
Trichocyclus arawari, HUBER, 2001
|
publication ID |
https://doi.org/10.1206/0003-0090(2001)260<0001:TPOAAP>2.0.CO;2 |
|
persistent identifier |
https://treatment.plazi.org/id/5F4E362A-FFCD-5C15-7847-FCCEFECBFF97 |
|
treatment provided by |
Marcus |
|
scientific name |
Trichocyclus arawari |
| status |
sp. nov. |
Trichocyclus arawari View in CoL , new species Figures 259 View Figs –266
TYPE: Male holotype from ThreeMile Valley , Wyndham (~15°30̍S, 126°05̍E), Western Australia, Australia ; June 30, 1981 (D. Hirst), in SAM (N1999/872).
ETYMOLOGY: Named for the Ngarinjin (also called Arawari), an aboriginal tribe in Western Australia. The species name is a noun in apposition.
DIAGNOSIS: Distinguished from the very similar T. worora by the shape of the procursus (compare figs. 265 and 268) and by the shape of the male chelicerae that lack the strong median projection (compare figs. 264 and 267); distinguished from all other known congeners also by the high elevations on the female carapace (fig. 261).
MALE ( holotype): Total length 4.1, carapace width 1.58. Leg 1: 36.9 (9.9 + 0.7 + 9.7 + 14.1 + 2.5), tibia 2: 6.9, tibia 3: 4.7, tibia 4: 6.7; tibia 1 l/d: 63. Prosoma shape and pattern as in T. nullarbor (cf. figs. 186– 189), but without radial marks; ocular area posteriorly with brown median band. Distance PMEPME 0.145; diameter PME 0.120; distance PMEALE 0.045; diameter AME 0.125. Clypeus with brown mark not reaching eyes; sternum brown with yellowish speckles. Chelicerae as in fig. 264; brown, with only one pair of simple apophyses. Palps as in figs. 259–260; procursus as in fig. 265. Legs ochre, with indistinct darker rings on femora (subdistally) and tibiae (proximally and subdistally); patellae also darker; most hairs on legs missing; retrolateral trichobothrium of tibia 1 at 14%; tarsus 1 distally with ~11 distinct pseudosegments, proximally the pseudosegmentation is very indistinct. Op isthosoma dorsoposteriorly very long, gray, covered with black spots except ventrally; genital plate light brown, about rectangular.
VARIATION: Tibia 1 in other males: 11.1, 11.7, 13.6, 13.7; in the males from Tunnel Creek National Park the prolaterodorsal
Figs. 264–268. Trichocyclus arawari (264–266), and T. worora (267, 268), males. 264, 266, 267. Chelicerae, frontal views (264: male from ThreeMile Valley, Wyndham; 266: male from Napier Range). 265, 268. Left cymbia and procursi, retrolaterodorsal views; asterisk: weak zone on cymbium; arrows: prolaterodorsal apophysis. Scale lines: 0.3 mm (264–268).
apophysis on the procursus (arrow in fig. 265) is simpler, more like that in T. worora (cf. fig. 268); the males from Geikie Range and Napier Range have a posteriorly rounded opisthosoma and lack both radial and lateral marks on the carapace; the males from Napier Range differ significantly with respect to the chelicerae (fig. 266), but have apparently almost identical palpal structures. The specimens from Napier Range are therefore assigned tentatively.
FEMALE: In general very similar to male, but carapace with pair of conspicuous elevations (fig. 261). Opisthosoma frontodorsally with pair of indistinct, transparent humps that oppose elevations on carapace. Tibia 1 in 3 females: 10.1, 11.2, 11.3. Epigynum as in fig. 262; dorsal view as in fig. 263.
DISTRIBUTION: Known from several localities in northern Western Australia (map 13).
MATERIAL EXAMINED: AUSTRALIA: Western Australia: ThreeMile Valley, Wyndham : Male holotype above, with 13 2♀ ( SAM N1999 View Materials /873–5) ; Napier Range , cave KN90 (17°14̍S, 124°39̍E), July 24, 1998 ( S. M. Eberhard), 23 1♀ 1 juvenile ( WAM 99 About WAM / 2481–4), assigned tentatively ; Tunnel Creek National Park (17°37̍S, 125°10̍E), June 7, 1999 ( M. Gray, G. Milledge, H. Smith), in webs, cave wall, 23 1♀ ( AMS KS56188 ) ; Geikie Range , cave KG47 (18°02̍S, 125°44̍E), July 1, 1998 ( S. M. Eberhard), 13 ( WAM 99 About WAM /2473) .
Trichocyclus worora , new species Figures 267, 268
TYPE: Male holotype from Kandiwal ( Camp Creek) (14°52̍S, 125°42̍E), Mitchell Plateau, Western Australia, Australia ; Dec. 11, 1993 (A. F. Longbottom), in WAM (99/ 1759).
ETYMOLOGY: Named for the Worora, an aboriginal tribe in Kimberley, Western Australia. The species name is a noun in apposition.
DIAGNOSIS: Distinguished from the very similar T. arawari by the shape of the procursus (compare figs. 265 and 268) and by the male chelicerae with their pair of strong median projections (compare figs. 264 and 267); from all other known congeners also by the high elevations on the female carapace (cf. fig. 261).
MALE ( holotype): Total length 3.3, carapace width 1.52. Leg 1: 33.9 (9.2 + 0.7 + 9.0 + 13.3 + 1.7), tibia 2: 6.4, tibia 3: 4.4, tibia 4: 6.3; tibia 1 l/d: 64. Prosoma shape and pattern as in T. nullarbor (cf. figs. 186– 189), but without radial marks; ocular area and clypeus dark brown; sternum almost black. Distance PMEPME 0.145; diameter PME 0.120; distance PMEALE 0.045; diameter AME 0.135. Chelicerae as in fig. 267, dark brown, with pair of simple distal apophyses, pair of strong median projections, and low lateral humps, all black. Palps in general as in T. arawari (cf. figs. 259, 260), procursus as in fig. 268. Legs light brown, with black rings on femora (subdistally) and tibiae (proximally and subdistally); patellae also darker, tips of femora and tibiae whitish; without spines, without curved and vertical hairs; retrolateral trichobothrium of tibia 1 at 11%; pseudosegmentation of tarsi hardly visible. Opisthosoma posteriorly more pointed than in T. nullarbor ; gray, with black spots except ventrally; genital plate dark brown.
VARIATION: Tibia 1 in male from Cockatoo Island: 8.2. This male has a slightly more pointed prolaterodorsal apophysis on the procursus (arrow in fig. 268), and the median projection on the chelicerae is longer.
FEMALE: In general very similar to male, but carapace with pair of conspicuous elevations (cf. fig. 261), and opisthosoma dorsoposteriorly much longer. Opisthosoma frontodorsally with pair of fairly distinct, transparent humps opposing elevations on carapace. Tibia 1 in 2 females: 6.5, 8.0. Epigynum ventrally as in T. arawari (cf. fig. 262).
DISTRIBUTION: Known from two localities in northern Western Australia (map 13). Note, however, that the locality Cockatoo Island is not certain.
MATERIAL EXAMINED: AUSTRALIA: Western Australia: Kandiwal ( Camp Creek): Male holotype above, with 2♀ 1 juvenile ( WAM 99 About WAM /1759 –63) ;? Cockatoo Island (16°06̍S, 123°37̍E), Oct. 30, 1961 ( A. R. Main), 13 ( WAM 99 About WAM /1693) .
Trichocyclus arnga , new species Figures 269–272
TYPE: Male holotype from Jeremiah Hills (15°26̍S, 128°44̍E), Kimberley, Western Australia, Australia ; May 4, 1994 (R. D. Brooks), cave KJ8, in WAM (99/2047).
ETYMOLOGY: Named for the Arnga, an aboriginal tribe in Kimberley, Western Australia. The species name is a noun in apposition.
DIAGNOSIS: Easily distinguished from known congeners by the huge median apophyses on the male chelicerae (figs. 269, 270).
MALE ( holotype): Total length 3.9, carapace width 1.65. Leg 1: 41.7 (11.3 + 0.7 + 11.5 + 16.1 + 2.1), tibia 2: 8.3, tibia 3: 5.4, tibia 4: 7.6; tibia 1 l/d: 78. Prosoma shape similar to T. nullarbor (cf. figs. 186–189); carapace ochre with dark median band and wider spot behind ocular area, without lateral marks. Ocular area dark brown; distance PMEPME 0.175; diameter PME 0.120; distance PMEALE 0.040; diameter AME 0.120. Clypeus with large brown spot; sternum ochre with yellowish speckles. Chelicerae ochre with two very distinctive pairs of apophyses, as well as stridulatory ridges (figs. 269, 270). Palps in general as in T. nullarbor (cf. figs. 191, 192), with distinctive procursus (similar only to T. bugai ) and bulb (figs. 271, 272). Legs ochre, with darker rings on femora (subdistally) and patellae + tibiae proximally; legs without spines, without curved and vertical hairs; retrolateral trichobothrium of tibia 1 at 7%; tarsus 1 distally with ~20 quite distinct pseudosegments, proximally pseudosegmentation not visible in dissecting microscope. Opisthosoma roundish, but higher than long (2.7/2.4); gray, with blackish spots except ventrally; genital plate not darker than surrounding area.
VARIATION: The other known male (see below) is much smaller (carapace width: 1.00; tibia 1: 5.6), but the shapes of chelicerae and palpal structures appear identical.
FEMALE: Unknown.
DISTRIBUTION: Known from two localities in Kimberley, Western Australia (map 13).
MATERIAL EXAMINED: AUSTRALIA: Western Australia: Jeremiah Hills: Male holotype above ; Great Northern Highway , 52 km N of Turkey Creek (16°38̍S, 128°12̍E), June 7, 1999 ( M. Gray, G. Milledge, H. Smith), in webs, cave wall, 13 ( AMS KS56182 ) .
Trichocyclus bugai , new species Figures 273, 274
TYPE: Male holotype from Drysdale River Station airfield (15°43̍S, 126°23̍E), under airfield runway marker, Kimberley, Western Australia, Australia ; March 10, 1994 (A. F. Longbottom), in WAM (99/1750).
ETYMOLOGY: Named for the Bugai, aboriginal people of the Wenambal tribe in Kimberley, Western Australia. The species name is a noun in apposition.
DIAGNOSIS: Distinguished from known congeners by the single pair of cheliceral apophyses that are divided into two lobes each (fig. 274), and by the shape of the procursus (fig. 273; similar only to T. arnga , cf. fig. 271).
MALE ( holotype): Total length 2.4, carapace width 1.03. Leg 1: 18.7 (5.2 + 0.4 + 5.1 + 6.8 + 1.2), tibia 2: 3.5, tibia 3: 2.4, tibia 4: 3.5; tibia 1 l/d: 55. Prosoma shape similar to T. nullarbor (cf. figs. 186–189); carapace ochre with pattern as in T. nullarbor (cf. fig. 187). Ocular area with darker median and lateral marks; distance PME PME 0.120; diameter PME 0.080; distance PMEALE 0.040; diameter AME 0.105. Clypeus with wide dark band tapering distally; sternum ochre, medially with light brown speckles. Chelicerae with only one pair of bilobed apophyses, as well as stridulatory ridges (fig. 274). Palps in general as in T. arawari (cf. figs. 259, 260, rather than
Figs. 269–274. Trichocyclus arnga (269–272), and T. bugai (273, 274), males. 269, 270, 274. Chelicerae, frontal and lateral views. 271, 273. Left procursi and genital bulbs, retrolateral views; ‘‘a’’, ‘‘p’’ = apophysis and pocket. 272. Left genital bulb, prolateral view. Scale lines: 0.3 mm.
T. nullarbor !), with distinctive procursus (similar only to T. arnga ) and bulb (fig. 273). Legs ochre, with darker rings on femora (subdistally), patellae + tibiae proximally, and tibiae distally; tips of femora and tibiae whitish; legs without spines, curved, and vertical hairs; retrolateral trichobothrium of tibia 1 at 16%; tarsus 1 distally with ~15 very distinct pseudosegments, proximally pseudosegmentation not visible in dissecting microscope. Opisthosoma probably roundish (damaged); gray, with blackish spots except ventrally; genital plate dark brown, rectangular.
FEMALE: In general very similar to male, but sternum with wide brown median band; tibia 1: 4.7. Opisthosoma frontodorsally without humps. Epigynum in general similar to T. arabana (cf. fig. 213) but with pair of distinctive, round invaginations near raised median part of frontal plate.
DISTRIBUTION: Known only from type locality in Kimberley, Western Australia (map 13).
MATERIAL EXAMINED: AUSTRALIA: Western Australia: Kimberley: Drysdale River Station airfield: Male holotype above ; same collection data, 1♀ ( WAM 99 About WAM /1749) .
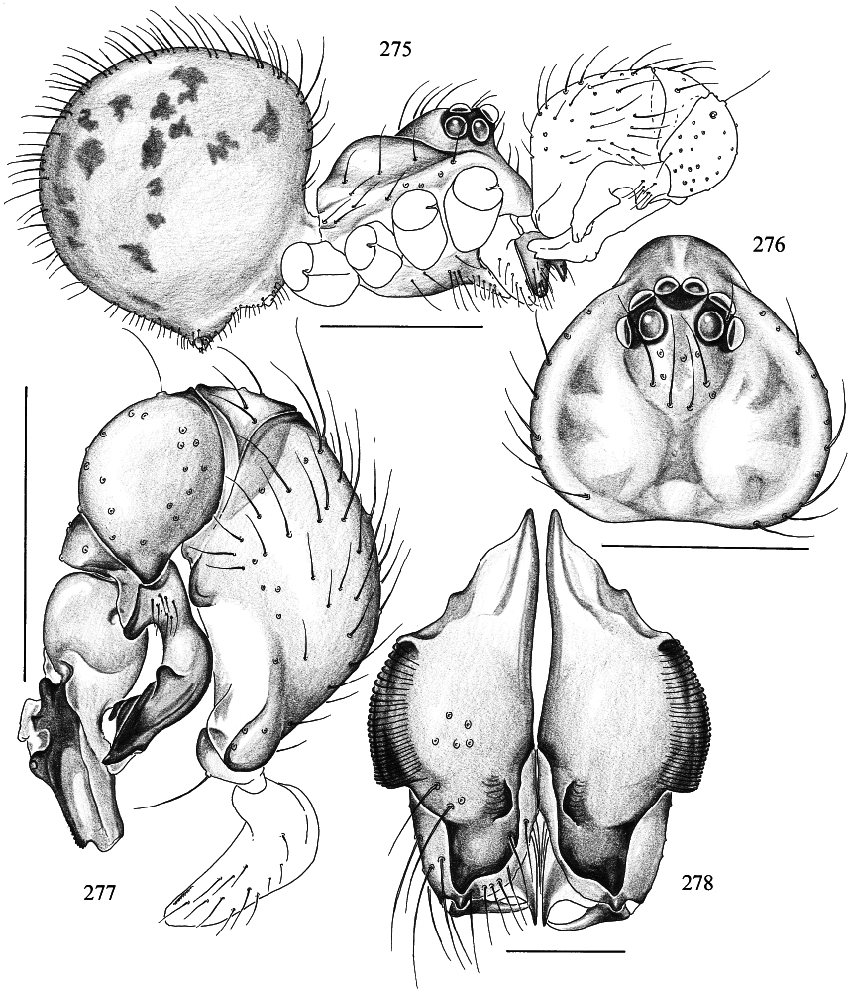
Trichocyclus harveyi , new species Figures 275–282 View Figs View Figs
TYPE: Male holotype from N of Larryoo (14°51̍S, 126°49̍E), Kimberley , Western Australia, Australia ; June 12, 1992 (M. S. Harvey, J. M. Waldock), under rock, in WAM (99/1654).
ETYMOLOGY: Named for the first collector of the present material and of many more pholcids in the Western Australian Museum.
DIAGNOSIS: Small species, distinguished from known congeners by the shape of the procursus (figs. 277, 280) and bulb (figs. 277, 279), and by its small size and short legs (the legs of T. ungumi , the second smallest known species, are more than twice as long).
MALE ( holotype): Total length 1.4, carapace width 0.74. Leg 1: 8.07 (2.16 + 0.26 + 2.26 + 2.68 + 0.71), tibia 2: 1.52, tibia 3: 1.03, tibia 4: 1.65; tibia 1 l/d: 34. Habitus and prosoma shape as in figs. 275 and 276; carapace pale ochre with slightly darker spots medially and laterally. Distance PME PME 0.075; diameter PME 0.080; distance PMEALE 0.025; diameter AME 0.055. Clypeus slightly darker than carapace; sternum pale ochre, shape as in T. nullarbor (cf. fig. 189). Chelicerae pale ochre with two pairs of frontal apophyses, and stridulatory ridges (fig. 278). Palps as in fig. 277; femur ventrally with distinct brown hump (fig. 277); procursus as in fig. 280, bulb as in fig. 279. Legs ochregray, without rings; without spines, without curved and vertical hairs; retrolateral trichobothrium of tibia 1 at 30%; tarsus 1 distally with ~7 fairly visible pseudosegments, proximally pseudosegmentation not visible in dissecting microscope. Opisthosoma roundish (fig. 275), pale ochregray, with large blackish spots except ventrally; genital plate not darker than surrounding area.
FEMALE: In general very similar to male; tibia 1 in 2 females: 1.97, 2.03; carapace width in 2 females: 0.73, 0.77. Opisthosoma frontodorsally without humps. Epigynum as in fig. 281; dorsal view as in fig. 282.
DISTRIBUTION: Known only from type locality in Kimberley, Western Australia (map 14).
MATERIAL EXAMINED: AUSTRALIA: Western Australia: Kimberley, N of Larryoo: Male holotype above, with 2♀ 1 juvenile ( WAM 99/1655–7).
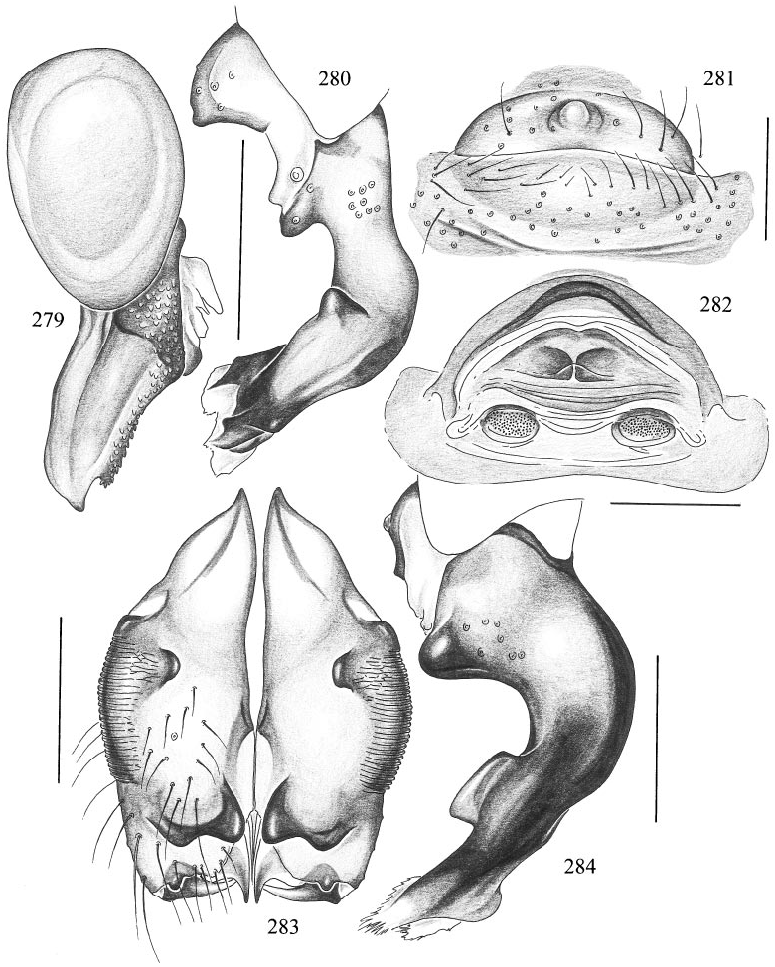
Trichocyclus ungumi , new species Figures 283, 284 View Figs
TYPE: Male holotype from 30 miles E of Derby (17°19̍S, 124°05̍E), Kimberley , Western Australia, Australia ; June 8, 1970 (Hemley Exped.), in WAM (99/1719).
ETYMOLOGY: Named for the Ongkomi (also called Ungumi), an aboriginal tribe from Kimberley, Western Australia. The species name is a noun in apposition.
DIAGNOSIS: Small species, distinguished from congeners by the twolobed distal male cheliceral apophyses and the position of the proximal apophyses (very proximal: fig. 283), and by the shape of the procursus (slightly similar to T. aranda , compare figs. 232 and 284).
MALE ( holotype): Total length 1.6, carapace width 0.97. Leg 1: 19.2 (5.2 + 0.4 + 5.2 + 7.2 + 1.2), tibia 2: 3.5, tibia 3: 2.5, tibia 4: 3.5; tibia 1 l/d: 52. Habitus and prosoma shape as in T. nullarbor (cf. figs. 186– 189), but much smaller; carapace ochreyellow with brown mark behind ocular area, black in thoracic groove, two pairs of lateral brown marks posteriorly, without radial marks. Ocular area with median dark band posteriorly. Distance PMEPME 0.095; diameter PME 0.085; distance PMEALE 0.040; diameter AME 0.095. Clypeus with pair of brown bands under ALE, distally converging into large spot; sternum medially light brown with yellowish spots, laterally ochreyellow. Chelicerae ochre with pair of twolobed distal apophyses and another pair very proximally, and stridulatory ridges (fig. 283). Palps in general as in T. harveyi (cf. fig. 277), femur medioventrally with distinct brown knob, bulb similar to that of T. aranda (cf. fig. 231), procursus as in fig. 284. Legs ochreyellow, with darker rings on femora (subdistally) and tibiae (proximally and subdistally); patellae also darker; tips of femora and tibiae whitish; most hairs on legs missing; retrolateral trichobothrium of tibia 1 at 14%; tarsus 1 distally with ~7 barely visible pseudosegments, proximally pseudosegmentation not visible in dissecting microscope. Opisthosoma roundish, only slightly higher than long, ochregray, with many blackish spots except ventrally; genital plate relatively large, brown, trapezoidal.
FEMALE: In general very similar to male, with three pairs of lateral spots on carapace; tibia 1: 4.5. Opisthosoma frontodorsally without humps. Epigynum in ventral view as in T. aranda (cf. fig. 233).
DISTRIBUTION: Known only from type lo cality in Kimberley, Western Australia (map 14).
MATERIAL EXAMINED: AUSTRALIA: Western Australia: Kimberley, 30 miles E of Derby: Male holotype above, with 1♀ ( WAM 99/1720).
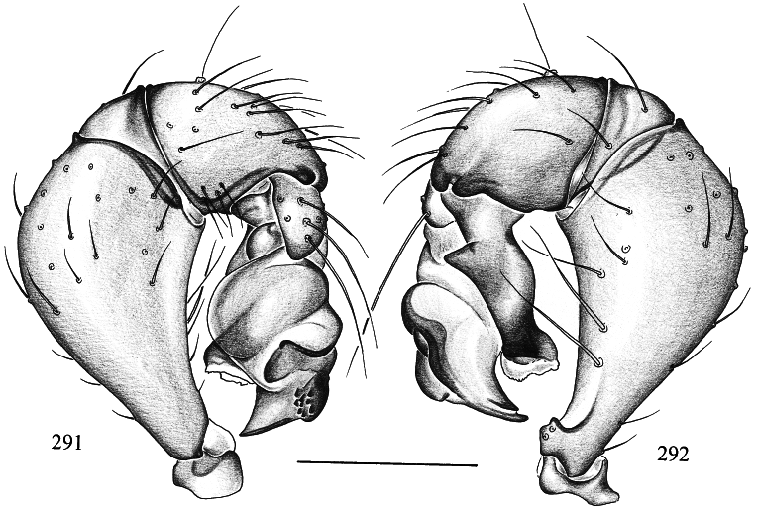
Trichocyclus watta , new species Figures 285–292 View Figs View Figs
TYPE: Male holotype from Gorge NE of Mt. Gilruth (13°02̍S, 133°05̍E), Northern Territory, Australia ; July 12, 1979 ( G. B. Monteith), rainforest, sieved litter, in QMB (S50178) .
ETYMOLOGY: Named for the Watta, an aboriginal tribe in the area of the Alligator Rivers, Northern Territory. The species name is a noun in apposition.
DIAGNOSIS: Tiny species, easily distinguished from all congeners and other possible relatives ( Wugigarra nauo , W. kalamai ) by the absence of AME. Also distinguished by the row of apophyses on the male chelicerae (fig. 288), and by the very short legs.
MALE ( holotype): Total length 1.2, carapace width 0.51. Leg 1: 3.50 (1.00 + 0.19 + 0.95 + 0.97 + 0.39), tibia 2: 0.84, tibia 3: 0.63, tibia 4: 0.95; tibia 1 l/d: 16. Habitus and prosoma shape as in figs. 285–287; entire prosoma monochromous ochre. Ocular area only slightly elevated (fig. 285); distance PMEPME 0.055; diameter PME 0.055; distance PMEALE 0.015; AME missing. Sternum wide, similar to T. nullarbor (cf. fig. 189). Chelicerae light brown, with row of apophyses on each side (fig. 288). Palps as in figs. 291 and 292, with weak zone dorsally on cymbium. Legs monochromous ochre; without spines, without curved and vertical hairs; retrolateral trichobothrium of tibia 1 apparently at 58% (difficult to see); tarsus 1 with ~5–7 pseudosegments (difficult to count). Opisthosoma oval, monochromous gray.
FEMALE: Very similar to male. Chelicerae apparently without stridulatory files. Epigynum as in fig. 289; dorsal view as in fig. 290.
DISTRIBUTION: Known from two localities in Northern Territory (map 14).
MATERIAL EXAMINED: AUSTRALIA: Northern Territory: NE of Mt. Gilruth: Male holotype above, with 1♀ 4 juveniles in same vial ; same locality and collector, July 10, 1979: 1♀ 1 juvenile ( QMB S50176 View Materials ) ; Kakadu National Park, Mirrai ( Mt. Cahill ) (12°53̍S, 132°42̍E), July 5–7, 1994 ( M. S. Harvey, D. M. Hyder), under stones, 1♀ ( WAM 99 About WAM / 1511), assigned tentatively .
Micromerys Bradley, 1877: 119 ( type species by monotypy M. gracilis Bradley, 1877 , examined).—Simon, 1893: 474.—DeelemanReinhold, 1986b: 205–224.—Huber, 1997c: 358.
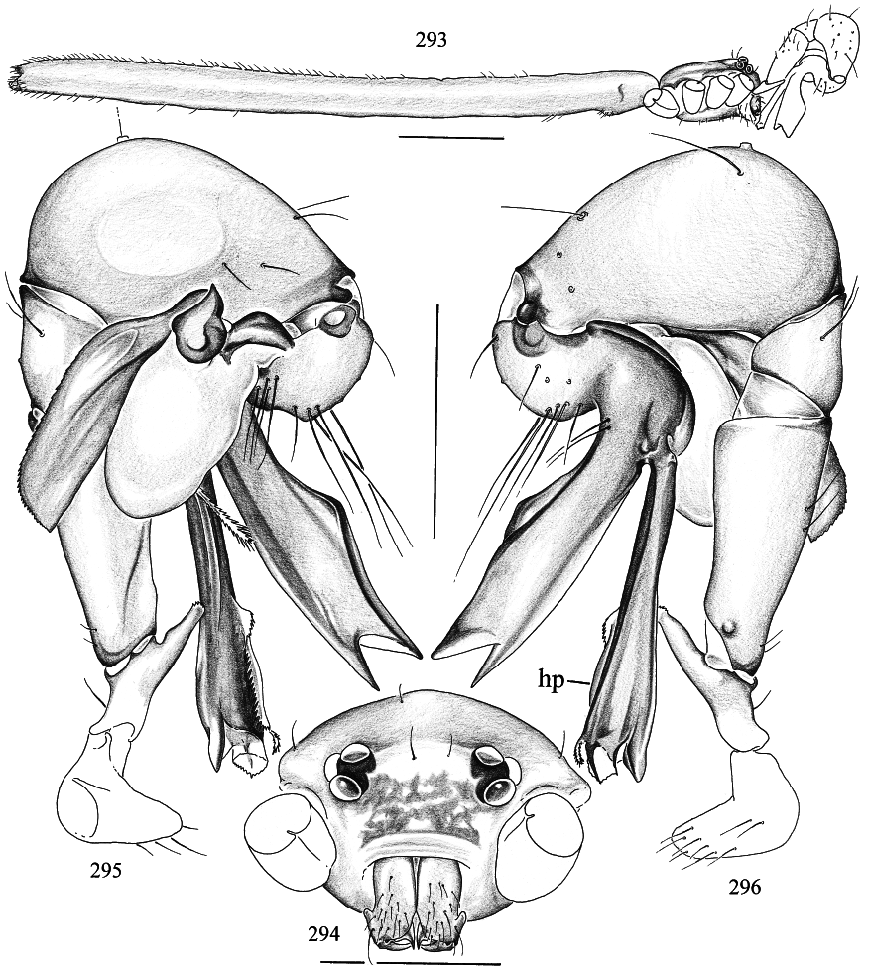
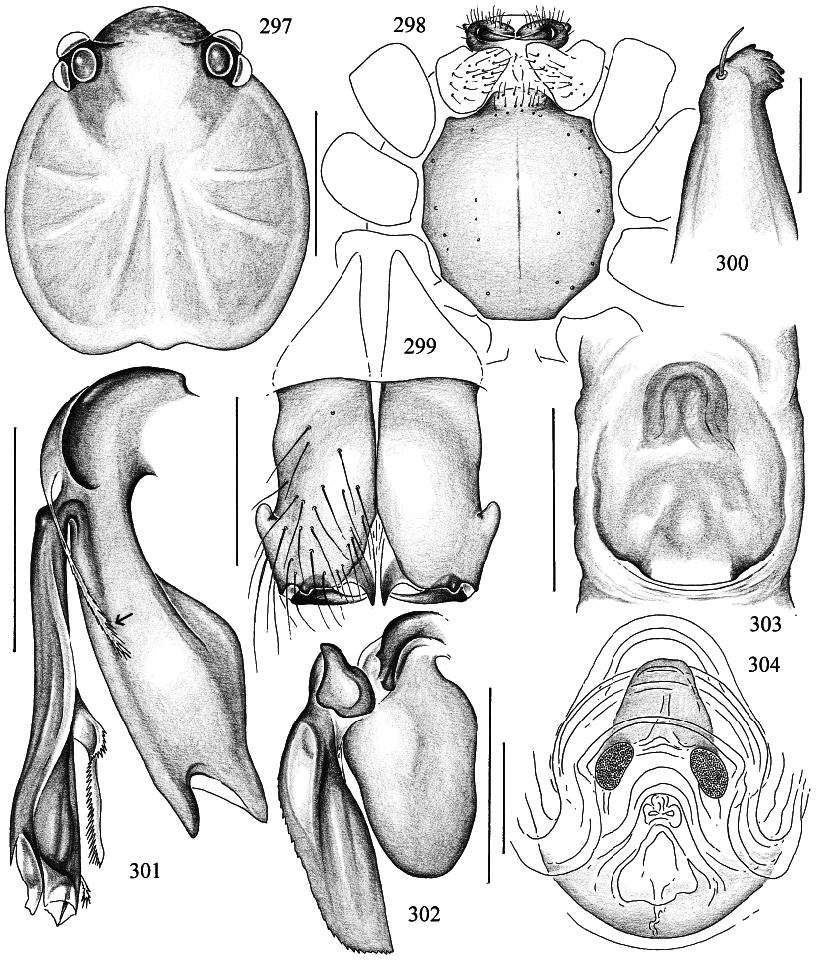
DIAGNOSIS: Pale longlegged pholcids with small flat prosoma and very long cylindrical opisthosoma (fig. 293); total length ~ 6–8 mm; apparently restricted to the Australian region. Six eyes, AME either completely missing or reduced to pigment specks. Distinguished from Panjange (which is the only similar genus in Australia) by the single sclerotized process on the bulb (e.g., figs. 302, 313, 316), the hinged process ventrally on the procursus (‘‘hp’’ in figs. 296, 318), the simple epigynum (e.g., figs. 303, 337; in Panjange , the epigynum has a long scape), the absence of eye turrets in the male, and the cylindrical opisthosoma. Distinguished from Leptopholcus , Calapnita , and probably also from Uthina (of which only females have been described) by the single bulbal projection and the hinged process on the procursus, and by the absence of frontal apophyses on the male chelicerae (fig. 299).
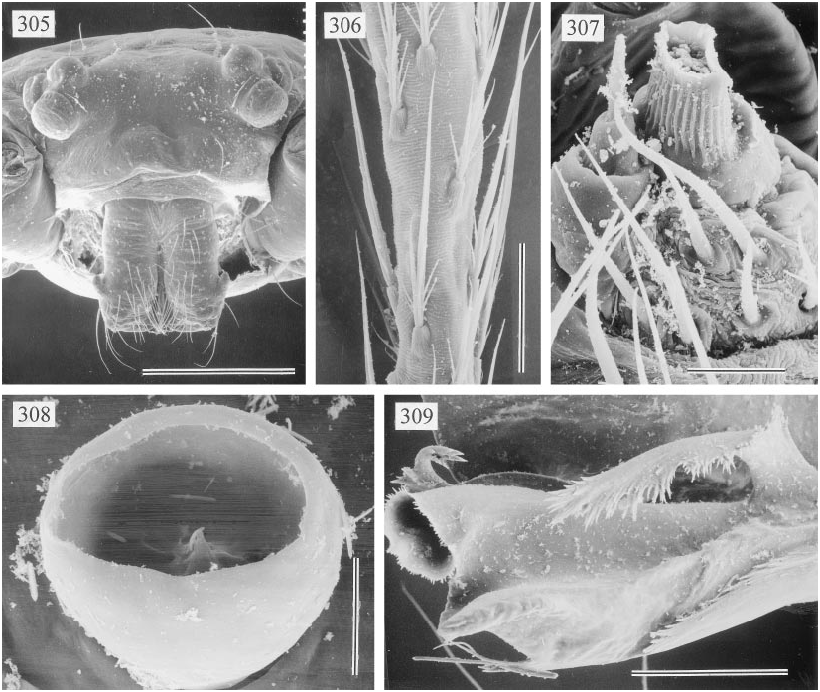
DESCRIPTION: Total length in males ~ 6–8 mm. Carapace roundish to slightly longer than wide, flat and without markings. Six eyes in two triads, sometimes slightly raised; AME missing or reduced to pigment specks (e.g., fig. 310). Distance PMEALE small (~25–55% of PME diameter). Clypeus never modified, in some species very short compared to all other pholcids (figs. 294, 305, 310). Male chelicerae extremely conservative, with pair of lateral apophyses, without frontal apophyses (fig. 299), without stridulatory ridges. Male palps large in relation to prosoma size (figs. 293); coxa without retrolateral apophysis, trochanter with long ventral apophysis that is distally provided with distinct ridges (figs. 300, 333); femur almost cylindrical, with small retrolateral hump proximally, with ventrodistal process in some species (fig. 314); patella triangular in lateral view; tibia considerably enlarged, with 2 trichobothria (fig. 296); cymbium and procursus characteristically Sshaped (e.g., figs. 296, 312, 318; following dorsal contour); procursus consisting of main branch, complex hinged process ventrally (e.g., figs. 301, 312, 318), and translucent fringed projection arising near base of hinged process (arrows in figs. 301, 317, 320); bulb consisting basically of globular part and flat sclerotized process (‘‘p’’ in fig. 313); globular part of bulb connected to cymbium by arched sclerotized structure (‘‘c’’ in fig. 313), to flat pro cess by triangular element (‘‘t’’ in fig. 313). (I have not been able to find the sperm duct opening; the sperm duct may or may not enter the flat sclerotized process.) Tarsal organ capsulate (figs. 308, 330). Legs long and thin (leg 1 ~4–5 X body length, ~27–35 X prosoma length; tibia 1 l/d ~75–85); leg formula 1423, leg 4 significantly longer than leg 2; with dark ring in patella area and dark tibiametatarsus joint; legs without spines, without vertical and curved hairs; retrolateral trichobothrium of tibia 1 at 2–4%; tarsi without pseudosegmentation but with unique striation of cuticle (figs. 306, 332); tarsal claws as usual for family (see Note below). Opisthosoma very long and cylindrical, spinnerets at the tip. Male gonopore with four epiandrous spigots (examined: M. daviesae , gracilis ; fig. 307). ALS with only one pair of spigots each (examined: M. daviesae , gracilis ; fig. 329); other spinnerets typical for family.
Sexual dimorphism slight, females with shorter legs, often with longer opisthosoma, eye triads less raised. Epigynum shape extremely simple and conservative; internally more diverse and complex (e.g., figs. 304, 315, 328), but mostly transparent and difficult to study.
NOTE: DeelemanReinhold (1986b) mentioned a difference between the claws on tarsi 1–3 versus tarsus 4. A detailed reexamination of M. gracilis revealed no difference between the claws on tarsi 1 and 4, except that the claws on tarsus 4 were slightly longer (32 versus 27 µm).
MONOPHYLY: All described species share the bulbal morphology (flat sclerotized process, triangular element) and the single pair of lateral apophyses on the male chelicerae. An undescribed species from Papua New Guinea (see below) has the ‘‘triangular’’ element slightly more elaborate (longer, distally divided into two parts), but it is otherwise very similar and clearly congeneric. The files on the tip of the male palpal trochanter apophysis and the transparent projection arising from near the basis of the hinged process might be further synapomorphies of the genus.
GENERIC RELATIONSHIPS: Two characters support the inclusion of Micromerys in the Pholcus group sensu Huber, 1995: the prox imolateral male cheliceral apophyses (functionally related to the ventral trochanter apophysis); and the spindleshaped male palpal tibia. This was supported by cladistic analyses using equally weighted characters, as well as by using successive weighting in Hennig86. However, implied weighting in PeeWee suggested an alternative scenario at all settings of the concavity constant except at conc = 6: the South American Metagonia was sister group of Micromerys , supported by the ventral hinged process on the procursus. (Both genera were nested in a ‘‘ Pholcus group sensu lato’’.)
NATURAL HISTORY: The genus seems to be restricted to humid forests, where the spiders live mainly on the underside of leaves (as judged from a few labels giving relevant information; see also DeelemanReinhold and Deeleman, 1983, DeelemanReinhold, 1986a, b). No singlespecies study, or any closer observation, is known to me.
DISTRIBUTION: Known from Australia and (one undescribed species in the CLD collection) from Papua New Guinea (Deeleman Reinhold [1986a: fig. 2; 1986b: 206] report ed ‘‘an unedited species from West Irian’ ’, which I have not seen). Within Australia, the genus is restricted to northern Northern Territory and to the Great Dividing Range in Queensland and New South Wales. It is remarkable that four of the seven known species occur within a limited area around Cairns .
COMPOSITION: The genus now includes 7 described species, 5 of them new. All described species are treated below. The collections studied contain numerous further vials with females only, which does not allow useful guesses about possible undescribed species. For species previously assigned to Micromerys , see Millot (1946), Brignoli (1980), DeelemanReinhold (1986a), and Huber (1997c, 2000).
Micromerys gracilis Bradley, 1877 Figures 293–309 View Figs View Figs View Figs
Micromerys gracilis Bradley, 1877: 119 .—Simon, 1893: 474.—DeelemanReinhold, 1986b: 206– 209, figs. 5–9, 60 (female only; male see M. yidin below).
TYPE: Female neotype designated by Dee lemanReinhold (1986b), from Lake Boronto near Somerset, 5 km S of Cape York (10°45̍S, 142°35̍E), Queensland, Australia ; Feb. 3–4, 1975 (R. Raven), in QMB (S885), examined.
REMARK: The selection of a female as type poses some problems as females of Micromerys are extremely difficult to identify. However, M. gracilis seems to be the only species on Cape York Peninsula (map 15; note that the locality Shiptons Flat in DeelemanReinhold [1986b] rests on a misidentification; see M. yidin below), suggesting that the material studied is indeed conspecific with the neotype.
DIAGNOSIS: Distinguished from congeners by the wide main branch of the procursus (fig. 301) and by the straight bulbal apophysis that is slightly longer than the bulb (figs. 295, 302).
MALE (Murray Island): Total length 5.8, carapace width 0.84. Leg 1: 27.6 (7.3 + 0.4 + 6.3 + 10.9 + 2.7), tibia 2: 5.1, tibia 3: 3.2, tibia 4: 5.9; tibia 1 l/d: 79. Habitus and prosoma shape as figs. 293, 294, 297, 298. Entire prosoma whitish ochre, only proximal part of clypeus light brown (fig. 294). Ocular area flat; distance PMEPME 0.280; diameter PME 0.105; distance PMEALE 0.050; AME missing, only tiny black spot between triads. Sternum slightly longer than wide (fig. 298). Chelicerae whitish, without modification except pair of lateral apophyses (fig. 299). Palps as in figs. 295 and 296, pale ochre, only procursus and bulbal apophysis light to dark brown; trochanter apophysis, procursus, and bulb as in figs. 300–302; tip of hinged process as in fig. 309; palpal tarsal organ as in fig. 308. Legs pale ochre; patellae, femora tips, and tibia–metatarsus joints darker; without spines, without curved and vertical hairs; retrolateral trichobothrium of tibia 1 at 2.3%; tarsi without pseudosegmentation (fig. 306). Opisthosoma shape as in fig. 293, monochromous pale ochre; two spigots on ALS (fig. 307).
VARIATION: Tibia 1 in 14 males: 6.1–7.4 (x = 6.7). Some males have no trace of AME, others have black spots in that area, but lenses are always missing. Some males have darker smudges on the opisthosoma.
FEMALE: In general very similar to male, but clypeus proximally not darker. Tibia 1 in 9 females: 5.2–6.2 (x = 5.8). Tibia 1 in neotype: 5.5. Variation in AME spots as in male ( neotype without spots). Epigynum very simple, unsclerotized, with possibly distinctive arch shining through (fig. 303); dorsal view as in fig. 304.
DISTRIBUTION: Known from several localities in northern Northern Territory and northern Queensland (map 15).
MATERIAL EXAMINED: AUSTRALIA: Queensland: Lake Boronto near Somerset: Female neotype above ; Dividing Range, 15 km
W of Captain Billy Creek (11°40̍S, 142°45̍E), July 4–9, 1975 (G. B. Monteith), 2 juveniles ( QMB S49743), assigned tentatively ; Andoom near Weipa (12°31̍S, 141°50̍E), Feb. 5–8, 1975 (G. B. Monteith), rainforest, 13 ( QMB S49738) ; Gordon Creek, Iron Range (12°43̍S, 143°19̍E), June 24–30, 1976 (R. Raven, V. E. Davies), me sophyll vine forest, 2♀ ( QMB S886 View Materials ) ; same data, ‘‘+ Lamond Hill’ ’, 3♀ several juveniles ( QMB S49741 View Materials ) ; West Claudie Range, Iron Range (12°45̍S, 143°14̍E), Dec. 3–10, 1985 (G. Monteith, D. Cook), rainforest, 50 m elev., 23 3 juveniles ( QMB S50306 View Materials ) ; same collection data, 2♀ 1 juvenile ( QMB S50289 View Materials ) ; McIlwraith Range (~13°43̍S, 143°15̍E), July 20, 1995 (J. Thompson, M. Moulds, T. Olive), 13 ( AMS KS46098 ) ; Silver Plains Station , near Chester R. (13°58̍S, 143°22̍E), Nov. 2, 1985 (I. Fanning) 1♀ ( QMB S50307 View Materials ), assigned tentatively ; Thursday Island, Torres Strait (10°35̍S, 142°13̍E), Aug. 1949 (N.L.H. Krauss), 1♀ in AMNH ; Hammond Island, Torres Strait (10°32̍S, 142°13̍E), July 4–8, 1974 (H. Heatwole, E. Cameron), 1♀ ( QMB S50226 View Materials ), assigned tentatively ; Murray Island, Torres Strait (9°56̍S, 144°04̍E), July 1974 (H. Heatwole, E. Cameron), 23 2♀ ( QMB S50228) ; Yam Island, Torres Strait (9°53̍S, 142°55̍E), Nov. 28– Dec. 2, 1986 ( J. Gallon), under bark, night coll., 1♀ ( QMB S12400 View Materials ), assigned tentatively. Northern Territory: Gorge NE of Mt Gilruth (13°02̍S, 133°05̍E), July 10–13, 1979 ( G. Monteith, D. Cook), 1♀ several juveniles ( QMB S49771), assigned tentatively ; Kakadu National Park, Baroalba (12°50̍S, 132°57̍E), July 6, 1994 (M. S. Harvey, D. M. Hyder), 1♀ ( WAM 99 About WAM /1509), assigned tentatively ; Radon Creek (12°45̍S, 132°53̍E), rainforest, July 14–16, 1979 (G. Monteith, D. Cook), 23 4♀ some juveniles ( QMB S49752 View Materials ) ; North Point, Kapalga (12°25̍S, 132°22̍E), July 19, 1979 ( G. Monteith, D. Cook), 13 ( QMB S49745) ; W Alligator , ‘‘mouth, WA2’’ (12°11̍S, 132°16̍E), Nov. 1979 (R. Raven), 13 1♀ ( QMB S49769) ; Kemp Airstrip (12°35̍S, 131°20̍E), rainforest, July 24–25, 1979 (G. Monteith, D. Cook), 23 1♀ 1 juvenile ( QMB S49768 View Materials ) ; same locality, Nov. 15–16, 1979 (R. Raven), 23 2♀ 2 juveniles ( QMB S49755 View Materials ) ; Litchfield National Park, Wangi Falls (13°09̍S, 130°38̍E), May 26, 1992 (M. S. Harvey, J. M. Waldock), 23 1♀ 5 juveniles ( WAM 99 About WAM / 1501–8) .


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.