enlarged. The dorsal and caudal margin of the caudolateral process is tightly jointed with the nasal in small and large individuals. In ROM 702 and CMN 8675, the caudolateral process and adjacent nasal have several small, rounded spurs that interlap along their shared margin.
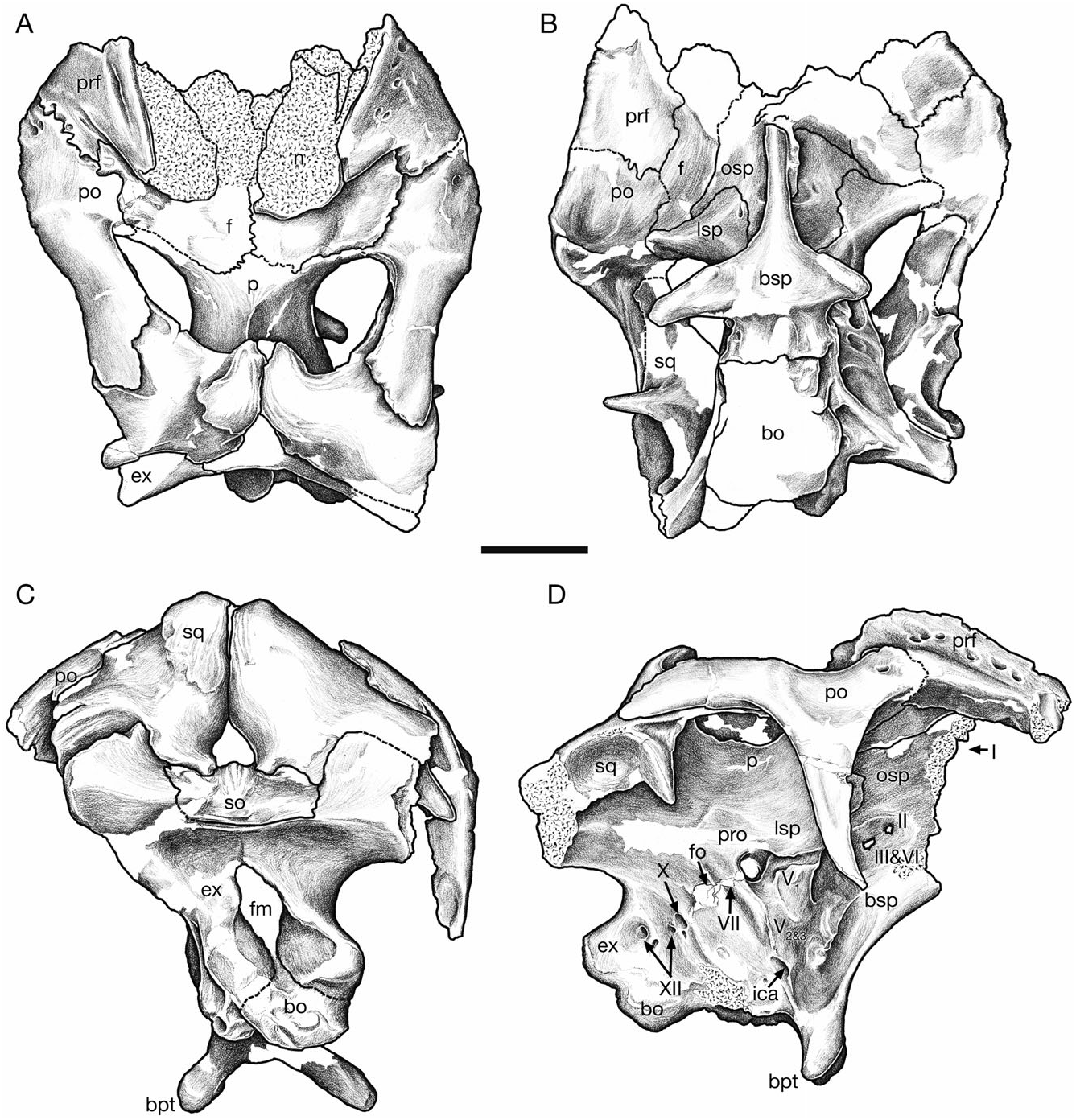
Nasal
The nasal forms the dorsal and caudal portion of the cranial crest in adults, and closely resembles that of Corythosaurus in shape and sutural relationships (Ostrom, 1961), but is less deep rostrally, where it is not strongly bifurcated. The nasal also bears a distinct resemblance to that of Velafrons coahuilensis ( Gates et al., 2007) . The nasal forms between 52% (CMN 8501) and 55% (CMN 8675) of the lateral crest surface in large adults ( Fig. 5 View Figure 5 ). Rostrally, the joint between the nasal and caudodorsal process of the premaxilla is best preserved in CMN 2247 ( Fig. 2 View Figure 2 ), the smallest known specimen in the series, and in the large individuals represented by CMN 8675 and ROM 702 ( Fig. 5 View Figure 5 ). The nasal has a long, rostrally tapered process that overlaps the surface of the dorsal premaxillary process laterally. The shape of this process varies from being distally pointed (CMN 2247) to bluntly round (CMN 8675) in complete, undamaged specimens. The general morphology of the nasal– caudodorsal process joint does not change significantly through ontogeny, although the relative position of the joint migrates dorsally and caudally with growth, as in other corythosaurins ( Lull & Wright, 1942; Evans et al., 2005).
Although relatively simple superficially, the nasal has a complex internal structure that reflects the morphology of the nasal passages it encloses ( Evans et al., 2009). In disarticulated adult crests (ROM 702 and CMN 8675), the medial surface forms a broad, flat sutural surface dorsally and caudally for the opposite nasal along the caudal periphery of the crest, as in Velafrons ( Gates et al., 2007) and Corythosaurus ( Sternberg, 1935) . Rostrally, the nasal has large lateral and medial fossae that are separated by a thin, vertical, parasagittal lamina. The lateral fossa, together with the caudolateral process of the premaxilla, encloses the lateral diverticulum. The medial fossa forms the common median chamber of the crest with the opposite nasal ( Weishampel, 1981; Evans et al., 2009).
Ventrally, the nasal broadens mediolaterally, and forms a thickened base where it meets the frontal (ROM 702). In disarticulated crests, the lateral base of the nasal has a shallow, semicircular sutural facet for the medial flange of the prefrontal. From the prefrontal facet, the nasal extends rostrally, and curves ventrally in accord with the rostrodorsal margin of the orbit ( Fig. 5 View Figure 5 ). Here, the nasal clearly has an extensive lateral contact with the prefrontal as it wraps around the orbit, and its extreme rostroventral extent suggests that the nasal may also contact the medial part of the lacrimal, although there is no direct evidence of a contact between the two bones, and no external contact can be observed in articulated skulls (e.g. CMN 8501).
Maxilla
The maxilla is a large and triangular bone, the base of which is formed by the long tooth-bearing ramus ( Fig. 6 View Figure 6 ). Maxillae span a size range from 149 mm (CMN 2247) to 356 mm (CMN 8765) in total length. In ROM 702, the complete tooth row is 279.9 mm, with 46–47 vertical tooth families, twelve (~25%) of which contain two functional teeth. The number of maxillary tooth families in ROM 702 are higher than the 42–45 range scored for Hypacrosaurus by Horner et al. (2004: appendix 1, character 1). This highlights the variability in tooth family number, a trait well known to be size-dependent in hadrosaurids ( Dodson, 1975).
In lateral view, the acuminate dorsal process of the maxilla is directed caudodorsally from the horizontal alveolar ramus. Its lateral surface consists predominantly of the rugose sutural surface for the rostral process of the jugal. A large foramen at the rostral base of the dorsal process, shrouded by the premaxilla in articulated skulls, opens onto the dorsomedially canted maxillary shelf. Additionally, three large foramina pierce the lateral surface of the maxilla along the ventral base of the dorsal process. The two most caudal foramina are separated by a thin septum internally, but exit together via a large, oval-shaped opening ventromedial to the rostral end of the ectopterygoid ridge, below the jugal. The number, size, and position of these foramina are individually variable in H. altispinus and other lambeosaurines (e.g. Corythosaurus casuarius, ROM 1933 ), although the largest of the foramina are invariably located beneath the jugal.
The ectopterygoid ridge forms a prominent horizontal lip along the caudal half of the maxilla, from the ventral base of the jugal sutural surface to the caudal end of the bone. The ectopterygoid ridge is particularly robust in the largest individual (CMN 8675), where it is ventrally concave, and projects laterally over 10 mm beyond the body of the bone at its midpoint. The ridge separates the caudal half of the maxilla into a ventral alveolar ramus and a transversely broad and flat ectopterygoid shelf. The palatine and pterygoid processes extend dorsally from the medial margin of the ectopterygoid shelf, and contact the palatine rostrally and the pterygoid caudally, respectively. The base of the palatine process to the end of the bone is notably rugose for the attachment of the palatal bones. Unlike the tall palatine process, the pterygoid process is only incipiently developed. The medial surface of the maxilla is essentially flat, with a dorsally convex row of alveolar foramina typical of hadrosaurids ( Fig. 6 View Figure 6 ).
Jugal
To reflect the increased width of the temporal region of the skull relative to the snout, the jugal is bowed laterally from its rostral contact with the maxilla to its union with the quadratojugal and quadrate below the infratemporal fenestra ( Figs 6 View Figure 6 , 7 View Figure 7 ). The postorbital process extends dorsally to meet the descending process of the postorbital to complete the postorbital bar. The postorbital process and the dorsal margin of the caudal blade form a tight parabolic arch that defines the lower half of the infratemporal fenestra. The angle between the blade and the postorbital process is more variable in other corythosaurins ( Corythosaurus and Lambeosaurus ), and is typically larger than in H. altispinus .
The rostral process is dorsoventrally expanded as in other lambeosaurines, but the rostral plate is relatively short with a straight rostral margin that is vertical to slightly caudodorsally inclined in lateral view. The rostral process changes shape minimally over the observed size range: it has a slightly more rounded profile in CMN 2247, but is rostrocaudally short with a straight rostral margin in CMN 2246 and throughout the remainder of the ontogenetic series. In L. lambei , Corythosaurus , and H. stebingeri , the rostral process tends to be broader with a distinctly more rounded profile, particularly in juveniles, and it shortens only slightly through ontogeny in these taxa (e.g. ROM 758, ROM 869, CMN 34825, ROM 870, and ROM 1933). Among North American corythosaurins, the shape of the rostral process in H. altispinus approaches the condition seen in Lambeosaurus magnicristatus (Evans & Reisz, 2007) . Olorotitan is also reported to have a short rostral process with a straight rostral margin ( Godefroit et al., 2003).
The shape of the subtemporal blade, or caudal process, and the ventral flange is unknown in both of the smallest juvenile specimens of H. altispinus : it is missing in CMN 2247, and is largely reconstructed in CMN 2246. The blade is relatively complete in TMP 2006.15.01 (a large juvenile) and TMP 82.10.01, a subadult individual ( Fig. 4A View Figure 4 ). The ventral flange of TMP 82.10.01 is angular, and the blade lacks a distinct mid-ventral constriction, as in Corythosaurus , Lambeosaurus , and P. walkeri . Most other individuals (ROM 702, ROM 789, and CMN 8501) have an angular ventral margin of the jugal, as in TMP 80.10.01. However, the ventral margin of the jugal in TMP 2006.15.01 and CMN 8673 (a subadult specimen similar in size to TMP 82.10.01) is somewhat more rounded than angular ( Fig. 7A View Figure 7 ), and an offset rostroventral lobe, as in most lambeosaurines, is clear. The angular nature of the ventral margin is related to the relatively steep, caudodorsal orientation of the blade and the constriction of the lower margin of the temporal fenestra in H. altispinus , relative to other lambeosaurines. Interestingly, a similar condition occurs in V. coahuilensis ( Gates et al., 2007) and MOR 609- 20. Although the shape of the blade is to some extent individually variable, as in all lambeosaurines, it is sufficiently distinct to be incorporated into phylogenetic analyses ( Suzuki et al., 2004). The medial surface of the blade has an extensive suture with the quadratojugal that is marked by fine longitudinally oriented striations. A low, circular depression is present in the centre of the blade immediately rostral to the quadratojugal sutural surface (ROM 702, CMN 8674, and CMN 8675).
Quadratojugal
When in articulation, the quadratojugal incompletely separates the jugal from the quadrate in several ontogenetic stages (AMNH 5278, CMN 8501, ROM 789, and CMN 2246). An incomplete, disarticulated quadratojugal is somewhat unusual, in that the sutural surface for the caudal blade of the jugal is inset from the smooth, externally exposed lateral surface of the bone ( Fig. 7B, C View Figure 7 ).
Ectopterygoid
The ectopterygoid is a long, dorsoventrally thin bone that articulates with the flattened caudal half of the maxilla (on the ectopterygoid shelf), and is excellently preserved in articulation with the maxilla in CMN 8675 and ROM 702 ( Fig. 6 View Figure 6 ). The dorsal surface is smooth and gently bowed dorsally. In dorsal view, the rostrolateral margin is inset from the underlying ectopterygoid shelf. The rostral end does not contact the jugal, as in non-hadrosaurid hadrosauriformes such as Protohadros byrdi ( Head, 1998) . The medial edge is poorly defined rostrally in the available material, but it appears to roll dorsally and lap on to the base of the palatine process of the maxilla. The caudal half of the ectopterygoid expands transversely to reach its maximum breadth near its caudal end, beyond which it curves ventrally along with the maxilla. Where the medial and lateral margins extend beyond the underlying ectopterygoid shelf, a short process, termed here the pterygoid process of the ectopterygoid, projects medially behind the pterygoid process of the maxilla. A shallow parasagittal groove on the dorsal surface of this process has subtle rugosity for articulation with the pterygoid. The extreme caudoventral end of the ectopterygoid is separated from the maxilla by a lateral interposition of the pterygoid.
Vomer
The vomer has been identified previously in only two lambeosaurine specimens, CMN 8676 ( Co. intermedius ) and ROM 1933 ( Co. casuarius ) (Ostrom, 1961; Heaton, 1972). An almost complete vomer is also preserved in one specimen of H. altispinus, CMN 2246 ( Fig. 7D View Figure 7 ). This thin, triangular bone is still partially embedded in matrix between the caudal wall of the premaxillae and the basisphenoid, and is displaced dorsally from its natural position. A drill hole, presumably to accommodate mounting hardware, pierces the middle of the bone. Because of a lack of correspondence with CMN 8676, it is difficult to determine the correct anatomical orientation of the vomer of CMN 2246: it is described and illustrated here as preserved, with the ventral margin longest and parallel with the maxillary tooth row. The vomer is notable for its large size: the length of the straight ventral margin ( 109.5 mm) is 52% the length of the maxilla. It is dorsoventrally flattened and sharply pointed rostrally. It deepens caudally such that the rostrodorsal margin is strongly concave. The maximum height of 51 mm (measured perpendicular to the ventral margin) is reached at 80 mm from the rostral tip. Caudal to the apex, the bone has an irregular, jagged profile reflecting breakage, but the bone clearly expands caudoventrally relative to the apex. The lateral outline of the vomer differs from that in Corythosaurus (CMN 8676) as reconstructed by Heaton (1972), as well as from those of hadrosaurines ( Heaton, 1972; Horner, 1992), suggesting that this bone may provide a phylogenetic signal when its morphology is known in a larger sample of hadrosaurid taxa.
Palatine
The palatine, not previously described in H. altispinus , is best preserved in CMN 8674, TMP 82.10.01, and the smallest individual CMN 2247, although it is fragmentary in several other specimens (CMN 8501 and CMN 8675). In lateral view, the thin palatine is triangular, with three unequal sides. The short, rostral edge is slightly inclined rostrodorsally, and the long caudodorsal margin has an articular facet for the pterygoid along its entire length. The prominent rostrolateral process extends at a right angle from the main body of the bone, and contacts the jugal adjacent to the caudal margin of the maxillary dorsal process. In CMN 8674, this process is uniformly thin throughout its length, but expands dorsoventrally at its distal, articular end. The ventral margin of the rostrolateral process defines the dorsal margin of a round foramen, completed by the maxilla, and possibly by the ectopterygoid, that separates the ectopterygoid shelf from the region deep to the base of the dorsal process of the maxilla. Horner (1992) identifies this foramen as the homologue of the antorbital fenestra in hadrosaurids. The foramen is roughly similar in size to the rostral maxillary foramen that opens onto the rostral maxillary shelf, although it is unclear whether the two are connected. Rostromedially, the palatine probably contacts the vomer, although this cannot be confirmed with the available material ( Heaton, 1972). The ventral margin contacts the palatine process of the maxilla, as in other hadrosaurids.
Pterygoid
The lambeosaurine pterygoid is rarely illustrated or described in detail, and it has only been illustrated for Corythosaurus ( Parks, 1923; Ostrom, 1961; Heaton, 1972). The pterygoid of H. altispinus is preserved in articulation in CMN 2246 and disarticulated (but broken) in ROM 702, and it closely resembles that of Corythosaurus ( Fig. 8 View Figure 8 ). The caudal quadrate wing contacts the medial surface of the complementary pterygoid wing of the quadrate along its entire length. The thin, dorsal plate of the quadrate wing is not preserved in ROM 702, but the robust ventral margin is complete to its distal tip, which supports a small but distinct caudal continuation of the lateral sutural surface that interlocks tightly with a notch in the quadrate ramus (Ostrom, 1961; Heaton, 1972). It is unlikely that this joint would have had mobility in life. The mediolaterally expanded ventral ramus and the quadrate process form a fossa that becomes progressively deeper rostrally, and terminates in a trough-like articular surface that cradles the basipterygoid process of the basisphenoid. The ectopterygoid extends ventrally and laterally to wrap around the caudal end of the maxilla. A deep, circular pit occurs at the base of the quadrate, maxillary, and medial (pterygoid) processes in caudal view. Ostrom (1961) hypothesized this region as the attachment site for M. pterygoideus ventralis. The caudoventral margin has an elongate, roughened fossa at the rostral base of the ectopterygoid process that contacts the pterygoid process of the maxilla. The transversely thin palatine process, which extends rostrally from the body of the pterygoid ( Heaton, 1972), is not preserved in the available material.
Quadrate
A complete quadrate is present in three specimens: CMN 2246 About CMN , TMP 82.10 View Materials .01, and CMN 8501 About CMN . The lateral ramus is also relatively complete in the smallest individual ( CMN 2247 About CMN ), in which it is 141-mm high, about 46% of the height of the quadrate of CMN 8501 About CMN ( 307 mm). The quadrate does not differ significantly from that of other lambeosaurines (Ostrom, 1961) .
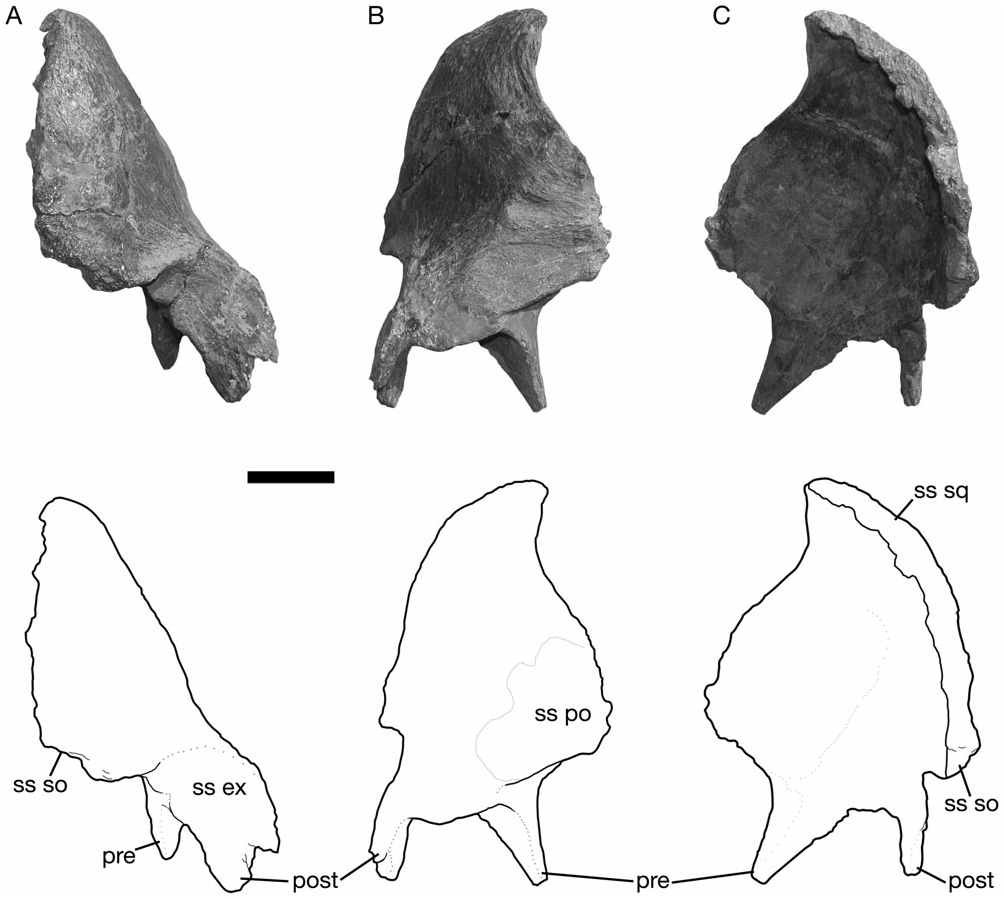
Squamosal
The squamosal is a thin, curved bone, concave both mediolaterally and dorsoventrally, that forms the caudal wall of the supratemporal fenestra and the internal half of the intertemporal bar ( Fig. 9 View Figure 9 ). It contacts the postorbital, the parietal, the quadrate, the exoccipital, and the supraoccipital (e.g. ROM 702 and CMN 8675). Rostrally, the squamosal extends along the medial surface of the postorbital as a thin, dorsoventrally tapered process that terminates at the rostral margin of the supratemporal fenestra. Ventrally, the quadrate cotylus is relatively deep and broadly ovate (with a parasagittal long axis). The cotylus is bordered rostrally by a triangular prequadratic process and caudally by a tongue-shaped process that buttresses the rostral base of the paroccipital process. At the caudomedial corner of the supratemporal fenestra, the squamosal arches rostrally at its contact with the parietal.
In caudal view, the lateral surface of the squamosal is steeply canted dorsomedially, as in other lambeosaurines ( Godefroit, Alifanov & Bolotsky, 2004a; Godefroit et al., 2004b), but distinct from the subhorizontal dorsal border in Jaxartosaurus , hadrosaurines (Horner, 1992), and non-hadrosaurid hadrosauroids ( Godefroit et al., 1998; Norman, 2002). In all of the available specimens, the squamosal contacts its complement at the midline above the supraoccipital. There is no indication that the parietal has any appreciable exposure between the squamosals, as in Jaxartosaurus , Aralosaurus , hadrosaurines, and non-hadrosaurid hadrosauroids. Ventromedially, a notch accepts the lateral boss of the supraoccipital. A large facet adjacent to the supraoccipital notch extends along the entire caudal surface of the postquadratic process, and contacts the exoccipital.
The disarticulated squamosals of AMNH 5248 reveal that the internal surface has a low, vertically oriented ridge that separates shallow lateral and medial fossae, hypothesized by Ostrom (1961) to mark the attachment sites of M. adductor externus medialis and M. adductor mandibulae externus profundus, respectively.
Lacrimal
The lacrimal is a small, wedge-shaped bone on the rostral margin of the orbit that contacts with the maxilla ventrally, the premaxilla rostrally, and the prefrontal dorsally. The lacrimal is relatively smaller at early ontogenetic stages (CMN 2246 and CMN 2247) than in the approximately equivalent juvenile stages of other Corythosaurus and Lambeosaurus species. Ventrally, the lacrimal abuts the jugal, but does not overlap it laterally as in P. walkeri .
Prefrontal
The prefrontal is similar in all significant aspects to that of other corythosaurins ( Figs 2 View Figure 2 , 3 View Figure 3 , 10 View Figure 10 , 11 View Figure 11 ). An interdigitating joint with the postorbital along the dorsal margin of the orbit excludes the frontal from reaching the orbital margin. The medial flange undergoes two significant changes through ontogeny, verticalization and caudal development, both of which are associated with the development of the cranial crest. The prefrontal flange is small and medially directed in CMN 2247. As it develops it becomes a vertical sheet, and is retracted over the dorsal surface of the frontal ( Figs 10A View Figure 10 , 11 View Figure 11 ). The flange has a significant medial deflection in CMN 2247 and CMN 2246, but is vertical in TMP 82.10.01 and adult-sized skulls. Despite the size-related shape change, the medial flange retains contact with the nasal throughout ontogeny. Contact between the lacrimal occurs along the rostral margin of the orbit, where the prefrontal overlaps the lacrimal dorsolaterally, and extends ventrally to the top of the lacrimal duct medially.
Postorbital
The postorbital is almost identical to that of other corythosaurins, except that it has a distinctly shaped caudal process. The caudal margin of the process is sinuous in lateral view (CMN 2246; Fig. 3 View Figure 3 ), and gently convex in all larger specimens (CMN 8501 and ROM 702; Fig. 11 View Figure 11 ). The distal end of the caudal process is never embayed, or bifurcated, as in all other hadrosaurids (including H. stebingeri ; MOR 549) and at least some non-hadrosaurid hadrosauroids.
Frontal
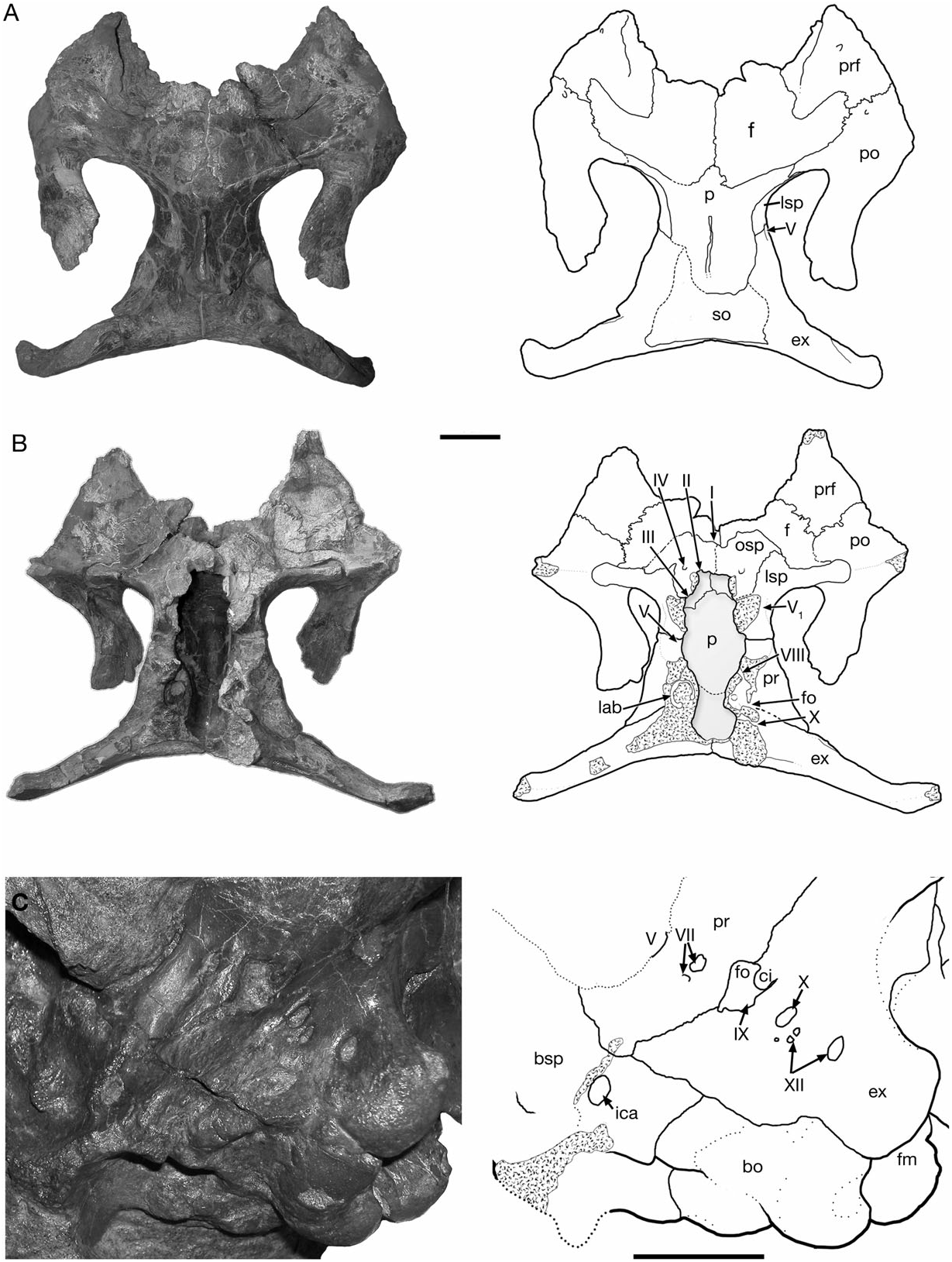
The frontal does not differ significantly from that of other corythosaurins, such as Corythosaurus and Lambeosaurus . In dorsal view, the frontal contacts the nasal rostrally, the prefrontal and the postorbital laterally, and the parietal caudally ( Figs 2 View Figure 2 , 3 View Figure 3 , 10 View Figure 10 , 11 View Figure 11 ). Ventrally, it is firmly united with the presphenoid, orbitosphenoid and laterosphenoid to enclose the telencephalon. The frontals unite along the midline of the skull roof to form a median dome that is more prominent in juveniles (CMN 2247) than in large specimens. The relative length of the externally exposed region between the nasal and parietal shows strong negative allometry associated with the ontogenetic development of the cranial crest. In large individuals the relative length of the frontal differs between taxa, and may provide character data important in the resolution of lower-level lambeosaurine phylogeny ( Godefroit et al., 2004a, b; Evans & Reisz, 2007). The two complete adult-sized braincases available for H. altispinus show that the interfrontal suture length: frontal width ratio ranges from 0.46 (CMN 8675) to 0.49 (ROM 702), whereas in the juvenile specimen CMN 2246, the length of the interfrontal joint ( 33.5 mm) is 75% of the maximum frontal width ( 87.3 mm).
Parietal
The unpaired parietal forms the medial and rostromedial margins of the supratemporal fenestrae. It contacts the frontal and postorbital rostrally, the squamosal caudally, and the laterosphenoid, the prootic, and the supraoccipital ventrally. Rostrally, a small interfrontal process of variable size is always present. The midline sagittal crest that separates the supratemporal fenestra is relatively weakly developed in CMN 2246, and it deepens throughout ontogeny. Ventrally, the parietal forms a large dorsal portion of the endocranial cavity, as observed in AMNH 5248 ( Fig. 10 View Figure 10 ). The interfrontal process, much larger endocranially than dorsally exposed, contributes to the caudomedial region of the cerebral fossa. Caudally, the ovate cerebellar fossa forms a deep pit for the metencephalon rostral to the supraoccipital. The parietal is more rostrocaudally compact than in Lambeosaurus , Corythosaurus , and hadrosaurines. In Corythosaurus and Lambeosaurus , the parietal is over 50% the interorbital width of the skull. In ROM 702, the length of the parietal ( 80 mm) is less than 40% of the interorbital width ( 217 mm). Hypacrosaurus stebingeri and P. walkeri have a similarly shortened parietal (see below).


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Hypacrosaurus altispinus
| Evans, David C. 2010 |
Hypacrosaurus altispinus
| Gilmore CW 1924: 50 |
Cheneosaurus tolmanensis
| Lambe LM 1917: 18 |
Hypacrosaurus altispinus
| Brown B 1913: 395 |