
Micromerys yidin, HUBER, 2001
|
publication ID |
https://doi.org/10.1206/0003-0090(2001)260<0001:TPOAAP>2.0.CO;2 |
|
persistent identifier |
https://treatment.plazi.org/id/5F4E362A-FFFD-5CF3-7A30-FB55FBDBF832 |
|
treatment provided by |
Marcus |
|
scientific name |
Micromerys yidin |
| status |
sp. nov. |
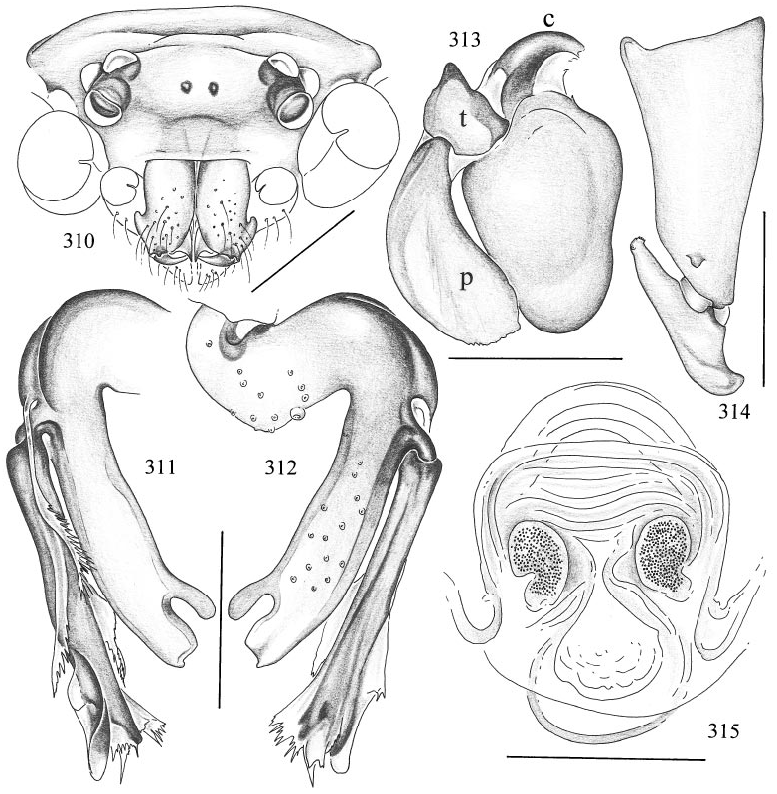
Micromerys yidin View in CoL , new species Figures 310–315 View Figs
Micromerys gracilis: DeelemanReinhold, 1986b: 206–209 View in CoL , figs. 1–4 (male misidentified).
TYPE: Male holotype from Spear Creek (16°42̍S, 145°24̍E), Queensland, Australia ; Nov. 3–10, 1975 ( R. Raven, V. E. Davies), in QMB ( S34685 View Materials ) .
ETYMOLOGY: Named for the Idindji (also called Yidin), aboriginal rainforest dwellers from the Cairns area, northwestern Queensland. The species name is a noun in apposition.
DIAGNOSIS: Distinguished from congeners by the shape of the lobes distally on the main branch of the procursus (figs. 311, 312), and by the very short, slightly curved bulbal apophysis (fig. 313).
MALE ( holotype): Total length 6.4, carapace width 0.71. Legs 1–3 missing, tibia 4: 5.9. Habitus and prosoma shape as in M. gracilis (cf. figs. 293, 294, 297, 298). Entire prosoma whitish ochre, only proximal part of clypeus light brown. Distance PMEPME 0.265; diameter PME 0.095; distance PME ALE 0.035; AME missing, but distinct black spots present (fig. 310). Chelicerae as in M. gracilis (cf. fig. 299). Palps in general as in M. gracilis (cf. figs. 295, 296), but femur ventrodistally with distinct apophysis (fig. 314); procursus and bulb as in figs. 311–313. Legs pale ochre, patellae (and femora tips) and tibia–metatarsus joints darker; apparently without spines, without curved and vertical hairs (many hairs missing); retrolateral trichobothrium of tibia 1 at 2.0% (male in SAM N1999/878); tarsi without pseudosegmentation. Opisthosoma shape as in M. gracilis (cf. fig. 293), monochromous pale ochre.
VARIATION: Tibia 1 in 8 males: 6.4–7.4 (x = 6.8).
FEMALE: In general very similar to male, but clypeus proximally not darker. Tibia 1 (N = 3) 5.9–6.1. Epigynum very simple, unpigmented roundish plate; dorsal view as in fig. 315.
DISTRIBUTION: Known only from the Cape Tribulation area and one suspicious locality in New South Wales (map 15; see Note below).
MATERIAL EXAMINED: AUSTRALIA: Queensland: Spear Creek : Male holotype above, with 13 ( QMB S49740 View Materials ) ; Cape Tribulation National Park , near beach (16°05̍S, 145°28̍E), July 24, 1992 (J. Coddington, G. Hormiga), 33 4♀ 2 juveniles (3 vials) ( USNM) ; Fritz Creek N of Bloomfield (15°52̍S, 145°21̍E), Dec. 1975 (M. Gray), rainforest foliage, 13 2♀ 5 juveniles ( AMS KS457 ) ; Twelve Mile Scrub (15°50̍S, 145°19̍E), Nov. 22–27, 1975 (collector not given), complex mesophyll vine forest, 13 1 juvenile ( QMB S49739 View Materials ) ; Shiptons Flat (15°48̍S, 145°15̍E), Nov. 16–21, 1975 (R. Monroe, V. E. Davies), vine forest on basalt, 23 1♀ ( QMB S887 View Materials , 891 View Materials ) ; Home Rule, Mt Hartley (15°44̍S, 145°18̍E), Nov. 11, 1974 ( V. E. Davies, D.J.), on leaves, 23 2♀ 1 juvenile ( QMB S49742 View Materials ). New South Wales: Bruxner Park , Coffs Harbour (30°10̍S, 153°05̍E), May 18, 1979 ( D. Hirst), 13 ( SAM N1999 View Materials /878), see Note below .
NOTE: The single record from New South Wales might be based on some labeling error and should be cited with some reservation until further material is collected from there. Revealingly, the single male shared the vial with a Micromerys male of another species ( M. gurran ) that is also otherwise only known from the Cape Tribulation area.
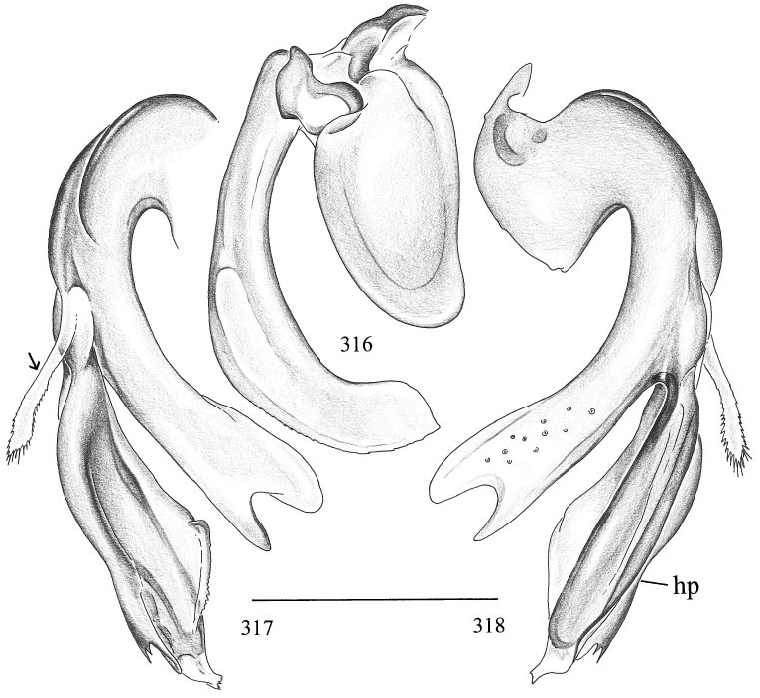
Micromerys gurran , new species Figures 316–318 View Figs
TYPE: Male holotype from Twelve Mile
Scrub (15°50̍S, 145°19̍E), Queensland, Australia ; Nov. 22–27, 1975 (collector not given), complex mesophyll vine forest on granite), in QMB ( S49763 View Materials ) .
ETYMOLOGY: In Yidini, the aboriginal language of the CairnsYarrabah region, gurran means ‘‘long’’. This refers to the long bulbal apophysis in this species. The species name is a noun in apposition.
DIAGNOSIS: Distinguished from congeners by the very long bulbal apophysis (fig. 316). M. wigi has a similar bulb, but the apophysis is closer to the globular part, and the distal lobes of the main branch of the procursus differ in shape (compare figs. 316–318 with 319–321).
MALE ( holotype): Total length 8.1, carapace width 1.23. Leg 1: 33.2 (9.1 + 0.5 + 8.0 + 12.7 + 2.9), tibia 2: 6.4, tibia 3: 4.1, tibia 4: 7.2. Habitus and prosoma shape as in M. gracilis (cf. figs. 293, 294, 297, 298). Carapace ochre to light brown, proximal part of clypeus light brown; sternum whitish. Distance PMEPME 0.305; diameter PME 0.120; distance PMEALE 0.045; AME with distinct black spots, but without lenses. Chelicerae as in M. gracilis (cf. fig. 299). Palps in general as in M. gracilis (cf. figs. 295, 296), but femur ventrodistally with distinct hump; procursus and bulb as in figs. 316– 318. Legs pale ochre, coxae, patella area and tibia–metatarsus joints darker; apparently without spines, without curved and vertical hairs (many hairs missing); retrolateral trichobothrium of tibia 1 at 2.0%; tarsi without pseudosegmentation. Opisthosoma shape as in M. gracilis (cf. fig. 293), monochromous ochregray.
VARIATION: Tibia 1 in 4 other males: 6.8, 7.2, 7.3, 7.5.
FEMALE: Unknown.
DISTRIBUTION: Known only from the Cape Tribulation area and one suspicious locality in New South Wales (map 15; see Note under M. yidin description above).
MATERIAL EXAMINED: AUSTRALIA: Queensland: Twelve Mile Scrub: Male holotype above ; Black Mtn (15°41̍S, 145°13̍E), Dec. 17, 1971 ( N. Clyde Coleman), 23 ( QMB S49746) ; Koah Road (16°49̍S, 145°31̍E), Apr. 2, 1972 ( N. Clyde Coleman), 13 ( QMB S49750 View Materials ). New South Wales: Bruxner Park, Coffs Harbour (30°10̍S, 153°05̍E), May 18, 1979 ( D. Hirst), 13 ( SAM N1999 View Materials / 877), see Note under M. yidin description above .
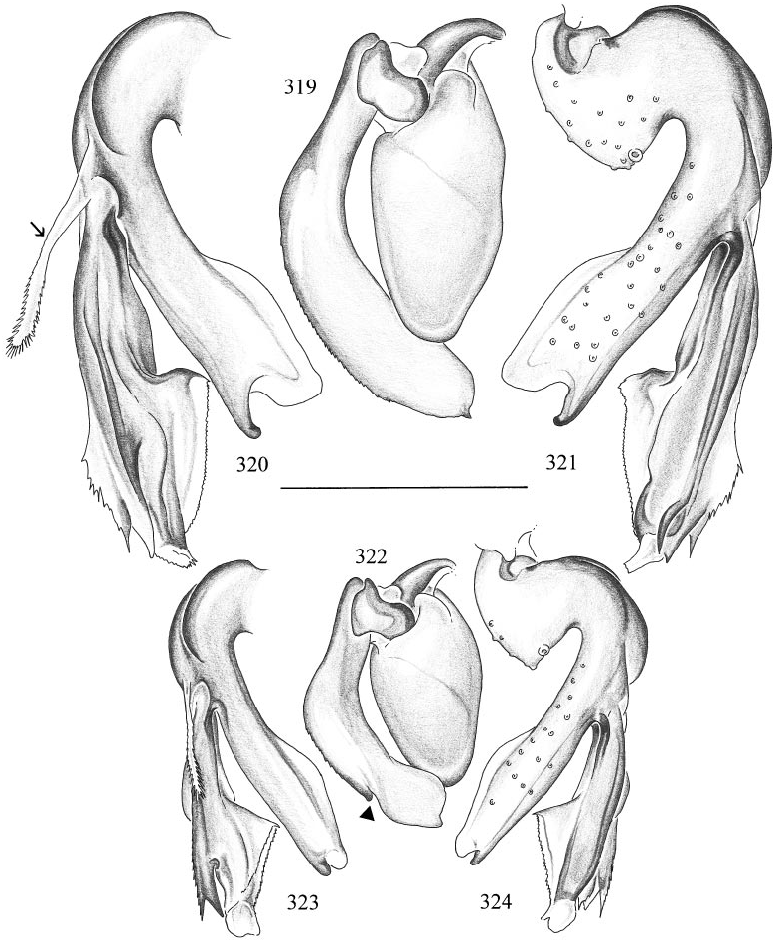
Micromerys wigi , new species Figures 319–321 View Figs
TYPE: Male holotype from Majors Mountain near Millaa Millaa (17°38̍S, 145°32̍E), Queensland, Australia ; Apr. 14–20, 1978 ( R. Raven, V. E. Davies), in QMB (S49753) .
ETYMOLOGY: In Yidini, the aboriginal language of the CairnsYarrabah region, wigi means ‘‘big’’. This refers to the larger size of this species compared to the closest known relative. The species name is a noun in apposition.
DIAGNOSIS: Distinguished from congeners by the shape of the bulbal apophysis (fig. 319) and by the shape of the distal lobes of the main branch of the procursus (figs. 320– 321). M. gidil is very similar in shape but has a notch on the bulbal apophysis, relatively and absolutely much smaller palps, and the PME are closer together.
MALE ( holotype): Total length 5.9, carapace width ~1.0 (deformed). Leg 1: 28.0 (7.3 + 0.4 + 6.7 + 11.1 + 2.5), tibia 2: 5.3, tibia 3: 3.5, tibia 4: 6.2. Habitus and prosoma shape similar to M. gracilis (cf. figs. 293, 294, 297, 298), but carapace more roundish in dorsal view, as in M. gidil . Carapace ochre, clypeus light brown except distal rim, sternum whitish. Distance PMEPME 0.305; diameter PME 0.095; distance PMEALE 0.040; with distinct black spots at AME position, but without lenses. Chelicerae as in M. gracilis (cf. fig. 299). Palps in general as in M. gracilis (cf. figs. 295, 296), but femur ventrodistally with distinct bulge; procursus and bulb as in figs. 319–321. Legs pale ochre, patella area and tibia–metatarsus joints darker; without spines, without curved and vertical hairs; retrolateral trichobothrium of tibia 1 at 4%; tarsi without pseudosegmentation. Opisthosoma shape as in M. gracilis (cf. fig. 293), pale ochre.
FEMALE: Unknown.
DISTRIBUTION: Known only from type locality, S of Cairns, Queensland (map 16).
MATERIAL EXAMINED: AUSTRALIA: Queensland: Majors Mountain: Male holotype above, with 2 juveniles in same vial.
Micromerys gidil , new species Figures 322–324 View Figs
TYPE: Male holotype from Gordonvale (17°06̍S, 145°47̍E), Queensland, Australia ; Feb. 1972 ( N. Clyde Coleman), in QMB ( S49772 View Materials ) .
ETYMOLOGY: In Yidini, the aboriginal language of the CairnsYarrabah region, gidil means ‘‘small’’. This refers to the small size of this species compared to the closest known relative. The species name is a noun in apposition.
DIAGNOSIS: Distinguished from congeners by the shape of the bulbal apophysis with its notch (arrow in fig. 322) and by the shape of the distal lobes of the main branch of the procursus (figs. 323–324). M. wigi is very similar in shape but has relatively and absolutely much larger palps (note that figs. 319–324 are all drawn to the same scale), and the PME are farther apart.
MALE ( holotype): Total length 6.1, carapace width 0.87. Leg 1: 24.7 (6.5 + 0.4 + 5.6 + 9.7 + 2.5), tibia 2: 4.4, tibia 3: 2.9, tibia 4: 4.9. Habitus and prosoma shape similar to M. gracilis (cf. figs. 293, 294, 297, 298), but carapace more roundish in dorsal view. Carapace ochre, proximal part of clypeus light brown; sternum whitish. Distance PMEPME 0.185; diameter PME 0.105; distance PMEALE 0.035; with distinct black spots in AME position, but without lenses. Chelicerae as in M. gracilis (cf. fig. 299). Palps in general as in M. gracilis (cf. figs. 295, 296), but femur ventrodistally with distinct bulge; procursus and bulb as in figs. 322–324. Legs pale ochre, patella area and tibia–metatarsus joints darker; without spines, without curved and vertical hairs; retrolateral trichobothrium of tibia 1 at 1.9%; tarsi without pseudosegmentation. Opisthosoma shape as in M. gracilis (cf. fig. 293), pale ochre.
VARIATION: Tibia 1 in 4 other males: 6.8, 7.2, 7.3, 7.5.
FEMALE: Very similar to male. Tibia 1 in 2 females: 5.9, 6.0. Epigynum extremely simple, unpigmented, roundish protruding plate (similar to M. raveni , cf. fig. 337); dorsal view similar to M. daviesae (cf. fig. 328).
DISTRIBUTION: Known only from the Cairns area, Queensland (map 16).
MATERIAL EXAMINED: AUSTRALIA: Queensland: Gordonvale: Male holotype above ; Bellenden Ker Range, Cableway Base Stn (17°16̍S, 145°54̍E), 100 m elev., Oct. 17–24, 1981 (Earthwatch/ QMB), 13 1♀ 2 juveniles ( QMB S27754 View Materials ) .
Micromerys daviesae DeelemanReinhold, 1986 Figures 325–333 View Figs View Figs
Micromerys daviesae DeelemanReinhold, 1986b: 209–210 View in CoL , figs. 10–17, 24, 59d, 60.
TYPE: Male holotype and female paratype from Finch Hatton (21°09̍S, 148°38̍E), Queensland, Australia ; Apr. 10, 1975 (R. Kohout, V. E. Davies), tangled web under leaf, in QMB (S888), examined.
DIAGNOSIS: Easily distinguished from congeners by the singlepointed end of the main branch of the procursus (figs. 326, 327).
MALE (Rundle Range): Total length 6.0, carapace width 0.97. Leg 1: 26.5 (6.8 + 0.4 + 6.0 + 10.5 + 2.8), tibia 2: 4.9, tibia 3: 3.5, tibia 4: 5.6; tibia 1 l/d: 75. Habitus and prosoma shape similar to M. gracilis (cf. figs. 293, 294, 297, 298), but carapace more roundish in dorsal view. Entire prosoma pale ochre, proximal part of clypeus darker; sternum whitish. Distance PMEPME 0.280; diameter PME 0.105; distance PMEALE 0.035; no trace of AME. Chelicerae as in M. gracilis (cf. fig. 299). Palps in general as in M. gracilis (cf. figs. 295, 296), but femur with ventrodistal bulge and procursus and bulb distinctive (figs. 325–327); tip of trochanter apophysis as in fig. 333; tip of hinged process as in fig. 331; palpal tarsal organ as in fig. 330. Legs pale ochre, patella area and tibia–metatarsus joints darker; apparently without spines, without curved and vertical hairs; retrolateral trichobothrium of tibia 1 at 3.5%; tarsi without pseudosegmentation (fig. 332). Opisthosoma shape as in M. gracilis (cf. fig. 293); monochromous pale ochre; gonopore with four epiandrous spigots (fig. 329).
VARIATION: Tibia 1 in 3 other males: 5.7, 6.7 ( holotype), 6.9. (Note that the original measurements are mostly wrong, based on miscalibration and mismeasurement; e.g., tibia 1 is 8.1 instead of 6.7, tibia 1 is shorter than tibia 2; etc.)
FEMALE: In general very similar to male, but clypeus proximally not darker. Tibia 1 in female paratype: 6.0. Epigynum very simple, unsclerotized, similar to M. raveni in ventral view (cf. fig. 337); dorsal view as in fig. 328.
DISTRIBUTION: Widely distributed in middleeastern Queensland (map 16).
MATERIAL EXAMINED: AUSTRALIA: Queensland: Finch Hatton: Male holotype and female paratype above ; Brandy Creek , E of Proserpine (20°21̍S, 148°43̍E), Apr. 24, 1975 ( R. Monroe, V. E. Davies), 13 ( QMB S889 View Materials ) ; Rundle Range (23°29̍S, 150°59̍E), on trees, Mar. 24–31, 1975 ( R. Kohout, V. E. Davies), 33 1♀ ( QMB S49766 View Materials ) ; same data, 13 1♀ ( QMB S890 View Materials ) .
Micromerys raveni , new species Figures 334–338 View Figs
TYPE: Male holotype from Stott’s Island (28°14̍S, 153°31̍E), New South Wales, Australia ; Nov. 18, 1978 ( R. Raven), in QMB ( S34684 View Materials ) .
ETYMOLOGY: Named for the collector of the type material and of many more pholcids in the Queensland Museum in Brisbane.
DIAGNOSIS: Easily distinguished from congeners by the two pointed lobes distally on the main branch of the procursus (figs. 335, 336).
MALE ( holotype): Total length 5.8, carapace width ~0.84 (deformed). Leg 1: 30.5 (7.9 + 0.4 + 7.2 + 11.9 + 3.1), tibia 2: 5.7, tibia 3: 3.9, tibia 4: 6.5; tibia 1 l/d: 83. Habitus and prosoma shape as in M. gracilis (cf. figs. 293, 294, 297, 298). Entire prosoma pale ochre, proximal part of clypeus light brown; sternum whitish. Distance PMEPME 0.280; diameter PME 0.100; distance PME ALE 0.025; single black spot in area of AME. Chelicerae as in M. gracilis (cf. fig. 299). Palps in general as in M. gracilis (cf. figs. 295, 296), but femur ventrodistally with rounded bulge; procursus, bulb, and trochanter apophysis as in figs. 334–336. Legs pale ochre, patella area and tibia–metatarsus joints darker; apparently without spines, curved, and vertical hairs (many hairs missing); retrolateral trichobothrium of tibia 1 at 2.0%; tarsi without pseudosegmentation. Opisthosoma shape as in M. gracilis (cf. fig. 293), monochromous pale ochre.
VARIATION: Tibia 1 in 2 other males: 6.8, 6.9.
FEMALE: In general very similar to male, but clypeus proximally not darker. Tibia 1 in 2 females: 5.9, 6.0. Epigynum very simple, unpigmented roundish protruding plate (fig. 337); dorsal view as in fig. 338.
DISTRIBUTION: Known from southeastern Queensland and northeastern New South Wales (map 16).
MATERIAL EXAMINED: AUSTRALIA: New South Wales: Stott’s Island: Male holotype above, with 13 2♀ 2 juveniles ( QMB S49765). Queensland: Searys Scrub, Cooloola (26°12̍S, 153°03̍E), Feb. 3, 1976 (R. Raven, V. E. Davies), 13 1♀ 1 juvenile ( QMB S49767 View Materials ).
REMARKS: With currently 110 nominal species, Pholcus is by far the largest genus
in pholcids. At first, it might seem that this simply results from Pholcus being the type genus, to which species were assigned before the establishment of other genera, and to which species tend to be assigned by workers unfamiliar with the other genera. Surprisingly, however, this seems not to be the case. Most of the ‘‘old’’ species not closely related to the type species have been removed (58 species previously assigned to Pholcus have been synonymized or transferred to other genera), and among the remaining 110 species there are very few that are obviously (e.g., P. everesti Hu and Li ) or probably (e.g., P. ciliatus Lawrence ) misplaced. At the same time, some species currently placed in other genera might actually be Pholcus (e.g., Spermophora elongata Yin and Wang ; S. longiventris Simon ; S. faveauxi Lawrence ).
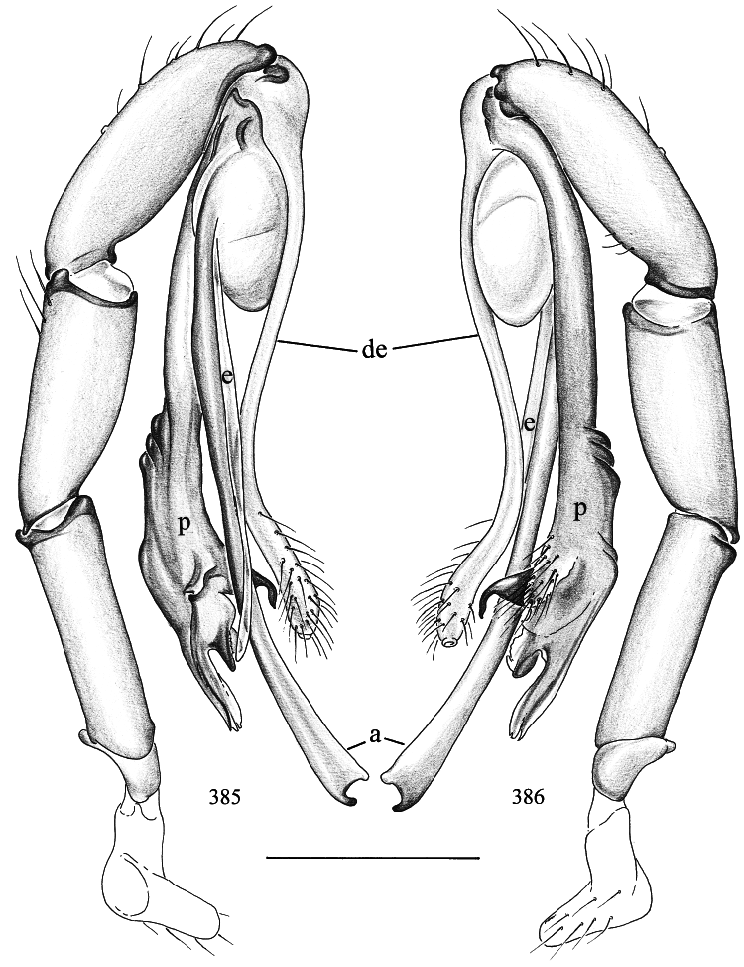
The characters most useful in distinguishing Pholcus from other genera seem to be the projections from the bulb, traditionally called uncus, appendix, and embolus (‘‘u’’, ‘‘a’’, and ‘‘e’’ in figs. 358, 372). The uncus is usually large, rather flat, heavily sclerotized, and is provided with many teeth or scales. The appendix is smaller, usually hookshaped, also sclerotized, and is either a single rod or split into two or even three parts. The em bolus lies between the uncus and appendix, is soft and transparent, and is thus easy to overlook. Other characters, not exclusive for Pholcus , are the conservative male chelicerae (a pair of dark frontal apophyses and a pair of light lateral apophyses; e.g., figs. 339, 369), the shape of the procursus (usually with ventral boss; e.g., figs. 343, 366), and the knob or wormshaped apophysis on the often roughly triangular or oval epigynum.
Most Pholcus species further share a cylindrical opisthosoma and eight eyes with lateral triads and small AME. The latter character is especially interesting, as eye position has traditionally been a key character for dis tinguishing pholcid genera. However, a recent cladistic analysis (Huber, 2000) has suggested that AME have been lost many times independently, and thus it comes as no surprise that this has also happened in Pholcus . The new Australian species described below as P. tagoman has the unmistakable genitalia of a real Pholcus , but it lacks any trace of the AME. I strongly suspect that Spermophora elongata was assigned to Spermophora simply because it lacks AME, but the genitalia and the long opisthosoma suggest it is also a sixeyed Pholcus .
The real taxonomic problem in Pholcus is its relationship to other genera of the Pholcus group, especially to Leptopholcus (Brignoli, 1980) . This genus shares with Pholcus all details of the chelicerae (including the modified hairs imbedded in the frontal apophyses), has a bulbal projection that seems to be a ho mologue of the uncus, and has a soft transparent embolus, but apparently no appendix. Some Korean and northern Chinese Pholcus species seem indistinguishable in their genitalia from Leptopholcus (e.g., most or all of those described in Paik [1978] as well as P. gaoi Song and Ren, and P. kwanaksaensis Namkung and Kim ). In fact, Leptopholcus might just be a polyphyletic group of pale, leafdwelling Pholcus species with a tendency to reduce the AME and the appendix.
More relevant for the present work is the presence of indigenous species of Pholcus in Australia. The only species previously recorded (except P. phalangioides ) was P. litoralis Koch, 1867 . It is thus not surprising that most Pholcus material in the collections had been identified as either P. phalangioides or P. litoralis . However, there is strong evidence that P. litoralis is a synonym of P. phalangioides ; that is, the illustrations in Koch (1872) clearly resemble P. phalangioides rather than any of the species newly described herein, and at least two of the three putative syntypes in the ZMH are P. phalangioides ( one male from Sydney and one female without locality information; the third specimen is a female from Queensland, but it lacks the opisthosoma; all were examined). All three vials carry labels presumably handwritten by Koch himself, and the last two could actually be types (the type localities are Brisbane and Rockhampton). Therefore, P. litoralis Koch, 1867 is here synonymized with P. phalangioides (Fuesslin, 1775) (NEW SYNONYMY).
A surprising fact is that P. ancoralis Koch, 1865 , which is very widespread in the Pacific region, is absent from the Australian collections studied. I have seen the type material of this species ( one male, two female prosomata from Upolu, Samoa Islands, in the ZMH) and numerous further specimens from the Samoa Islands (Sava’i), Fiji Islands, Moorea Islands (near Tahiti), New Hebrides (Espíritu Santo), and Loyalty Islands (Uréa) (all in AMNH). In contrast to the species newly described below, P. ancoralis is a representative of a mainly southeast Asian species group characterized by a pair of horns between the eye triads (including in addition at least P. bicornutus Simon from the Philippines and P. dentifrons Thorell from Burma).
Therefore, Pholcus is currently represent ed in Australia by P. phalangioides and the four species newly described below. Three of the endemic species occur in northern Queensland and are closely related to each other. The QMB has another closely related species, represented by a single poorly preserved specimen from Horn Island, Torres Strait. The sixeyed P. tagoman is restricted to the tropical north of Northern Territory, to Kimberley in Western Australia, and possibly reaches into northwestern Queensland. Its closest known relative is an undescribed species (or maybe two species) from Malaysia (in WAM 99/1994–5) and (interestingly) the Seychelles (M. Saaristo, unpubl. MS). The male pholcid from Sumatra identified by Kritscher (1957) (possibly correctly) as Spermophora longiventris Simon also belongs in this group. In fact, the Malaysian specimens mentioned above might be ‘‘ Spermophora ’’ longiventris .
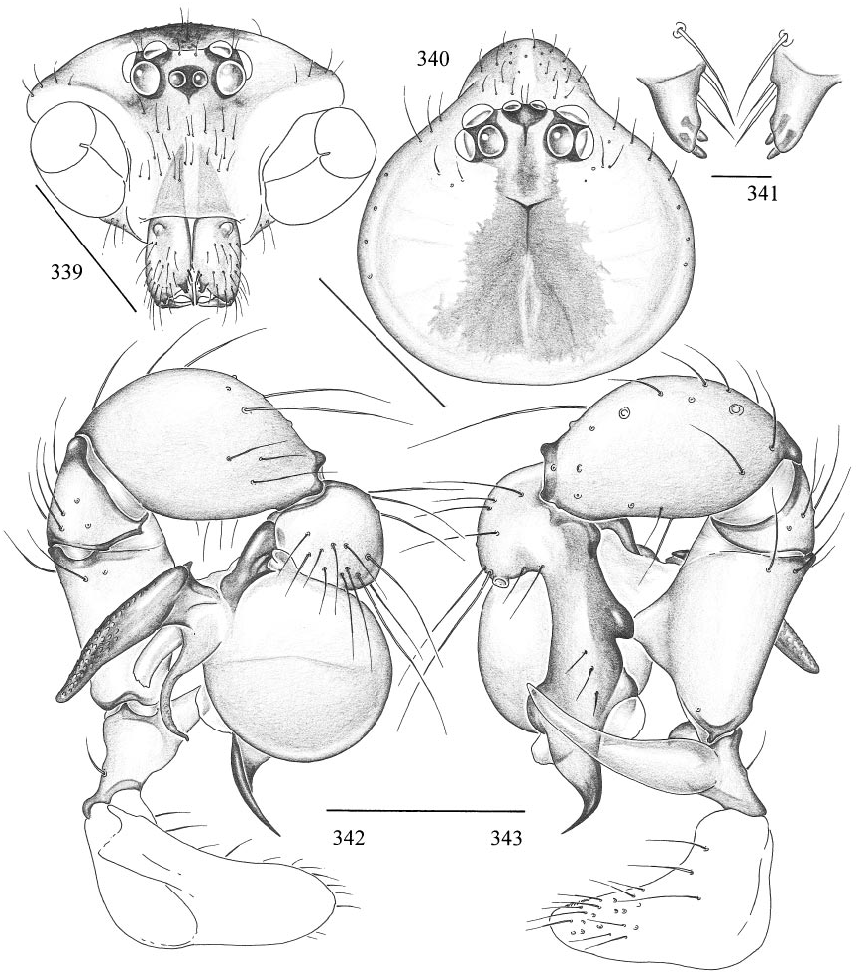
Pholcus jinwum , new species Figures 339–353 View Figs View Figs View Figs
TYPE: Male holotype from of E Claudie Scrub, Iron Range (12°43̍S, 143°18̍E), Queensland, Australia ; June 26, 1976 (collector unknown), in QMB ( S34689 View Materials ) .
ETYMOLOGY: Named for the Jinwum, an aboriginal tribe in the Iron Range area, northern Queensland. The species name is a noun in apposition.
DIAGNOSIS: Similar to P. dungara , distinguished by the shape of the procursus (compare figs. 343 and 355) and by the dark pattern on the epigynum (compare figs. 347 and 356).
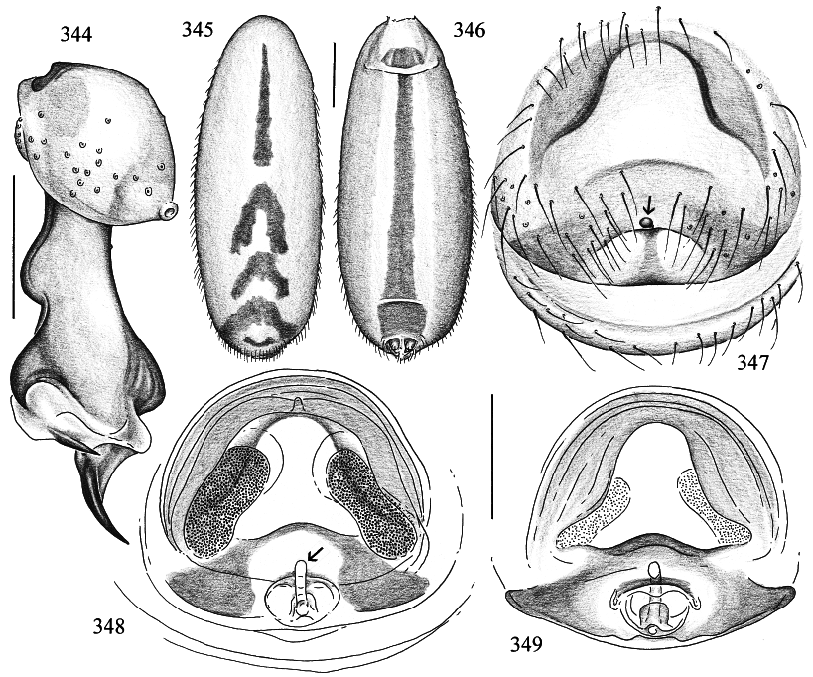
MALE ( holotype): Total length 6.9, carapace width 1.9. Leg 1: 48.4 (12.0 + 0.8 + 11.9 + 20.9 + 2.8), tibia 2: 8.3, tibia 3: 6.1, tibia 4: 7.7; tibia 1 l/d: 66. Prosoma shape as in figs. 339 and 340. Carapace ochre, with brown mark broadly connecting to ocular area (fig. 340); ocular area brown, clypeus ochre, without marks; sternum ochre to light brown. Distance PMEPME 0.295; diameter PME 0.160; distance PMEALE 0.025; diameter AME 0.120. Chelicerae as in fig. 339, with pair of black apophyses distally that are provided with two modified hairs each (fig. 341), and pair of unsclerotized rounded apophyses proximolaterally. Palps as in figs. 342 and 343; procursus as in fig. 344. Tarsal organ capsulate. Legs ochre, distal ends of femora and tibiae whitish, without dark rings; without spines, without curved and vertical hairs; retrolateral trichobothrium of tibia 1 at 4%; tarsal pseudosegments hardly visible in dissecting microscope, but regular in SEM (fig. 351). Opisthosoma shape as in figs. 345 and 346; pale ochre, dorsally with brown pattern as in fig. 345, ventrally with long brown line (fig. 346); gonopore in scanned male with five epiandrous spigots (fig. 350); several piriform gland spigots on ALS (fig. 353).
VARIATION: Tibia 1 in 8 males: 8.0–14.5 (x = 11.4). In some specimens the proximal cheliceral apophyses are slightly closer together.
FEMALE: In general very similar to male. Tibia 1 in 5 females: 8.7–10.8 (x = 9.6). Tip of palpal tarsus as in fig. 352. Epigynum as in fig. 347, ochre with distinctive light brown pattern; with wormshaped ‘‘knob’’ (arrows in figs. 347, 348). Dorsal view as in fig. 348, ventral cleared view as in fig. 349.
DISTRIBUTION: Known from several localities in northern Queensland (map 17).
MATERIAL EXAMINED: AUSTRALIA: Queensland: E of Claudie Scrub, Iron Range: Male holotype above, with 53 3♀ 1 juvenile ( QMB S49779 View Materials ) ; Portland Roads (12°35̍S, 143°23̍E), May 31–June 4, 1948 ( Archbold Exped.), 1♀ assigned tentatively, in AMNH ; Wenlock River Road Xing Moreton (12°03̍S, 141°56̍E), Sept. 11, 1985 ( M. Bennie), 2♀ assigned tentatively, ( QMB S49777 View Materials ) ; Prince of Wales Island (10°41̍S, 142°09̍E), July 3, 1976 ( E. Cameron), 13 ( QMB S50230 View Materials ) ; same locality, Feb. 11, 1975 ( E. Cameron), 1♀ ( QMB S50219 View Materials ) ; Horn Island, Torres Strait (10°37̍S, 142°17’), Dec. 2–8, 1986 ( J. Gallon), open forest, 13 1♀ ( QMB S12358 View Materials ) ; same locality, July 23, 1975 ( H. Heatwole, E. Cameron), 2♀ ( QMB S50220) ; Normandy Stn (15°23̍S, 144°52̍E), Sept. 13–17, 1984 ( C. Fearnley), sandstone cliffs, 13 ( QMB S49776 View Materials ) .
Pholcus dungara , new species Figures 354–357 View Figs
TYPE: Male holotype from Barron Gorge (16°50̍S, 145°38̍E), Queensland, Australia ; Jan. 1981 ( R. R. Jackson), in QMB ( S34686 View Materials ) .
ETYMOLOGY: Named for the Irukandji (also called Dungara), an aboriginal tribe from the Cairns area, northeastern Queensland. The species name is a noun in apposition.
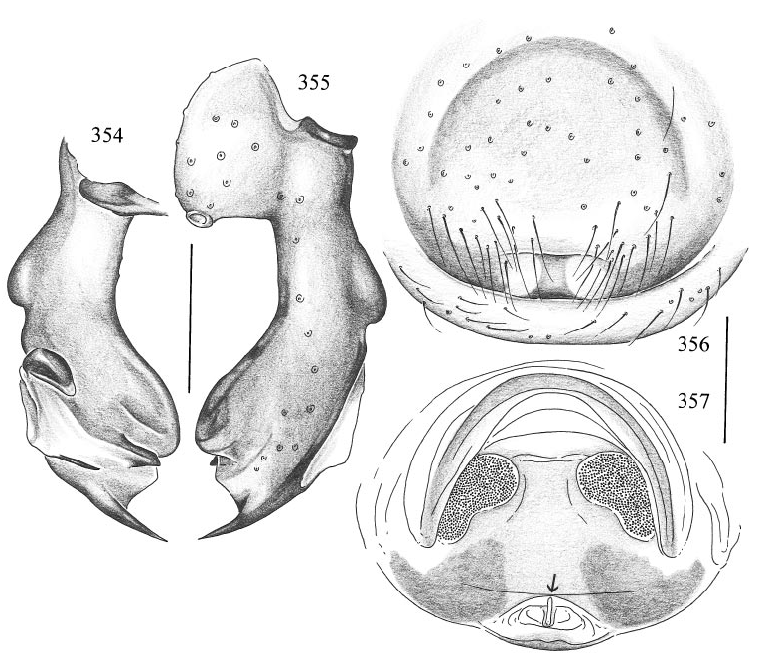
DIAGNOSIS: Very similar to P. jinwum , distinguished only by the shape of the procursus (compare figs. 343 and 355) and by the dark pattern on the epigynum (compare figs. 347 and 356).
MALE ( holotype): Total length 6.8, carapace width 1.8. Leg 1: 14.4 + 0.9 + 14.7, metatarsus and tarsus missing, tibia 2: 9.7, tibia 3: 6.7, tibia 4 missing; tibia 1 l/d: 79. Habitus and prosoma shape as in P. jinwum (cf. figs. 339, 340). Brown mark on carapace similar to P. jinwum (cf. fig. 340), but wider; ocular area brown, clypeus ochre, without marks; sternum pale ochre with small brown mark posteriorly. Distance PMEPME 0.265; diameter PME 0.120; distance PMEALE 0.040; diameter AME 0.080. Chelicerae as in P. jinwum (cf. fig. 339), but with additional indistinct hump between proximal apophyses. Palps in general as in P. jinwum (cf. figs. 342, 343), only procursus distinctively different (figs. 354, 355). Legs light brown, distal ends of femora and tibiae lighter; without dark rings; without spines, without curved and vertical hairs; retrolateral trichobothrium of tibia 1 at 4%; tarsus 2 distally with ~9 fairly distinct pseudosegments, proximally pseudosegments not visible in dissecting microscope. Opisthosoma shape as in P. jinwum (cf. fig. 345), with very similar brown pattern; four epiandrous spigots clearly visible in dissecting microscope.
FEMALE: In general very similar to male. Tibia 1 in 2 females: 12.7, 14.3. Epigynum as in fig. 356, ochre with distinctive light brown arch in front; wormshaped ‘‘knob’’ not visible in ventral view. Ventral cleared view as in fig. 357 (arrow points to small knob).
DISTRIBUTION: Known only from type locality near Cairns, Queensland (map 17).
MATERIAL EXAMINED: AUSTRALIA: Queensland: Barron Gorge: Male holotype above, with 13 3♀ ( QMB S49782 View Materials ).
Pholcus koah , new species Figures 358–362 View Figs
TYPE: Male holotype from Koah Road (16°49̍S, 145°31̍E), Queensland, Australia ; Apr. 2, 1972 ( N. Clyde Coleman), in QMB ( S34690 View Materials ) .
ETYMOLOGY: Named for the type locality. The species name is a noun in apposition.
DIAGNOSIS: Distinguished from the similar P. dungara and jinwum by the shape of the procursus (simple tip with two distinctive pointed terminal apophyses; figs. 359, 360) and from P. jinwum also by the dark pattern on the epigynum (compare figs. 347 and 361).
MALE ( holotype): Total length 6.6, carapace width ~1.9 (deformed). Leg 1: 46.2 (11.3 + 0.9 + 11.4 + 19.7 + 2.9), tibia 2: 8.1, tibia 3 missing, tibia 4: 7.7; tibia 1 l/d: 59. Habitus and prosoma shape as in P. jinwum (cf. figs. 339, 340). Brown mark on carapace similar to P. jinwum (cf. fig. 340), but without connection to ocular area; ocular area brown, clypeus ochre, without marks; sternum light brown with ochreyellow margins. Distance PMEPME 0.280; diameter PME 0.160; distance PMEALE 0.040; diameter AME 0.135. Chelicerae similar to P. jinwum (cf. fig. 339), but proximal apophyses closer together, more pointed and direct ed upward. Palps in general as in P. jinwum (cf. figs. 342, 343); bulb almost identical (fig. 358); ventral femur apophysis slightly more proximal; procursus distinctively different (figs. 359, 360). Legs light brown, distal ends of femora and tibiae lighter; without dark rings; without spines, without curved and vertical hairs; retrolateral trichobothrium of tibia 1 at 3.5%; tarsus 1 with many pseudosegments, but only ~13 distal ones distinct in dissecting microscope. Opisthosoma shape as in P. jinwum (cf. figs. 345, 346), with very similar brown pattern.
FEMALE: In general very similar to male. Tibia 1 in 4 females: 9.3, 9.5, 10.0, 10.0. Epigynum as in fig. 361; ochre with light brown arch in front, only tip of knob visible in ventral view. Dorsal view as in fig. 362 (arrows point to wormshaped ‘‘knob’’).
DISTRIBUTION: Known from three localities (15°41̍S, 145°20̍E), May 14, 1973 ( V. E. Davies), 1♀ assigned tentatively ( QMB S49775) ; Mt. Garnet (17°41̍S, 145°07̍E), Feb. 24, 1972 ( N. Clyde Coleman), 13 ( QMB S49773) ; Heron Island (23°27̍S, 151°55̍E), July 31, 1976 ( T. Newcomb), 2♀ assigned tentatively ( QMB S49780 View Materials ) .
in the Cairns area, northeastern Queensland, and possibly from Heron Island, southeastern Queensland (map 17).
MATERIAL EXAMINED: AUSTRALIA: Queensland: Koah Road: Male holotype above, with 2♀ ( QMB S49774 View Materials ) ; Amos Bay
Pholcus tagoman , new species
Figures 363–377 View Figs View Figs View Figs
TYPE: Male holotype from Ningbing Cave, cave KNI19 (15°18̍S, 128°37̍E), Western Australia; May 10, 1994 (R. D. Brooks), in WAM (99/2062).
ETYMOLOGY: Named for the Tagoman, an aboriginal tribe in the Daly River area in Northern Territory. The species name is a noun in apposition.
DIAGNOSIS: Easily distinguished from Aus tralian congeners by the absence of AME; from other described species also by the shapes of the bulbal apophyses and the procursus (figs. 365, 366, 371, 372). A very similar species occurs on the Seychelles (M. Saaristo, unpubl. MS). It differs by having a broader uncus and an appendix ending in two tips. Two specimens from Malaysia (WAM 99/1494 –5) and the male identified by Kritscher (1957) (possibly correctly) as Spermophora longiventris Simon might be conspecific with the Seychellian specimens. Finally, a single female specimen from Christmas Island, Phosphate Hill (10°26̍S, 105°42̍E) in the WAM (99/1512) might be conspecific with any of these species.
MALE ( holotype): Total length 3.8, carapace width 1.0. Leg 1: 37.3 (9.2 + 0.5 + 9.5 + 16.8 + 1.3), tibia 2: 5.6, tibia 3: 3.6, tibia 4: 5.3; tibia 1 l/d: 101. Habitus and prosoma shape as in figs. 363, 364, and 367. Carapace ochre, with graybrown median stripe; ocular area and clypeus without darker marks; sternum brown, shape as in fig. 368. Distance PMEPME 0.215; diameter PME 0.095; distance PMEALE 0.025; AME absent. Chelicerae as in fig. 369, with pair of black apophyses distally (figs. 370, 376) and pair of unsclerotized rounded apophyses proximolaterally. Palps as in figs. 365 and 366, procursus and bulb as in figs. 371 and 372. Palpal tarsal organ capsulate (fig. 377), on conical projection of cymbium (fig. 366). Legs ochrebrown, with barely visible darker rings on femora (subdistally) and tibiae (proximally, subdistally); without spines, without curved and vertical hairs; retrolateral trichobothrium of tibia 1 at 2%; tarsus 1 with>13 pseudosegments (proximally they are difficult to count, distally they are quite distinct; fig. 375). Opisthosoma shape as in fig. 363; gray, with blackish spots except ventrally; genital plate not distinct. ALS with several piriform gland spigots.
VARIATION: Tibia 1 in 8 males: 6.8–9.7 (x = 8.2). Opisthosoma in some males without dark spots.
FEMALE: In general very similar to male. Tibia 1 in 18 females: 5.5–8.5 (x = 7.5). Epigynum as in fig. 373; gray as opisthosoma with darker structures shining through, only knob brown to black; dorsal view as in fig. 374, apparently with divided pore plate. (Using the light microscope I could not see whether both plates are actually pore plates.)
DISTRIBUTION: Apparently widely distribut ed in northern Northern Territory, extending into northeastern Western Australia and possibly into northwestern Queensland (map 17).
MATERIAL EXAMINED: AUSTRALIA: Western Australia: Ningbing Cave , cave KNI19: Male holotype above, with 1♀ ( WAM 99 About WAM / 2063) ; same data but May 7–June 21, 1994 ( R. D. Brooks, B. Vine, W. Binks, K. Fear, C. Yeates, P. Fox, W. F. Humphreys), 13 14♀ several juveniles ( WAM 99 About WAM /2048–53, 2064, 2067, 2070–87, 3196) ; Ningbing Cave , cave KNI9 (15°17̍S, 128°37̍E), May 10, 1994 ( R. D. Brooks), 1♀ ( WAM 99 About WAM / 2065) ; Ningbing Cave , cave KNI29 (15°16̍S, 128°39̍E), May 9, 1994 ( R. D. Brooks), 13 2♀ 5 juveniles ( WAM 99 About WAM /2054–61) ; Ningbing Cave , cave KNI27 (15°17̍S, 128°41̍E), May 16, 1994 ( R. D. Brooks), 1♀ ( WAM 99 About WAM /2066) ; Jeremiah Hills , cave KJ8 (15°26̍S, 128°44̍E), May 17, 1994 ( W. F. Humphreys), 13 1♀ ( WAM 99 About WAM /2068–9) ; same locality, May 4, 1994 ( B. Vine), 1♀ ( WAM 99 About WAM /2046) ; Point Spring Nature Reserve (15°25̍S, 128°53̍E), May 28–June 11, 1999 ( M. Gray, G. Milledge, H. Smith), rainforest patch, 13 1♀ ( AMS KS56184–5 ) ; same data, under log, 13 1♀ ( AMS KS56186 ). Northern Territory: Cahills Crossing (12°25̍S, 132°58̍E), May 29, 1992 ( M. S. Harvey, J. M. Waldock), 13 ( WAM 99 About WAM /1513) ; Cutta Cutta Cave , cave K1 (14°35̍S, 132°25̍E), June 25, 1994 ( B. Jones, W. Binks, R. D. Brooks), 23 2♀ 6 juveniles ( WAM 99 About WAM /2035–44) ; Katherine, Cutta Cutta Guy Cave , 1988 ( E. Holland), 13 ( AMS KS22416 ) ; Kemp Airstrip (12°35̍S, 131°20̍E), rainforest, Nov. 15–16, 1979 ( R. Raven), 13 ( QMB S49805 View Materials ) ; Radon Creek (12°45̍S, 132°53̍E), rainforest, July 14–16, 1979 ( G. Monteith, D. Cook), 1♀ 1 juvenile ( QMB S49806 View Materials ). Queensland: Camooweal area, Great Nowranie Cave C6 (~20°05̍S, 138°10̍E), 1996 (collector unknown), 1♀ ( WAM 99 About WAM /2106), assigned tentatively ; same locality, Oct. 30, 1993 ( S. Eberhard), 1♀ ( AMS KS40772 ), assigned tentatively .
PANJANGE DEELEMANREINHOLD AND
DEELEMAN, 1983
REMARKS: I have seen only two of the six described species of the genus ( P. mirabilis and P. sedgwicki ), and can therefore only add a few details to what has been said before by DeelemanReinhold and Deeleman ( 1983), DeelemanReinhold (1986b), and Deeleman Reinhold and Platnick ( 1986).
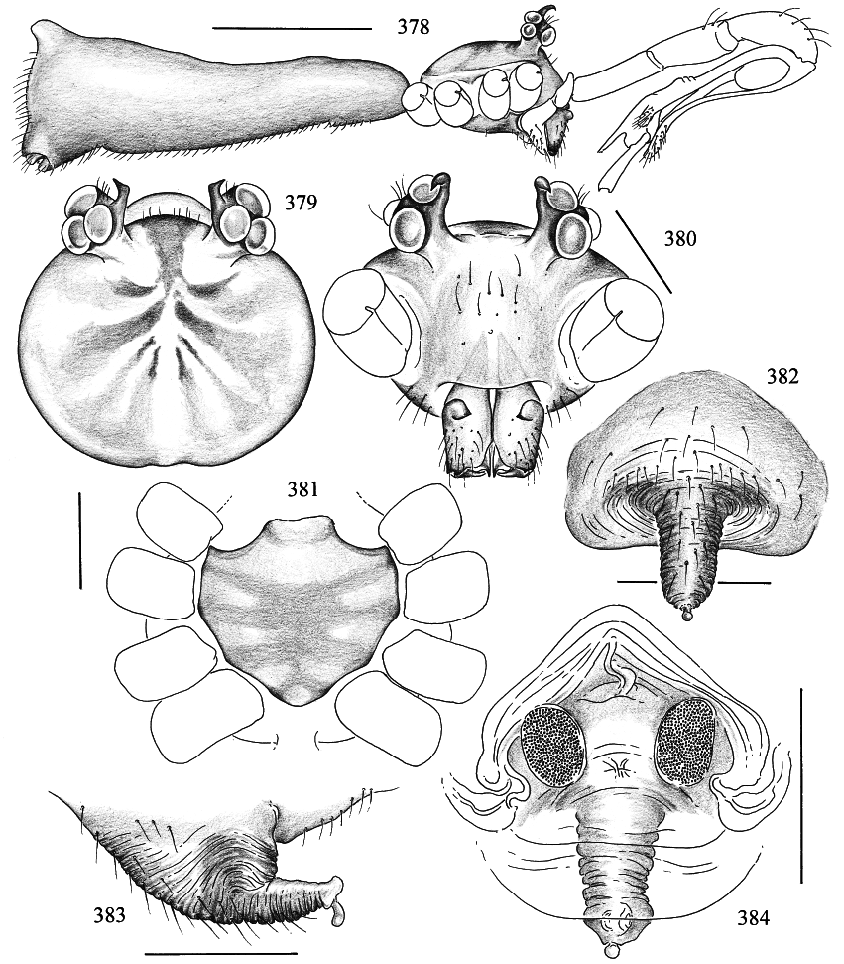
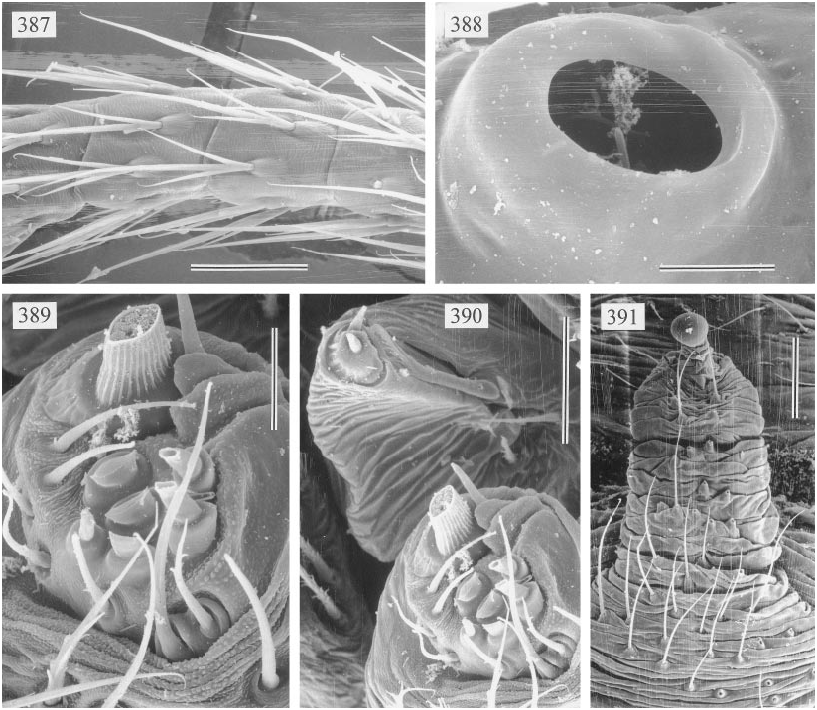
Several characters support the inclusion of Panjange in the Pholcus group of genera sensu Huber ( 1995). The ALS are provided with several piriform gland spigots (fig. 389), the tarsal organ is capsulate (fig. 388), and the male chelicerae are provided with proximal lateral apophyses (fig. 380). The knobshaped apophysis at the tip of the epigynum (figs. 382, 383, 391) is here considered a homologue of a similar structure in Pholcus and a few related genera, but this may be wrong. In Pholcus phalangioides and P. opilionoides , this structure is grasped by the male frontal cheliceral apophyses during copulation (Uhl et al., 1995; Huber 1995); some Panjange species have these frontal apophyses, but the species redescribed herein lacks the apophyses but nevertheless has the female knobshaped apophysis.
Panjange mirabilis DeelemanReinhold, 1986 Figures 378–391 View Figs View Figs View Figs
Panjange mirabilis DeelemanReinhold, 1986b: 220–222 View in CoL , figs. 45, 52–58, 59g.
TYPE: Male holotype ( QMB S883 View Materials ; right palp and chelicerae missing) and female paratype ( QMB S884 View Materials ) from Gordon Creek , Iron Range (12°43̍S, 143°19̍E), Queensland, Australia ; June 24–30, 1976 (R. Raven, V. E. Davies), mesophyll vine forest; examined.
DIAGNOSIS: Distinguished from known congeners by the shapes of the bulbal and cymbial apophyses (figs. 385, 386) (compare figures of congeners in DeelemanReinhold and Deeleman [1983] and DeelemanReinhold and Platnick [1986]).
MALE ( type locality, QMB S49788 View Materials ): Total length 3.2, carapace width 0.93; all legs loose or missing. Habitus and prosoma shape as in figs. 378–381. Entire prosoma whitish, only carapace with light brown pattern (fig. 379), and sternum with pale ochreyellow pattern. Eye triads elevated, with sclerotized spines (figs. 378–380); distance PMEPME 0.320; diameter PME 0.120; distance PME ALE 0.030; AME absent. Sternum as in fig.
381. Chelicerae whitish, with pair of pointed apophyses proximally, otherwise unmodified (fig. 380). Palps as in figs. 385 and 386, cymbium with complex procursus and simple dorsal elongation that bears the tarsal organ at its tip; bulb with simple tubular embolus and extremely long apophysis that ends in clawlike tip. Legs whitish, only patella area and tibia–metatarsus joints darker; most hairs missing. Opisthosoma shape as in fig. 378, whitish.
VARIATION: Carapace width in holotype: 0.84; tibia 2: 4.1, tibia 3: 2.5; the holotype has several dark spots on the opisthosoma.
FEMALE: In general very similar to male, but eyes not elevated and closer together. Females from West Claudie Range with very distinct brown pattern on carapace; sternum mostly brown; coxae brown; opisthosoma with large brown and blackish spots. Tibia 1 in 5 females: 5.7–6.3 (x = 6.0). Tarsal pseudosegments difficult to see in dissecting microscope, but distinct in SEM (fig. 387). Tarsal organ as in fig. 388. Epigynum as in figs.
382, 383, and 391; dorsal view as in fig. 384. Several piriform gland spigots on ALS (figs. 389, 390).
DISTRIBUTION: Adults have only been collected at Iron Range, northern Queensland. The QMB has two juveniles from Lockerbie and 3 km E Lockerbie (10°48̍S, 142°27– 28̍E) that might be conspecific (QMB S4786–7) (map 19).
MATERIAL EXAMINED: AUSTRALIA: Queensland: Iron Range, Gordon Creek: Male holotype and female paratype above ; same data, 13 2♀ several juveniles, ( QMB S49788 View Materials ) ; Iron Range, West Claudie Range (12°45̍S, 143°14̍E), Dec. 3–10, 1985 ( G. Monteith, D. Cook), rainforest, 50 m elev., 3♀ 1 juvenile ( QMB S34664 View Materials ) .
SPERMOPHORA HENTZ, 1841 AND BELISANA THORELL, 1898
REMARKS: Spermophora is probably the most chaotic genus within pholcids, and I have seen too few of the 69 species currently included in Spermophora and its possible closest relatives ( Spermophorides Wunderlich , Belisana Thorell , Paramicromerys Millot ) to attempt a significant improvement at this point. A few remarks might prove helpful for future workers.
Traditionally, almost any small pholcid with six eyes has been assigned to Spermophora . So far, the New World species have been either removed to Metagonia or Anopsicus , or synonymized with the synanthropic S. senoculata (Simon, 1893; Gertsch, 1939, 1971, 1977, 1982; Schmidt, 1971; Yaginuma, 1974; Brignoli, 1983; Huber, 2000). However, the remaining Old World species seem to be a highly heterogenous group of rather small, sixeyed pholcids. Simon (1893: 471) noted that the genus ‘‘ n’est pas très homogène ’’, and he distinguished two groups: one group (including the type species S. senoculata ) consisting of pale species with eye triads far apart, the other group characterized by black patterns on prosoma and opisthosoma and eye triads closer together.
Wunderlich (1992) formalized this distinction by creating the new genus Spermophorides for the second group, but unfortunately he treated representatives of the Canary Islands almost exclusively, ignoring even the Mediterranean species. These species ( S. huberti , mediterranea, mammata, petraea, simoni, valentiana, all described by Senglet) are obviously close to the Canary Island species, but they formally remain in Spermophora . More serious than that, the characters separating the two groups seem to work for Canary Island (and Mediterranean) species versus S. senoculata , but may not work in a worldwide perspective. For example, the new species S. yao described below has the eye triads far apart and a ventral flap on the pro cursus like S. senoculata , but it has blackish spots like Spermophorides . The only way to solve this satisfactorily will be a worldwide revision of the genus.
Unfortunately, revising just Spermophora and Spermophorides will not suffice, as the species described below as Belisana australis demonstrates. Belisana has been a monotypic genus since its creation by Thorell (1898), and it is with some hesitation that I assign the Australian species to this genus. I have not been able to see the type species B. tauricornis Thorell , which Simon (1903) first synonymized with Uthina Simon , but later (Simon, 1909) suspected to be just another Spermophora . Therefore, I follow mainly the opinion of DeelemanReinhold (1986a: 47), who reestablished Belisana based on a collection of 28 morphospecies ranging from Sri Lanka to Queensland, and who kindly sent me some of the specimens identified by her as Belisana . DeelemanReinhold (1986a) characterized the genitalic structure of Belisana as ‘‘somewhat similar to that in Spermophora ’’, and she (1986b: 223) used the eye distance to separate the two genera (she indicated the distance between triads as being 1–2 d in Spermophora , 3 d or more in Belisana ; this is a typing error, as the opposite is the case). Thus, Belisana seems to be something like a pale Spermophorides , or better, Spermophorides a dark Belisana ! Again, the conclusion is that only a deep revision will clarify the issue, and this will have to include Belisana as well.
Several EastAsian Spermophora might actually belong to Belisana (if Deeleman’s interpretation proves right). For example, illustrations of the genitalia of the Chinese S. anhuiensis Xu and Wang , of S. rollofoliolata Wang , and of the Japanese S. akebona Komatsu suggest a closer affinity to Belisana than to Spermophora .
Some Spermophora species are too obviously misplaced to present a serious problem. For example, judging from the existing illustrations, S. elongata Yin and Wang and S. faveauxi Lawrence are very probably Pholcus (the latter is close to, or is a synonym of Pholcus circularis Kraus ); S. baso Roewer from Sumatra and S. tessellata Simon from Malaysia are close relatives and certainly not Spermophora (types examined), but they probably represent a new genus; S. dubia Kulczynski from New Guinea is a relatively large spider with long opisthosoma and an epigynum that is almost indistinguishable from that of Panjange mirabilis (cf. figs. 382, 383), but the slightly damaged, opisthosoma of the female holotype (examined) was apparently not pointed dorsoposteriorly as in typical Panjange ; and S. longiventris from Sumatra was described from a juvenile, and is probably a sixeyed Pholcus , close to P. tagoman .
At least some Spermophora species from Madagascar seem to be misplaced and might end up in new genera (e.g., S. andrei , impudica), while some others ( S. ankaranae , combesi, imerinensis, megaceros) resemble Paramicromerys in genitalic structure, a monotypic genus endemic in Madagascar. Millot (1946: 146) interpreted Paramicromerys as ‘‘ faisant transition entre les Spermophora et les Micromerys véritables ’’, adding yet another genus to the task awaiting the future revisor of Spermophora .
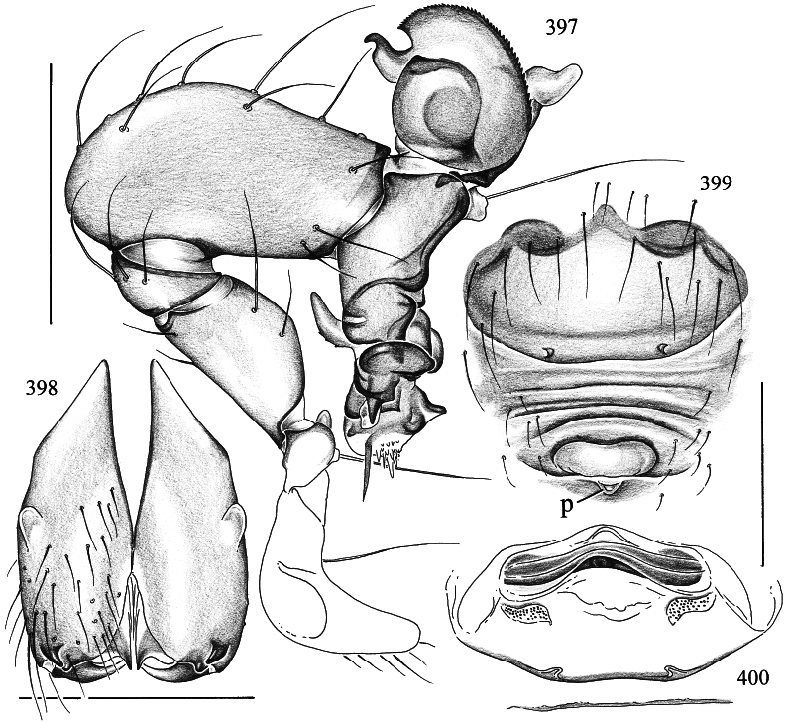
Spermophora yao , new species Figures 392–405 View Figs View Figs View Figs
TYPE: Male holotype from Gordon Creek , Lower Lamond Hill, Iron Range (12°43̍S, 143°19̍E), Queensland, Australia ; June 15, 1976 ( R. Raven, V. E. Davies), litter, in QMB ( S34666 View Materials ) .
ETYMOLOGY: Yao is an alternative name for the Pakadji, an aboriginal tribe in northeastern Queensland. The species name is a noun in apposition.
DIAGNOSIS: Distinguished from congeners by the shape of procursus and bulbal apophyses (figs. 396, 397), and by the shape of the epigynum (fig. 399).
MALE ( holotype): Total length 1.1, carapace width 0.55. Leg 1: 4.34 (1.13 + 0.19 + 1.15 + 1.26 + 0.61), tibia 2: 0.74, tibia 3: 0.61, tibia 4: 1.10; tibia 1 l/d: 18. Habitus and prosoma shape as in figs. 392–395. Carapace and sternum ochre to light brown with blackish speckles, ocular area and clypeus without speckles (figs. 393–395). Eye triads hardly elevated; distance PMEPME 0.215; diameter PME 0.050; distance PMEALE 0.010; AME absent. Chelicerae as in fig. 398, with pair of simple pointed apophyses dis tally and pair of rounded light apophyses proximolaterally. Palps as in figs. 396 and 397, with complex procursus; bulb with semitransparent projection (embolus?) and sclerotized serrate apophysis with distal hook. Legs ochreyellow, without rings; without spines, without curved and vertical hairs; retrolateral trichobothrium of tibia 1 at 15%; tarsus 1 with ~10 fairly distinct pseudosegments. Opisthosoma globular (fig. 392); pale ochre gray, with some indistinct large spots dorsally. ALS with several piriform gland spigots, as in female.
VARIATION: Tibia 1 in 6 males: 1.05–1.23 (x = 1.12).
FEMALE: In general very similar to male. Tibia 1 in 18 females: 0.87–1.13 (x = 0.98). Tarsal pseudosegments as in fig. 402. Palpal tarsal organ and tip as in figs. 403 and 405. Epigynum as in figs. 399 and 401; frontally brown plate with pair of pockets, posteriorly dark ridges, and short, transparent scape (arrow in fig. 401) with pocket (‘‘p’’ in fig. 399) between epigynum and spinnerets (scape hardly visible in dissecting microscope); dorsal view as in fig. 400. Several piriform gland spigots on ALS (fig. 404).
DISTRIBUTION: Known only from Iron Range, northern Queensland (map 18).
MATERIAL EXAMINED: AUSTRALIA: Queensland: Iron Range, Gordon Creek: Male holotype above, with 43 12♀ several juveniles ( QMB S50172 View Materials ) ; Iron Range, West Claudie Range (12°45̍S, 143°14̍E), sieved litter in rainforest, 50 m elev., Dec. 4, 1985 ( G. Monteith), 23 10♀ several juveniles ( QMB S33856 View Materials ) .
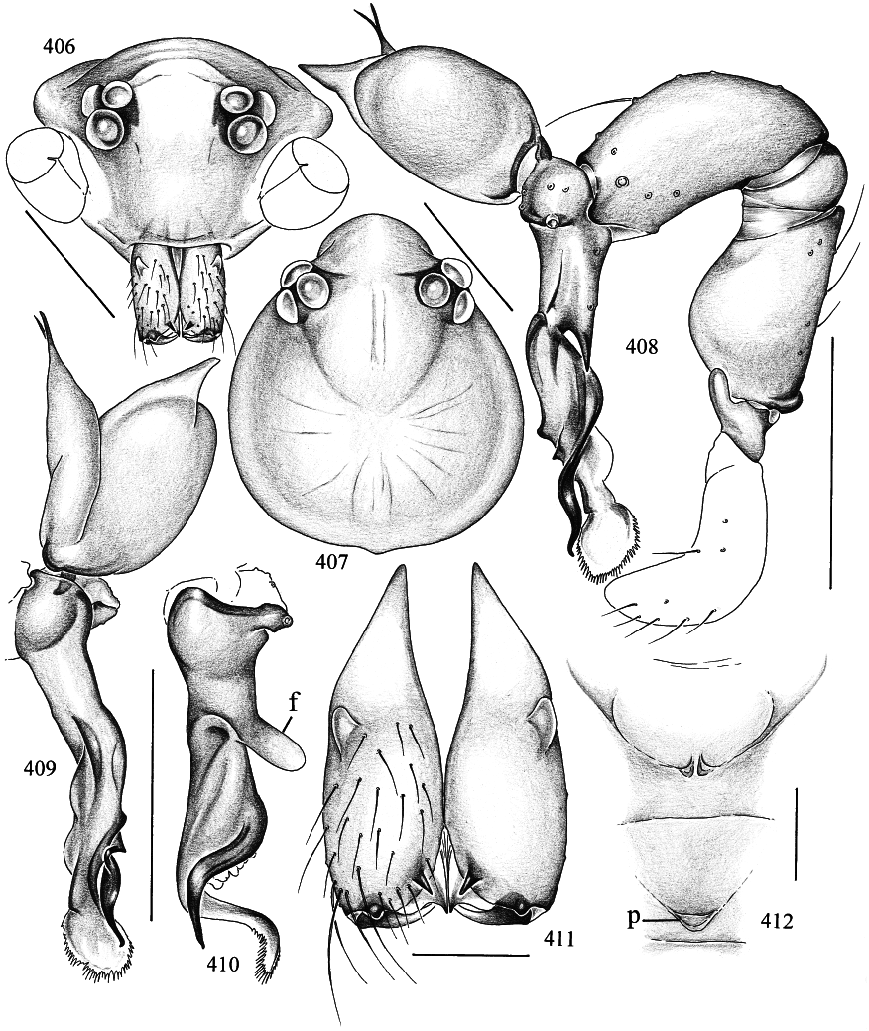
Spermophora paluma , new species Figures 406–415 View Figs View Figs
TYPE: Male holotype from Paluma SF (19°01̍S, 146°13̍E), Queensland, Australia ; Sept. 2, 1988 ( R. Raven, T.C., J. Gallon), in QMB ( S14092 View Materials ) .
ETYMOLOGY: Named for the type locality. The species name is a noun in apposition.
DIAGNOSIS: Distinguished from known congeners by the shape of procursus and bulbal apophyses (figs. 408–410), and by the shape of the epigynum (fig. 412). The AMS has a very close undescribed relative from Lord Howe Island (31°33̍S, 159°05̍E) (KS56205, 56217) that is significantly larger (total length 2–2.1; carapace width 0.8; tibia 1: 4.5–4.6) and has only one tip on the putative embolus (instead of two as in figs. 408, 409); chelicerae and procursus are almost identical to S. paluma .
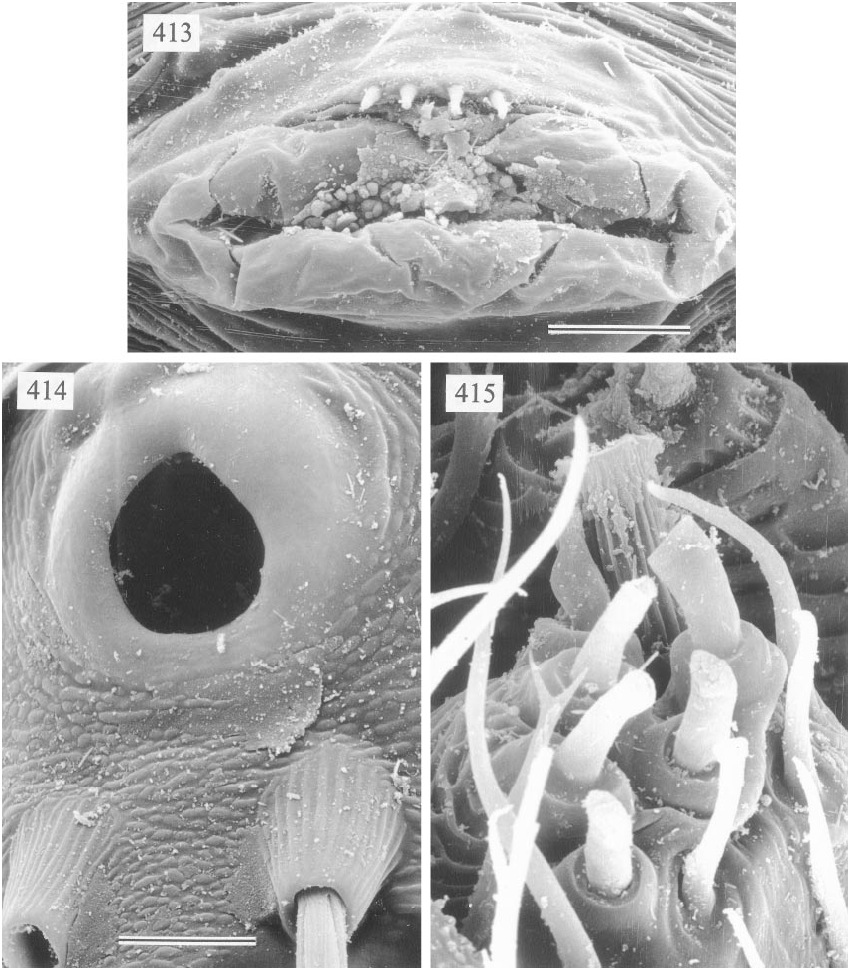
MALE ( holotype): Total length 1.4, cara pace width ~0.56 (deformed). Leg 1: 17.2 (4.3 + 0.3 + 4.1 + 7.3 + 1.2), tibia 2: 2.6, tibia 3: 1.6, tibia 4: 2.7; tibia 1 l/d: 68. Prosoma shape as in figs. 406 and 407; entire prosoma pale ochre; sternum whitish. Eye triads hardly elevated; distance PMEPME 0.185; diameter PME 0.070; distance PME ALE 0.010; AME absent. Chelicerae as in fig. 411, with pair of simple pointed apophyses distally and pair of rounded light apophyses proximolaterally. Palps as in fig. 408, with complex procursus; bulb with two simple projections, one of them (putative embolus) ending in two black tips (figs. 408 and 409); procursus as in figs. 408–410. Legs pale ochreyellow, patella area slightly dark er, ventroproximal margin dark brown; without spines, without curved and vertical hairs; retrolateral trichobothrium of tibia 1 at 3%; tarsus 1 distally with ~10 fairly distinct pseudosegments, proximally possibly more. Tarsal organ as in fig. 414. Opisthosoma pale gray, shape similar to S. yao (cf. fig. 392), but slightly longer (0.93 long, 0.67 high);
gonopore with four epiandrous spigots (fig. 413); several piriform gland spigots on ALS (fig. 415).
VARIATION: Tibia 1 in 10 males: 3.5–4.1 (x = 3.8). Some males with large blackish spots on opisthosoma.
FEMALE: In general very similar to male. Epigynum as in fig. 412; extremely pale, with pair of pockets in frontal plate and unpaired pocket on short, transparent posterior scape between epigynum and spinnerets (‘‘p’’ in fig. 412).
DISTRIBUTION: Known from several localities in northeastern Queensland (map 18).
MATERIAL EXAMINED: AUSTRALIA: Queensland: Paluma SF: Male holotype above, with 23 ( QMB S14092 View Materials ) ; Wallaman Falls via Ingham (18°36̍S, 145°48̍E), rainforest, 500 m elev., Oct. 1, 1980 ( G. Monteith), 13 3 juveniles ( QMB S50247 View Materials ) ; Mt. Graham (18°23̍S, 145°52̍E), Dec. 26–30, 1986 ( S. Hamlet), rainforest, 600–700 m elev., 13 3 juveniles ( QMB S18422 View Materials ) ; Kirrama Range (18°12̍S, 145°45̍E), Douglas Creek Road, 800 m elev., Dec. 9–12, 1986 (G. Monteith, G. Thompson, S. Hamlet), 13 ( QMB S49803) ; Kirrama Range via Kennedy (~18°09̍S, 145°43̍E), rainforest, 500 m elev., Oct. 2, 1980 ( G. Monteith), 13 1 juvenile ( QMB S49760) ; Boonjee (17°24̍S, 145°44̍E), Apr. 3–6, 1978 ( V. E. Davies, R. Raven), 23 1 juvenile ( QMB S50270 View Materials ) ; Windin Falls, via Butchers Creek (17°19̍S, 145°45̍E), rainforest, Oct. 9, 1980 (G. Mon teith), 33 3 juveniles ( QMB S50241 View Materials ) ; 3 km N of Kuranda, Kuranda State Forest (~16°47̍S, 145°38̍E), rainforest, 360 m elev., June 25–Oct. 3, 1982 ( S. & J. Peck), 13 ( AMNH) ; 20 km SW of Mossman, Mt. Lewis (~16°35̍S, 145°15̍E), rainforest, 900 m elev., June 26–Oct. 1, 1982 ( S. & J. Peck), 13 ( AMNH) ; Fritz Creek , N of Bloomfield (15°52̍S, 145°21̍E), rainforest foliage, Dec. 1975 ( M. Gray), 13 ( AMS KS65695 ) .
Belisana australis , new species
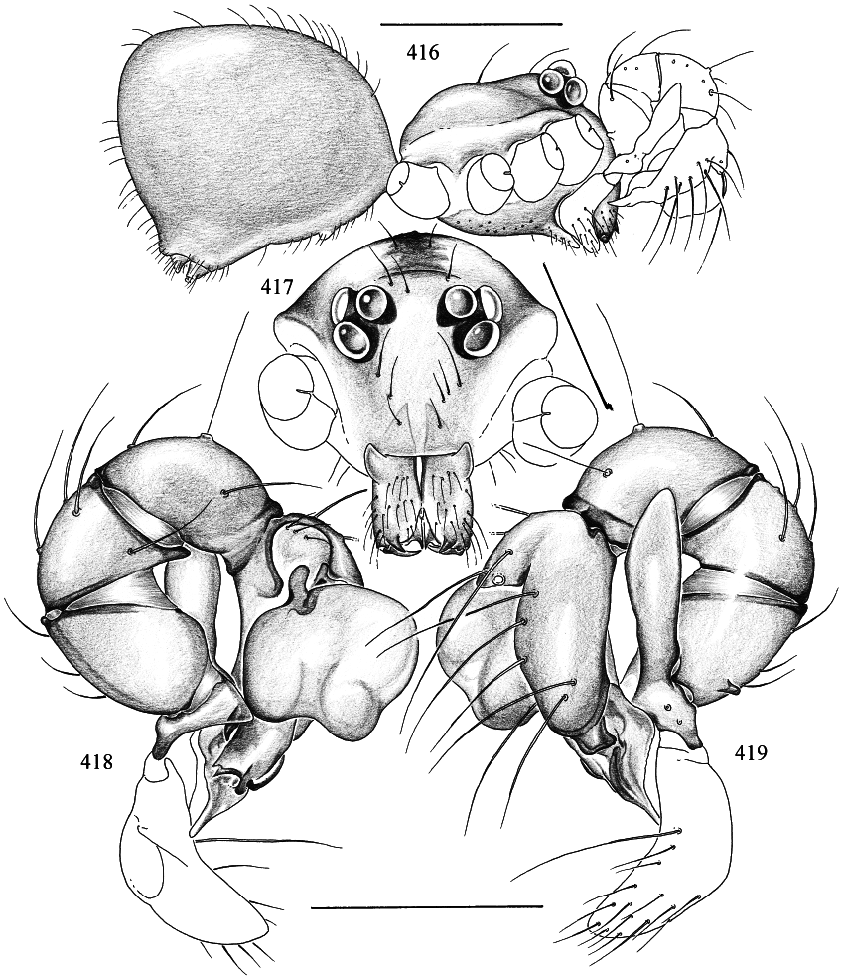
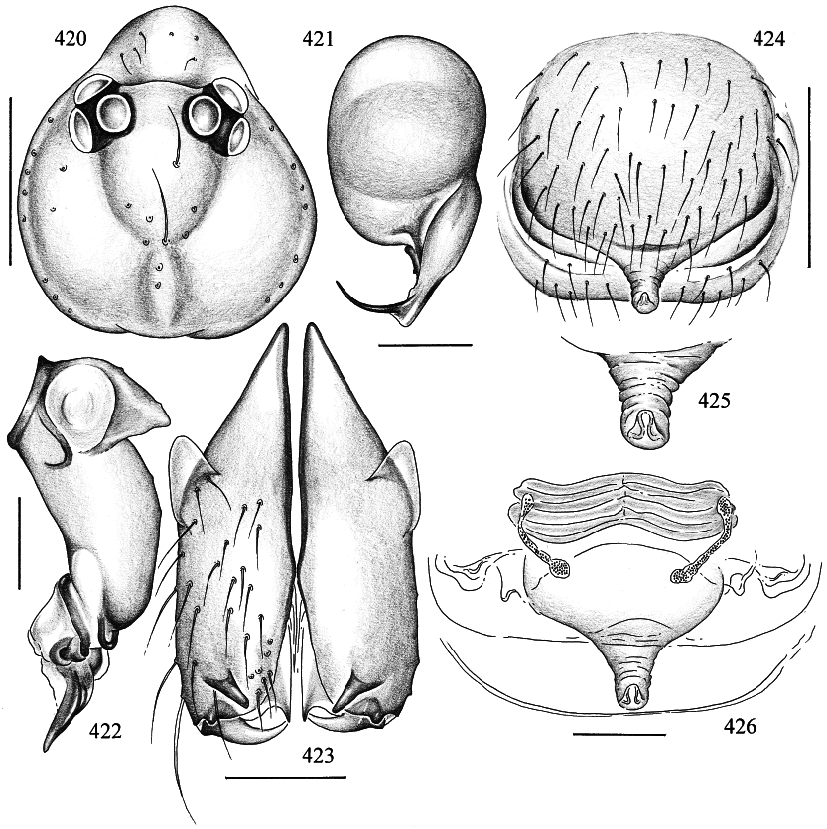
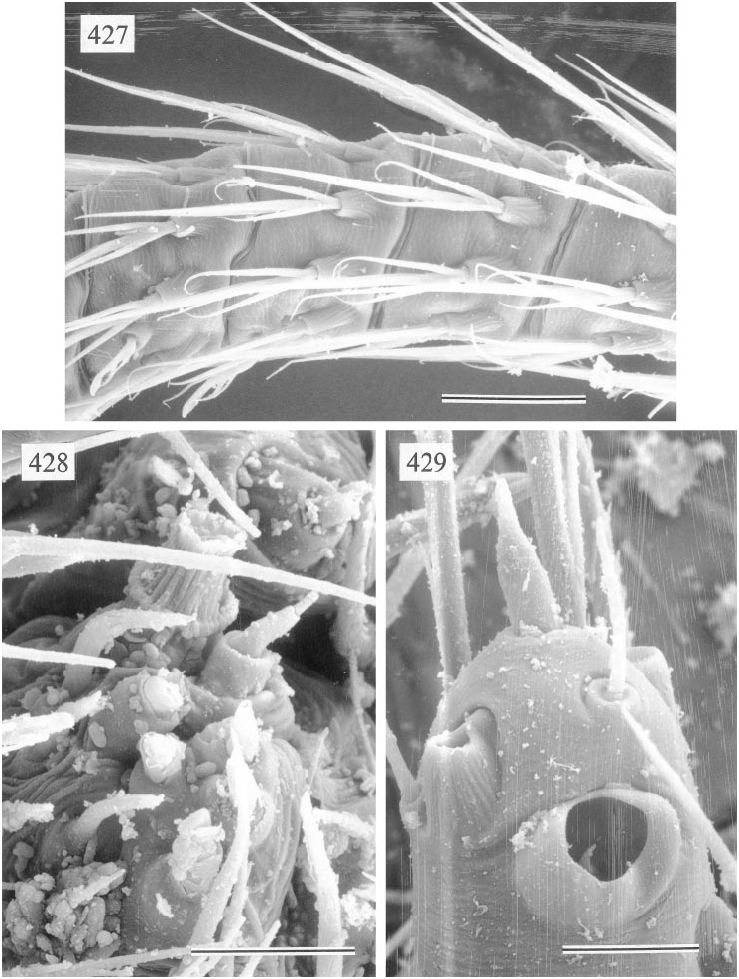
Figures 416–429 View Figs View Figs View Figs
TYPE: Male holotype from Home Rule (15°44̍S, 145°18̍E), Queensland, Australia ; Oct. 28–Nov. 18, 1974 ( V. E. Davies), litter, in QMB ( S50273 View Materials ) .
ETYMOLOGY: The species name (Latin australis , ‘‘southern’’) refers to the geographic distribution of this southernmost Belisana species.
DIAGNOSIS: Tiny sixeyed pholcid with globular opisthosoma; distinguished from all described pholcids where the male is known by the huge apophysis on the male palpal trochanter (fig. 419). The collection CLD has a male of a probably undescribed species from Thailand with a similar apophysis, but with very different chelicerae.
MALE ( holotype): Total length 1.2, carapace width 0.52. Leg 1: 8.65 (2.19 + 0.19 + 2.24 + 3.03 + 1.00), tibia 2: 1.42, tibia 3: 0.87, tibia 4: 1.34; tibia 1 l/d: 48. Habitus and prosoma shape as in figs. 416, 417, and 420. Carapace light ochre, with slightly dark er bands posteriorly and around ocular area (fig. 420); ocular area and clypeus without markings; sternum pale ochre. Ocular area not elevated; distance PMEPME 0.105; diameter PME 0.070; distance PMEALE 0.020; AME absent. Chelicerae as in fig. 423, with pair of simple pointed apophyses distally and pair of rounded light apophyses proximolaterally. Palps as in figs. 418 and 419, with huge trochanter apophysis and small apophysis on femur retrolaterodorsally; procursus and bulb relatively simple (figs. 421, 422). Legs light ochre, without rings; without spines, curved, and vertical hairs; retrolateral trichobothrium of tibia 1 at 18%; tarsus 1 distally with ~14 fairly distinct pseudosegments (fig. 427), proximally pseudosegmentation difficult to see in dissecting microscope. Opisthosoma almost globular (fig. 416), ochregray; ALS with several piriform gland spigots; gonopore with four epiandrous spigots in two groups.
VARIATION: Tibia 1 in 5 males: 1.7–2.5 (x = 2.13).
FEMALE: In general very similar to male. Palpal tarsus tip as in fig. 429. Epigynum as in fig. 424, with distinctive scape provided with distal pair of pockets (fig. 425); dorsal view as in fig. 426. Several piriform gland spigots on ALS (fig. 428).
DISTRIBUTION: Known from several localities in Australia (northern Northern Territory and northern Queensland; map 19) and from Ambon, Moluccas.
MATERIAL EXAMINED: AUSTRALIA: Queensland: Home Rule: Male holotype above, with 1♀ (epigynum lost) in same vial ; Clifton Beach (16°46̍S, 145°40̍E), 1971– 1972 (N. ClydeColeman), 1♀ ( QMB S50284 View Materials ) ; West Claudie Range, Iron Range (12°45̍S, 143°14̍E), Dec. 3–10, 1985 (G. Monteith, D. Cook), rainforest, 33 3 juveniles ( QMB S34660 View Materials ) ; Murray Island, Torres Strait (9°56̍S, 144°04̍E), Aug. 1974 (H. Heatwole, E. Cameron), 13 1 juvenile ( QMB S50231 View Materials ). Northern Territory: West Alligator River , ‘‘mouth, WA2’’ (12°11̍S, 132°16̍E), rainforest, Nov. 12, 1979 (R. Raven), 3♀ 2 juveniles ( QMB S50173 View Materials ) ; same locality, Jan. 1979 (R. Raven), 1♀ 2 juveniles ( QMB S50180 View Materials ) ; West Alligator River , ‘‘mouth, WA1’’ (12°12̍S, 132°13̍E), Nov. 12, 1979 (R. Raven), 13 1♀ 5 juveniles ( QMB S50175) ; East Alligator Xing (12°25̍S, 132°58̍E), sieved litter in rainforest, July 18, 1979 (G. B. Monteith), 23 6 juveniles ( QMB S50179) ; North Point, Kapalga (12°25̍S, 132°22̍E), rainforest, Nov. 11, 1979 (R. Raven), 13 1♀ ( QMB S50171 View Materials ) ; Radon Creek (12°45̍S, 132°53̍E), Nov. 14, 1979 ( R. Raven), litter, wet rainforest, 23 3 juveniles ( QMB S50177). INDONESIA: Moluccas: Ambon , Jan. 17, 1995 ( C. L. & P. R. Deeleman), 13 ( CLD) .










No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Micromerys yidin
| HUBER, BERNHARD A. 2001 |
Micromerys gracilis: DeelemanReinhold, 1986b: 206–209
| : Deeleman-Reinhold 1986: 206 - 209 |
Micromerys daviesae DeelemanReinhold, 1986b: 209–210
| Deeleman-Reinhold 1986: 209 - 210 |
Panjange mirabilis DeelemanReinhold, 1986b: 220–222
| Deeleman-Reinhold 1986: 220 - 222 |